
B. Singh
Department of Botany, Faculty of Sciences, Udai Pratap Autonomous College, Varanasi, India
D. Katiyar
Department of Biochemistry and Bioprocess Technology, Jacob School of Biotechnology and Bio-engineering, Sam Higginbottom Institute of Agriculture, Technology and Sciences, Allahabad, India
D.V. Singh
Department of Botany, Faculty of Sciences, Udai Pratap Autonomous College, Varanasi, India
A.K. Kashyap
Department of Botany, Faculty of Sciences, Banaras Hindu University, Varanasi, India
A.M. Lall
Department of Biochemistry and Bioprocess Technology, Jacob School of Biotechnology and Bio-engineering, Sam Higginbottom Institute of Agriculture, Technology and Sciences, Allahabad, India
International Journal of Agricultural Research
Year: 2011 | Volume: 6 | Issue: 9 | Page No.: 699-706







ABSTRACT
Rice fields are one of the major biogenic sources of atmospheric methane. Apart from this contribution to the greenhouse effect, rice paddy soil represents a suitable model system to study fundamental aspects of microbial ecology, such as diversity, structure and dynamics of microbial communities as well as structure function relationships between microbial groups. The present study was conducted in rainfed rice fields planted to rice (Oryza sativa) cultivar, NDR-97, to evaluate the variation of population of Methane Oxidizing Bacteria (MOB) in different soil type (Bare, Bulk and rhizosphere) over a period of 13 weeks. Urea was the only fertilizer applied, at a rate of 100 kg N ha-1 in three split doses. The experiment was laid out in a randomized complete block design with three replicate plots for treatments. The soil exhibited higher numbers of MOB in control plots of bulk and rhizospheric (37.4x106 and 58.87x106 cells g-1 dry soil) than in plots treated with urea (28.6x106 and 51.9x106 cells g-1 dry soil) at 80 Days after Sowing (DAS) and were highest in the rhizospheric soil (58.87x106 cells g-1 dry soil) followed by bulk (37.4x106 cells g-1 dry soil) and bare (2.2x106 cells g-1 dry soil) in unfertilized soil but bare (control) soil was attained highest MOB (2.7x106 cells g-1 dry soil) on 40 DAS and MOB significantly decreased in fertilized soil. The concentrations of NH4+-N were significantly (p<0.05) lower in the rhizosphere (1.3 μg g-1 soil) than in bulk (3.7 μg g-1 soil) and bare soils (4.1 μg g-1 soil) on 80 DAS in unfertilized plots. In fertilized soil NH4+-N concentration were increased due to lower number of population at different day’s intervals. The study suggests that the development of the rice rhizosphere brings about a spatial pattern in the distribution of methanotrophic bacteria which increases in size, over time; within the rhizosphere and adjoining bulk soil and that the rhizosphere is a potential microsite of methanotrophic bacterial activity.
PDF Abstract XML References Citation
Received: December 31, 2010;
Accepted: October 01, 2011;
Published: October 31, 2011
How to cite this article
B. Singh, D. Katiyar, D.V. Singh, A.K. Kashyap and A.M. Lall, 2011. Impact of Urea on Spatio-temporal Distribution of Methanotrophic Bacteria in Rainfed Rice Agro Ecosystem. International Journal of Agricultural Research, 6: 699-706.
DOI: 10.3923/ijar.2011.699.706
URL: https://scialert.net/abstract/?doi=ijar.2011.699.706
DOI: 10.3923/ijar.2011.699.706
URL: https://scialert.net/abstract/?doi=ijar.2011.699.706
INTRODUCTION
Rice (Oryza sativa) is one of the most important cereal crops, with 143 million ha under cultivation globally and grown in wide range of climatic zones, to nourish the mankind (Roger et al., 1993). It is principal food crop of some state of India. Narendra-97 (NDR-97) variety is popular commercial central zone of Uttar Pradesh, Orissa, Assam and West Bengal. NDR 97 rice is the source of cash income for many farmers of Chandauli district.
Rice fields are one of the major anthropogenic sources of methane (CH4); a greenhouse gas (Neue, 1997) to the atmosphere. The atmospheric concentration of CH4 is expected to increase further due to expansion of rice cultivation (Singh and Singh, 1995). The only known biological sink for atmospheric methane is its oxidation in aerobic soil by methanotrophic bacteria (Hutsch et al., 1996). This sink can contribute up to 15-45 Teragram CH4 year-1 to the total methane destruction. CH4 is produced in the saturated soils of rice fields by anaerobic bacteria, the methanogens and escapes to the atmosphere mainly through the system of airspaces in the plant body (Singh and Singh, 1995). Association of populations of Methane Oxidizing Bacteria (MOB) with rice rhizosphere contributes to CH4 oxidation. The rainfed rice soil which is largely aerobic and harbours a substantial size of MOB population (Dubey and Singh, 2000) has been shown to be a net sink for atmospheric methane (Singh et al., 1998, 1999a). The total rice cultivated area in India is approximately 42.3x106 ha, out of which 6.3x106 ha (15%) is under upland rice cultivation (Adhya et al., 2000). The dryland rice areas may assume importance is taken up by these soils. Methanotrophs (gram negative, aerobic bacteria belonging to the family Methylococcaceae) oxidize CH4 via methane monooxygenase enzyme (Holmes et al., 1995). The absence of this soil sink would cause the atmospheric concentration of methane to increase about 1.5 times the current rate (Duxbury, 1994).
On a global scale, methanotrophic bacteria oxidize more than half of the methane produced. Rice fields account for approx. 20% of global methane emissions, estimations ranging from 10 to 25%. Field measurements indicate that 10 to 50% of the methane produced in rice fields is not emitted due to its reoxidation in the rhizosphere and at the soil surface (Denier van der Gon and Neue, 1996). Three major habitats for microorganisms in paddy fields can be specified: (1) the anoxic bulk soil (2) the oxic surface soil and (3) the partially oxic rhizosphere with increased substrate concentration (Conrad, 2007). Methanotrophs can be found in habitats where methane and oxygen gradients overlap (Henckel et al., 2001; Eller and Frenzel, 2001). Particularly this gradient is present at the surface of the paddy soil and the rhizosphere (Bosse and Frenzel, 1997; Gilbert and Frenzel, 1995). However, a large amount of methanotrophs can be detected in the anoxic bulk soil (Eller et al., 2005; Eller and Frenzel, 2001). MOB population size differed among bare, bulk and rhizosphere soils of a dry land rice field and the MOB population growth was suppressed by the application of urea (Dubey and Singh, 2000). We have also seen that rhizosphere soil has a greater CH4 oxidizing capacity than the bulk and bare soils (Dubey and Singh, 2000). It has remained to be seen whether or not the CH4 oxidation capacity of the soil is influenced, in conformity with the MOB population size, by the N-fertilizers commonly used in dryland rice cultivation. Methanotrophs are strictly aerobic because their key enzyme, methane monooxygenase, requires molecular oxygen. They occur at oxic-anoxic interfaces where both methane and oxygen are available. In rice fields, the rhizosphere is such an environment because rice roots are supplied with atmospheric oxygen through the aerenchyma. Oxygen diffuses into the soil, creating an oxygenated zone around the roots (Frenzel et al., 1992). On the other hand, the aerenchyma serves as a conduit for methane from methanogenic bulk soil to the atmosphere. Both the rhizoplane and the rhizosphere are therefore suspected to house methane-oxidizing bacteria (MOB). The association of MOB with plants has been studied with both classical and molecular techniques. Our own MPN counts revealed a significant enrichment of MOB in the rice rhizosphere (Gilbert and Frenzel, 1995). Sediment free roots of many aquatic macrophytes oxidized methane (King, 1994), so did rice roots too (Frenzel and Bosse, 1996). Recently, reported on the oxidation of propylene to propylene oxide by excised roots and a basal portion of the stem indicating the presence and activity of methane monooxygenase (Watanabe et al., 1997).
Application of nitrogen fertilizers, among which NH4+-based fertilizers are most common, is necessary for rice production. Consequently, effects of NH4+-based fertilizers on CH4 emission greatly attract the attention of scientists. However, the results from numerous studies have so far been inconsistent, ranging from stimulation (Banik et al., 1996; Singh et al., 1999b) to inhibition (Bodelier et al., 2000a, b). The effects of NH4+-based fertilizers depend on type and amount of the fertilizer, as well as on mode and time of application (Neue and Sass, 1994). The present study system comprised bare, bulk and rhizosphere soils of control and fertilizer urea treated plots of a dryland rice field.
MATERIALS AND METHODS
Experimental site and rice cultivation: Present study was carried out on the rainfed rice field of the Chandauli district in July 2007, India. The region is characterized by seasonally dry tropical climate with typical monsoonal features and the year is divisible into a cold winter (November-February), a hot summer (April-June) and a warm rainy season (July-September). During the experiment, minimum temperatures ranged from 14 to 27°C and the maximum from 22 to 38°C. The soil is a well-drained Inceptisol, pale brown, silty loam (sand 32, silt 65 and clay 3%) with pH 7-7.8. The experimental field consisted of 12 plots each measuring 5x3 m. The experiment was laid down in a completely randomized block design. A 0.5 m strip separated plots. Basal treatment of KCl+P2O5+farm-yard manure was applied at a rate of 60:60:1000 kg ha-1, to all plots during plowing. Six plots were fertilized with urea and the remaining served as control. In the fertilized plots, urea was applied in three split doses, at the time of tillering, flowering and grain filling stage at the rates of 40, 30 and 30 kg N ha-1, respectively. Among the 12 plots, six plots (three with and three without urea) were sown to rice while the other six (three with and three without urea) were maintained as bare soil. Thus the experiment had three plots each for bare control, bare fertilized, vegetated control and vegetated fertilized treatments. Seeds of rice (Oryza sativa L., cultivar Narendra-97) were sown by dibbling on July 1997, at a spacing of 15 cm (hill-to-hill) by 20 cm (row-to-row) in the plots designated as vegetated plots. No irrigation was provided throughout the experiment and the sole source of water was rainfall.
Soil sampling and analysis for NH4+-N: Samples of bulk (between the plant rows), bare (bare plots) and rhizosphere soil were collected separately for each plot from 0-10 cm depth using a 5 cm diameter soil corer. The 0-10 cm soil depth was chosen because observation indicated that ≥92% roots are concentrated in this soil layer. The rhizospheric soil was collected by tapping the roots on a plastic sheet (Lee et al., 1997). The soil samples were sieved (2 mm) and fine roots were removed. Field moist samples stored at 4°C were used for chemical analyses and methanotrophic population counts within 2 days after sampling. The soil sampling was carried 20-day intervals after sowing (DAS). Ammonium nitrogen (NH4 +-N) was measured by the phenate method (Claude, 1979) in an extract with 2 M KCl.
Plant growth measurements: The growth of the rice plants was monitored every 20 days up to harvest. One rice hill was harvested from each experimental plot on each sampling date and roots were collected as soil as a block (15x20x15 cm depth) using a rectangular open-top plastic chamber. Roots were washed with water. The roots and shoots were dried separately at 60°C for 48 h for biomass determination. The soil was subjected to careful washing with tap water. Subsequently, the roots and shoots were separated from each other. All estimations described above were conducted in triplicate.
Population of methanotrophs: The numbers of methanotrophic bacteria were enumerated by the MPN (most probable number) technique as described by Bender and Conrad (1992). The pH was adjusted to 6.8. A trace element solution was added after autoclaving (Gilbert and Frenzel, 1995). Dilution was carried out from 10-1 to 10-9, as described by Espiritu et al. (1997). Each dilution, 1 mL was inoculated into tubes containing 3 mL NMS medium. There were six replicates for each dilution. After inoculation under aseptic conditions, the tops of the tubes were closed with sterilized cotton plugs. The tubes were incubated under 20% methane in air at 25°C in the dark in atmosbags (Sigma, USA) for 3 weeks. For control, culture tubes were prepared without soil inoculum (Espiritu et al., 1997). In tests we had used control with sterilized soil and found that control without soil was as good as a control with sterilized soil. After 3 weeks of incubation, positive wells had a cloudy appearance. Most probable numbers were obtained using Rowe's tables (Rowe et al., 1977). Further, a more reliable method to enumerate cultivable MOB would be MPN in tubes (6-8 weeks incubation).
Statistical analysis: Data were checked for normality and homogeneity of variances and subjected to Analysis of Variance (ANOVA) according to Snedecor and Cochran (1989). All data analyses and statistical comparisons were performed using an SPSS package (SPSS 13). A General Linear Model (GLM) two-way ANOVA with repeated measures was used to analyze the effect of soil type, fertilizer on soil methanotrophic bacterial population. To determine the significance of differences between means, a Tukey’s HSD test was used to determine the significance of differences between cropping season averages.
RESULTS
Ammonium-N (NH4+-N): Present results showed a greater accumulation of NH4+-N in bare soil which was followed by bulk and rhizosphere soil (Table 1) and the differences were significant (F 2,12 = 102.3; p<0.05)(Table 2). Urea treated soil had the greater concentration of NH4+-N followed by control soils (Table 1) at 80 days intervals. Differences due to treatment were significant (F1,12 = 397.97; p<0.05). HSD test detect significant differences in NH4+-N concentrations between control and fertilized soils of different soil type (bare, bulk and rhizospheric).
Crop growth pattern: In the present study, we measured plant height, root biomass and shoot biomass as affected by urea fertilization at different day’s interval (Table 3). There was a significant effect of urea treatment on these growth characteristics. Levels of urea fertilizer significantly affected plant height. Plant height ranged from 33.2 to 70.4 cm. ANOVA indicated significant differences due to treatment (F1,4 = 17.5; p<0.05), day interval (F3,12 = 81.5; p<0.05). The root biomass peaked earlier than shoot biomass and thereafter declined slowly. The highest shoot biomass was attained on 80 DAS in both control (260.0±13.8 g m-2) and fertilized (380.0±4.0 g m-2). Similar to root biomass also peaked at the flowering stage of the plant, but following the peak, root volume declined rather sharply. ANOVA for shoot biomass, root biomass showed significant differences due to treatment (F1, 4 = 110.2; p<0.05, F1,12 = 204.5; p<0.05) and day’s interval (F3,12 = 929.0; p<0.05, F3,12 = 541.38; p<0.05) and their interaction treatmentxvarieties (F3, 12 = 63.4; p<0.05, F3,12 = 208.2; p<0.05), respectively. The decline in growth variables after a certain stage was evident in response to senescence and weathering growth in all the varieties, both in control as well as in fertilized plots. Application of urea enhanced the growth of rice plant in this study.
| Table 1: | Size of MOB population (x 106 cells g-1 dry soil) and NH4+-N concentration (μg g-1 dry soil) in rhizosphere, bulk and bare soil from control and fertilized (0 and 100 kg N ha-1) planted to rainfed rice variety NDR-97 on six sampling dates (DAS = days after sowing) as affected by N fertilizers |
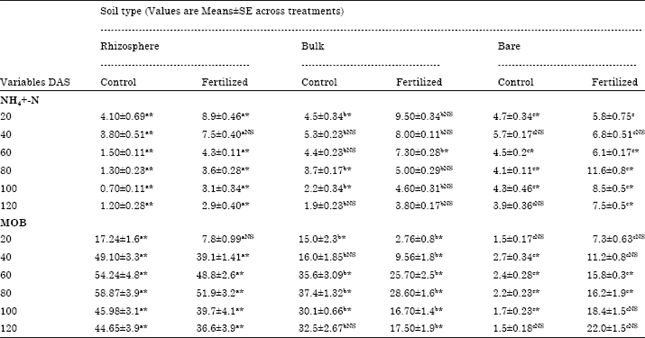 | |
| Data are expressed as Mean±SE of three replicates in each treatments of vegetative and unvegetative (bare) plots; aRhizosphere vs. bare, bBulk vs. Rhizosphere and cBare vs. Bulk showed comparison between different soil types. Values in a row bearing superscript * are significantly different and NS for not significant from each other at p<0.05 according to Tukey’s HSD test. | |
| Table 2: | F-ratio and their significance levels for two-way ANOVA with repeated measures for soil parameters NO3--N, NH4+-N and CH4 Oxidizers for three soil type (rhizospheric, bulk and bare of a variety NDR-97 and two fertilization treatments (0 and 100 kg N ha-1) |
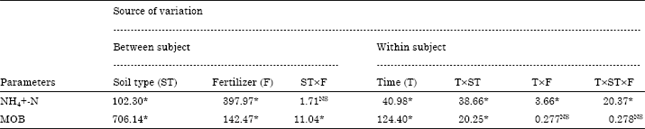 | |
| NS: Not significant, * Significant at p<0.05 | |
Population of methanotrophs: Most of the known methanotrophic bacteria can grow on nitrate-based mineral salt medium in this study. However, there may be some that do not and there are probably many others that have not been cultivated at all. Therefore, the numbers given here are likely to underestimate the population size, but comparisons between the soil type and treatments should be possible and valid (Bosse and Frenzel, 1997). The mean largest population of MOB in this study was recorded for rhizosphere soil followed by bulk and bare soil (Table 1). The differences due to soil were significant (F2,12 = 706.14; p<0.05) (Table 2). Lowest mean value for MOB population was estimated for urea treated soil (Table 1). Our results showed that MOB population size was significantly lower for fertilized soil as compared to control soil (F1,12 = 142.47; p<0.05).
| Table 3: | Cropping season averages (Mean±SE) for plant height (cm), Root biomass and shoot biomass in control and fertilized (0 and 100 kg N ha-1) planted to rainfed rice variety NDR-97 on six sampling dates (DAS = days after sowing) as affected by N fertilizers utilized |
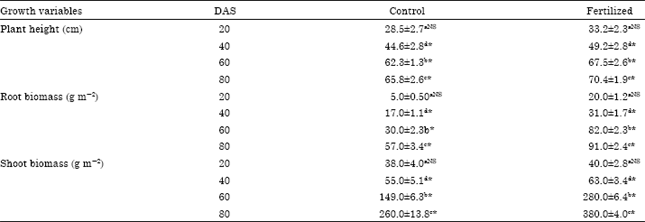 | |
| Data are expressed as mean±SE of three replicates in each treatments of vegetative control and fertilized plots. a20 DAS vs. 40 DAS, b20 DAS vs. 60 DAS, c20 vs. 80 DAS and 40 vs. 80 DAS showed pairwise comparison between days intervals. Values in a row bearing superscript * are significantly different and NS for not significant from each other at p<0.05 | |
DISCUSSION
Gilbert and Frenzel (1998) found that active CH4 oxidizing bacteria (MOB) occurred near to root mat similar to the dense root texture in the upper layer of rice fields. In the present dryland rice field, the MOB population was much higher in the bulk soil compared to the bare soil, indicating that the bulk soil was not entirely free from the influence of roots. The soil of dryland rice field also gets periodically saturated due to heavy rainfall events when CH4 emission instead of net consumption occurs (Singh et al., 1998, 1999a). The O2 supplying potential of plant roots is a major factor for the multiplication, growth and sustenance of methanotrophic bacteria in the rhizosphere. The aerenchymatous tissue of rice plant serves as a conduit to transport CH4 from the anoxic soils to the atmosphere (Mariko et al., 1991) and oxygen from the atmosphere to the rhizosphere (Frenzel et al., 1992). The supply of both CH4 and oxygen would thus be more favorable for the methanotroph population to develop in rhizosphere than in the bulk or bare soil. The view that supply of both CH4 and O2 is essential for methanotroph population is supported by the findings that the population size in paddy soils exposed to air enriched with 20% methane, increased to 2.3x107 cells g-1 in comparison to control soil (Bender and Conrad, 1992). Singh et al. (1998, 1999b) found that plant variables, especially plant height, root biomass and shoot biomass representing the conduit and ventilation effects were important for CH4 oxidation in dryland rice agriculture.
The application of urea resulted in a higher NH4+-N concentration, as a consequence of hydrolysis, which proceeds rapidly in warm, moist soils. The low NH4+-N in the rhizosphere soil evidently resulted from the continuous uptake by rice and uptake and oxidation by microorganisms such as ammonia oxidizers and MOB (Arth et al., 1998).
CONCLUSION
In conclusion, the development of the rhizosphere brings about a spatial pattern in the distribution of methanotrophic population, which increases in size during the vegetative period and within the rhizosphere and adjoining bulk soil as compared to the bare soil. Greater O2 availability due to ventilation by rice plants, lower concentrations of NH4+-N due to continuous plant uptake and a larger methanotroph population make the rice rhizosphere a microsite for intense CH4 oxidation activity. We thus demonstrate that plant, plant age and fertilization affect MOB in dryland rice field.
ACKNOWLEDGMENT
The authors express their sincere gratitude to the Head Department of Botany and Department of Agronomy, Institute of agricultural Sciences Banaras Hindu University, for providing laboratory facilities. We are also grateful to principal, U.P College, Varanasi for providing necessities of this work to pursue higher studies.
REFERENCES
- Adhya, T.K., K. Bharati, S.R. Mohanty, B. Ramakrishnan, V.R. Rao, N. Sethunathan and R. Wassmann, 2000. Methane emission from rice fields at Cuttack, India. Nutr. Cycl. Agroecosyst., 58: 95-105.
CrossRef - Arth, I., P. Frenzel and R. Conrad, 1998. Denitrification coupled to nitrification in the rhizosphere of rice. Soil Biol. Biochem., 30: 509-515.
CrossRef - Banik, A., M. Sen and S.P. Sen, 1996. Effects of inorganic fertilizers and micronutrients on methane production from wetland rice (Oryza sativa L.). Biol. Fertil. Soils, 21: 319-322.
CrossRef - Bender, M. and R. Conrad, 1992. Kinetics of CH4 oxidation in oxic soils exposed to ambient air or high CH4 mixing ratios. FEMS Microbiol. Lett., 101: 261-269.
CrossRefDirect Link - Bodelier, P.L.E., A.P. Hahn, I.R. Arth and P. Frenzel, 2000. Effects of ammonium-based fertilisation on microbial processes involved in methane emission from soils planted with rice. Biogeochemistry, 51: 225-257.
CrossRef - Bosse, U. and P. Frenzel, 1997. Activity and distribution of methane oxidizing bacteria in flooded rice soil microcosms and in rice plants (Oryza sativa). Applied Environ. Microbiol., 63: 1199-1207.
Direct Link - Claude, E.B., 1979. Determination of total ammonia nitrogen and chemical oxygen demand in fish culture systems. Trans. Am. Fisher. Soc., 108: 314-319.
Direct Link - Denier van der Gon, H.A.C. and H.U. Neue, 1996. Oxidation of methane in the rhizosphere of rice plants. Biol. Fertil. Soils, 22: 359-366.
CrossRef - Dubey, S.K. and J.S. Singh, 2000. Spatio-temporal variation and effect of urea fertilization on methanotrophs in a tropical dryland rice field. Soil Biol. Biochem., 32: 521-526.
CrossRef - Duxbury, J.M., 1994. The significance of agricultural sources of greenhouse gases. Fertilizer Res., 38: 151-163.
CrossRefDirect Link - Eller, G. and P. Frenzel, 2001. Changes in activity and community structure of methane oxidizing bacteria over the growth period of rice. Applied Environ. Microbiol., 67: 2395-2403.
PubMed - Eller, G., M. Kruger and P. Frenzel, 2005. Comparing field and microcosm experiments: A case study on methano and methylotrophic bacteria in paddy soil. FEMS Microbiol. Ecol., 51: 279-291.
PubMed - Frenzel, P. and U. Bosse, 1996. Methyl fluoride, an inhibitor of methane oxidation and methane production. FEMS Microbiol. Ecol., 21: 25-36.
CrossRef - Gilbert, B. and P. Frenzel, 1998. Rice roots and CH4 oxidation: The activity of bacteria, their distribution and the microenvironment. Soil Biol. Biochem., 30: 1903-1916.
CrossRef - Frenzel, P., F. Rothfuss and R. Conrad, 1992. Oxygen profiles and methane turnover in a flooded rice microcosm. Biol. Fertil. Soils, 14: 84-89.
CrossRef - Gilbert, B. and P. Frenzel, 1995. Methanotrophic bacteria in the rhizosphere of rice microcosms and their effect on pore water methane concentration and methane emission. Biol. Fertil. Soils, 20: 93-100.
CrossRef - Holmes, A.J., A. Costello, M.E. Lidstrom and J.C. Murrell, 1995. Evidence that particulate methane monooxygenase and ammonia monooxygenase may be evolutionarily related. Federation Eur. Microbiol. Soc. Microbiol. Lett., 132: 203-208.
CrossRef - Hutsch, B.W., K. Mengel and P. Russell, 1996. CH4 oxidation in two temperate arable soils as affected by nitrate and ammonium application. Biol. Fertil. Soils, 23: 86-92.
CrossRef - Henckel, T., U. Jackel and R. Conrad, 2001. Vertical distribution of the methanotrophic community after drainage of rice field soil. FEMS Microbiol. Ecol., 34: 279-291.
PubMed - King, G.M., 1994. Associations of methanotrophs with the roots and rhizomes of aquatic vegetation. Applied Environ. Microbiol., 60: 3220-3227.
Direct Link - Lee, K.K., S.P. Wani, K.L. Sahrawat, N. Trimurtulu and O. Ito, 1997. Nitrogen and/or phosphorus fertilization effects on organic carbon and mineral contents in the rhizosphere of field grown Sorghum. Soil Sci. Plant Nutr., 43: 117-126.
Direct Link - Mariko, S., Y. Harazano, N. Owa and I. Nouchi, 1991. Methane in flooded soil water and the emission through rice plants to atmosphere. Environ. Exp. Bot., 31: 343-350.
CrossRef - Neue, H.U., 1997. Fluxes of methane from rice fields and potential for mitigation. Soil Use Mgmt., 13: 258-267.
CrossRef - Rowe, R., R. Todd and J. Waide, 1977. Microtechnique for most-probable-number analysis. Applied Environ. Microbiol., 33: 675-680.
PubMedDirect Link - Singh, J.S. and S. Singh, 1995. Methanogenic bacteria, methanogenesis and methane emission from rice paddies. Trop. Ecol., 36: 145-165.
Direct Link - Singh, S., J.S. Singh and A.K. Kashyap, 1999. Methane flux from irrigated rice fields in relation to crop growth and N-fertilization. Soil Biol. Biochem., 31: 1219-1228.
CrossRef - Singh, S., J.S. Singh and A.K. Kashyap, 1999. Methane consumption by soils of dryland rice agriculture: Influence of varieties and N-fertilization. Chemosphere, 38: 175-189.
PubMed - Singh, S., J.S. Singh and A.K. Kashyap, 1998. Contrasting pattern of methane flux in rice agriculture. Naturwissen Schaften, 85: 494-497.
CrossRef - Watanabe, I., T. Hashimoto and A. Shimoyama, 1997. Methane-oxidizing activities and methanotrophic populations associated with wet land rice plants. Biol. Fertil. Soils, 24: 261-265.
CrossRef