
A. Joseph
Joseph Ayo Babalola University, P.M.B. 5006, Ilesa, Osun State, Nigeria
D.B. Olufolaji
Federal University of Technology, P.M.B. 704, Akure, Ondo State, Nigeria
F.E. Nwilene
Africa Rice Center (AfricaRice), P.M.B. 5320, Ibadan, Nigeria
A. Onasanya
Africa Rice Center (AfricaRice), P.O. Box 33581, Dares-Salaam, Tonzania
M.M. Omole
Joseph Ayo Babalola University, P.M.B. 5006, Ilesa, Osun State, Nigeria
R.O. Onasanya
Federal University of Technology, P.M.B. 704, Akure, Ondo State, Nigeria
Y. Sere
Africa Rice Center (AfricaRice), 01 BP 2031, Cotonou, Benin
International Journal of Agricultural Research
Year: 2011 | Volume: 6 | Issue: 9 | Page No.: 691-698







ABSTRACT
Chlorophyll reduction in rice leaves is a prominent feature of rice yellow mottle virus-infected plants. This research work was carried out to investigate the effect of rice yellow mottle disease on the chlorophyll content of eight differential rice cultivars screened for resistance to rice yellow mottle virus (RYMV) with vector transmission technique. Border row rice seedlings mechanically inoculated with RYMV isolate at 21 days after sowing metamorphosed into infected rows, following which the rice cultivars were sown 15 days after. Two days after sowing the test cultivars, life adults of three vectors (Oxya hyla, Locris rubra and Chnootriba similis) of rice yellow mottle virus were released onto the infected rows inside separate insect-proof screen house to acquire and transmit the virus to the test cultivars. The chlorophyll contents of the leaves were then measured at 42, 56 and 70 days after sowing. The result of the genotype by environment interaction showed that environment exerted the most profound effect on chlorophyll reduction (36.26%) in the rice cultivars screened with O. hyla while the least effect (5.87%) was recorded when the screening was carried out with C. similis. Findings from this research work showed that chlorophyll reduction varied among the rice cultivars at different stages of rice plant. Though, the photosynthetic rate of the rice leaves were not investigated, it could be deduced that this physiological function would invariably reduce in the rice cultivars.
PDF Abstract XML References Citation
Received: June 21, 2011;
Accepted: September 21, 2011;
Published: October 31, 2011
How to cite this article
A. Joseph, D.B. Olufolaji, F.E. Nwilene, A. Onasanya, M.M. Omole, R.O. Onasanya and Y. Sere, 2011. Genotype by Environment Interaction of Chlorophyll Reduction in Rice Cultivars Screened for Resistance to Rice Yellow Mottle Disease with Vector Transmission Method. International Journal of Agricultural Research, 6: 691-698.
DOI: 10.3923/ijar.2011.691.698
URL: https://scialert.net/abstract/?doi=ijar.2011.691.698
DOI: 10.3923/ijar.2011.691.698
URL: https://scialert.net/abstract/?doi=ijar.2011.691.698
INTRODUCTION
Rice yellow mottle virus (RYMV) remains the major biotic constraint to rice production in Africa and has the propensity to lead rice to the verge of extinction in the continent. The disease is transmitted through mechanical contact and by insect vectors (Nwilene et al., 2009). Insects are the first agent to convey the virus to rice fields and have therefore been recommended in screening for resistance to rice yellow mottle disease (Sere et al., 2008; Nwilene et al., 2009; Joseph et al., 2009).
The disease is characterized by yellowing and mottling of leaves and stunting of infected plants (Reckhaus and Andriamasintseheno, 1995; Nwilene et al., 2009; Ndikumana et al., 2011; Ochola and Tusiime, 2011). Partial emergence of panicles results in infected plants, with spikelets bearing sterile or unfilled grains (Thottappilly and Rossel, 1993). Losses in yield arising from RYMV vary from 54-97% (Reckhaus and Andriamasintseheno, 1995).
Plants infected with virus often show symptoms such as chlorosis, resulting from changes in the structure and function of the chloroplast (Ryslava et al., 2003). Infection of plants by virus leads to a reduction in chloroplast in the affected leaf areas (Balachandran et al., 1994) and this is usually accompanied by concomitant reduction in photosynthesis (Rahoutei et al., 2000). Rice plant is composed of leaves which differ in age (Ramesh et al., 2002). These leaves vary in contribution to the growth of the whole plant and the resulting grain yield. Biological interactions between rice plant and RYMV result in alteration of physiological processes of the host, particularly photosynthesis (Mandal et al., 2009). The virus prevents the host from maximum utilization of nutrients required for optimum growth, development and yield (Saravanan et al., 2010). Leaf chlorophyll content is the major determinant of photosynthetic capacity, contributing substantially to grain yield (Ramesh et al., 2002). This research work was therefore carried out to investigate the genotype by environment interaction of RYMV on the chlorophyll content of eight rice cultivars screened for resistance with insect vectors.
MATERIALS AND METHODS
Study site: The research work was carried out in the screen house at Africa Rice Centre (Nigeria Station), Ibadan, between April 2008 and October 2009.
Isolate used: The RYMV isolate characterised as pathotype one, recommended for vector-rice screening experiment (Sere et al., 2008; Nwilene et al., 2009) was used for the study. The isolate was obtained from infected plants at the Plant Pathology Unit of Africa Rice Center, Cotonou, Republic of Benin.
Insect vectors used: Life adult populations of Oxya hyla, Locris rubra and Chnootriba similis used for the trial were collected with a sweep net from Africa Rice Center rice fields at the International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria and were caged in a screen house to feed on cowpea plants for three weeks in order to lose RYMV infectivity.
Rice cultivars evaluated: The eight differential rice cultivars evaluated (Table 1) were obtained from AfricaRice Plant Pathology Unit, Cotonou.
Isolate propagation: The isolate was first propagated on BG 90-2 (a highly susceptible cultivar), following mechanical inoculation of 21-day-old rice seedlings in the screen house. Four weeks after inoculation, leaves showing typical yellow mottle symptoms characteristic of RYMV infection were harvested and used to prepare the viral inoculum. The inoculum was prepared by grinding the infected leaf samples in 0.01 M phosphate buffer (pH 7) in the ratio of 1:10 weight by volume and the resulting homogenate filtered through cheesecloth. Carborundum powder (600-mesh) was added to the inoculum to aid the penetration of the virus into leaf tissues.
Planting and inoculation of border rows: Seeds of BG90-2 were sown in plastic pots as border rows and the seedlings were mechanically inoculated with the viral suspension at three weeks after sowing. The inoculated rice seedlings metamorphosed to infected rows.
| Table 1: | Identity of rice cultivars evaluated |
 | |
Planting of test cultivars: The test cultivars were planted at two weeks after inoculating the border rows. Two plastic pots of 20 cm diameter, 19.5 cm height and 5 L capacity, filled with 2.5 kg of sterile sandy-loam soil were used for sowing each cultivar. The treatments comprised of eight rice cultivars sown at a distance of 0.5 m from the infected rows in each screen house. The experiment was laid out in a Randomized Complete Block Design (RCBD) with 3 replicates. Nine seeds (three seeds per hill and three hills per pot) of each cultivar were sown at a depth of 2 cm. The plants were later thinned to three seedlings per pot at one week after emergence. Two days after planting the test cultivars, 120 adults each of non-viruliferous O. hyla, L. rubra and C. similis were separately introduced onto the infected rows in each screen house to feed, acquire and transmit the virus into the test cultivars.
In the control experiment, the procedures of planting border row rice plants and test cultivars were carried out in a separate screen house. However, the border row rice seedlings were left un-inoculated. The vectors were then released unto the rice plants.
Data collection and analysis: The chlorophyll contents of the leaves were measured at 42, 56 and 70 days after sowing using a SPAD 502 chlorophyll meter (Monje and Bugbee, 1992; Martines and Guiamet, 2004). SPAD-502 meter determines the relative amount of chlorophyll present in the leaves by measuring the absorbance of the leaf in two wavelength regions. The genotype by environment interaction effect of variety by chlorophyll reduction was performed using additive main effects and multiplicative interaction (AMMI) analysis (Ebdon and Gauch, 2002). The AMMI analysis combines analysis of variance and principal component (PC) analysis into a single model with additive and multiplicative parameters. All analyses and drawing of figures were carried out using the CropStat 7.2 software packages.
RESULTS AND DISCUSSION
The results showed considerable variations in variety by chlorophyll content among the rice cultivars. In Fig. 1, chlorophyll reductions were statistically similar in Bouake 189, ITA 235 and H232-44-1-1 cultivars at 42 DAS. At 56 DAS, there was no significant difference in chlorophyll reduction between Moroberekan and PNA647F4-56. Similarly, chlorophyll reductions were not significantly different among Gigante, FARO 11, LAC 23, ITA 235 and H232-44-1-1 at 70 DAS.
In Fig. 2, chlorophyll reductions were not significantly different among Gigante, FARO 11, Moroberekan and PNA647F4-56 at 42 DAS. At 56 DAS, no significant difference was observed in chlorophyll reduction among Bouake 189, FARO 11, Moroberekan, LAC 23, ITA 235 and PNA647F4-56. Leaf chlorophyll contents were observed to be statistically similar among Gigante, Bouake 189, LAC 23 and ITA 235 at 70 DAS.
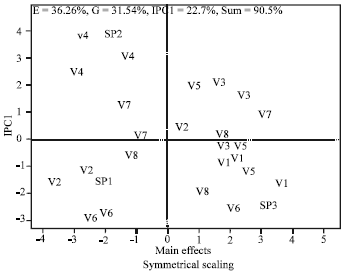 | |
| Fig. 1: | Genotype (variety) by environment (chlorophyll content) interaction effects with O. hyla using Additive Main effects and Multiplication Interaction (AMMI) analysis. Varieties V1 = Gigante, V2 = Bouake 189; V3 = FARO 11; V4 = Moroberekan; V5 = LAC 23; V6 = ITA 235; V7 = PNA 647F4-56; V8 = H 232-44-1-1; SP1, SP2 and SP3 = Leaf chlorophyll content at 42, 56 and 70 days after sowing |
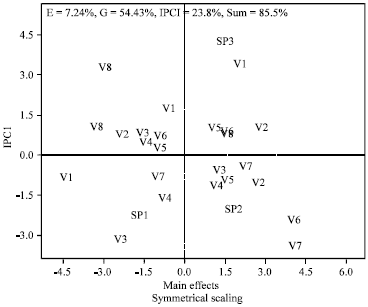 | |
| Fig. 2: | Genotype (variety) by environment (chlorophyll content) interaction effects with L. rubra using Additive Main effects and Multiplication Interaction (AMMI) Analysis. Varieties V1 = Gigante, V2 = Bouake 189; V3 = FARO 11; V4 = Moroberekan; V5 = LAC 23; V6 = ITA 235; V7 = PNA 647F4-56; V8 = H 232-44-1-1; SP1, SP2 and SP3 = Leaf chlorophyll content at 42, 56 and 70 days after sowing |
Figure 3 showed that the there was no significant difference in chlorophyll reductions in the rice leaves among Bouake 189, FARO 11, ITA 235, PNA647F4-56 and H232- 44-1-1 cultivars at 42 such as RYMV, that causes a reduction in the chlorophyll content of the leaves would invariably have a negative impact on the photosynthetic capacity of rice plant. Although, the present study DAS.
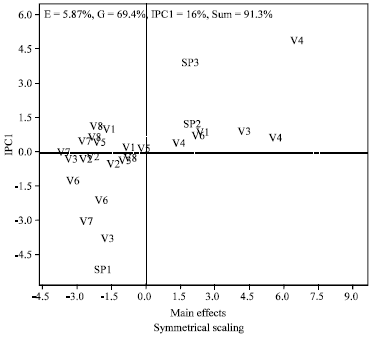 | |
| Fig. 3: | Genotype (variety) by environment (chlorophyll content) interaction effects with C. similis using Additive Main effects and Multiplication Interaction (AMMI) Analysis. Varieties: V1 = Gigante, V2 = Bouake 189; V3 = FARO 11; V4 = Moroberekan; V5 = LAC 23; V6 = ITA 235; V7 = PNA 647F4-56; V8 = H 232-44-1-1; SP1, SP2 and SP3 = Leaf chlorophyll content at 42, 56 and 70 days after sowing |
At 56 and 70 DAS, the chlorophyll content of Gigante, FARO 11, Moroberekan and ITA 235 were statistically similar (Fig. 3).
Environment exerted the most profound effect on chlorophyll reduction (36.26%) in the rice cultivars screened with O. hyla (Fig. 1) while the least effect of environment on chlorophyll reduction (5.87%) was recorded when the screening of the rice cultivars was carried out with C. similis (Fig. 3). In Fig. 2, environment contributed 7.24% to chlorophyll reduction in the evaluated rice cultivars.
Plants, by virtue of their green pigment (chlorophyll) have the ability to synthesize complex carbohydrates. This process is the foundation of dry matter production in plants. Thus, any factor, did not proceed to assess the photosynthetic rates of the rice cultivars evaluated for chlorophyll reductions, earlier studies confirmed a general reduction in the photosynthetic capacities of crop plants infected with pathogen and these effects were positively correlated with chlorophyll reduction in the leaves of such infected crops.
Reduction in chlorophyll content due to microbial pathogens is almost universal in majority of plants. Shtienberg (1992) reported that the pattern and amount of photosynthetic rates in annual and perennial crops infected with foliar diseases is a function of the type of disease and the causative organism. Akter et al. (2004) reported photosynthetic capacity as a function of chlorophyll content of rice leaves. Similarly, Poshtmasari et al. (2007) observed a positive correlation between leaf chlorophyll content and rice yield. Ninety percent of the grain yield in rice originates from photosynthetic activity of leaves (Xie et al., 2011). As such, reduction in photosynthetic rate would have a negative effect on rice productivity, resulting in significant yield losses. Reduction in net photosynthetic rates in some crops due to pathogens is well documented in literature. Results from this study agrees with the findings of Bassanezi et al. (2002), who reported a reduction in net photosynthetic rate in mung beans infected with rust, angular leaf spot and anthracnose, owing to reduction in leaf chlorophyll content. This observation is also consistent with the findings of Maust et al. (2003), who reported a decrease in photosynthesis in coconut palm trees infected with lethal yellowing due to chlorophyll reduction in the leaves. Similarly, the observation is in agreement with the report of Ryslava et al. (2003), who showed that potato virus Y (PVY) affected photosynthesis negatively in the leaves of Nicotiana tabacum L., attributing the decrease to a reduction in chlorophyll content. However, findings from this study is in partial agreement with the observation of Ingram (1981) and Lindenthal et al. (2005), who reported that chlorophyll reduction alone does not wholly account for photosynthetic net rate in crops infected with downy mildew disease.
Varying percentage chlorophyll reduction have also been observed in several crops: 39% in pea plants affected by Mycosphaerella pinodes; 10-48% in oak infected with Microsphaera alphitoides; 36% in poplar infected with Melampsora tremulae; 63% in Cirsium arvense infected with Puccinia suaveolens and 32-64% in early rose potatoes infected with Xanthomonas vesicatoria (Kupeevicz, 1947). In the instances outlined above, Kupeevicz (1947) observed decrease in photosynthetic efficiencies of the crops which he attributed to chlorophyll reductions. Similarly, Krishnamani and Lakshmanan (1976) showed that cotton cultivars infected with Fusarium vasinfectum showed a reduced rate of photosynthesis, owing to a reduction in total chlorophyll content of infected leaves.
Plant age could have a significant effect on chlorophyll reduction. In the present study, there were variations in chlorophyll content of the leaves of the differential rice cultivars at various stages of rice development.
CONCLUSION
Findings from this research work showed that chlorophyll reduction varied among the rice cultivars at different stages of rice plant. Though, the photosynthetic rate of the rice leaves were not investigated, it could be deduced that this physiological function would invariably reduce in the rice cultivars based on the findings of earlier researchers.
ACKNOWLEDGMENT
The authors are very grateful to the Government of Japan (Ministry of Foreign Affairs) for funding this research work. We also wish to acknowledge the technical support of Mr. Bayo Kehinde.
REFERENCES
- Balachandran, S., C.B. Osmond and P.F. Daley, 1994. Diagnosis of the earliest strain-specific interactions between tobacco mosaic virus and chloroplasts of tobacco leaves in vivo by means of chlorophyll fluorescence imaging. Plant Physiol., 104: 1059-1065.
Direct Link - Bassanezi, R.B., L. Amorim, A.B. Filho and R.D. Berger, 2002. Gas exchange and emission of chlorophyll fluorescence during the monocycle of rust, angular leaf spot and anthracnose on bean leaves as a function of their trophic characteristics. J. Phytopathol., 150: 37-47.
CrossRefDirect Link - Ebdon, J.S. and H.G. Gauch, 2002. Additive main effect and multiplicative interaction analysis of National Turfgrass performance trials. Crop Sci., 42: 489-496.
CrossRefDirect Link - Krishnamani, M.R.S. and M. Lakshmanan, 1976. Photosynthetic changes in Fusarium-infected cotton. Can. J. Bot., 54: 1257-1263.
CrossRefDirect Link - Lindenthal, M., U. Steiner, H.W. Dehne and E.C. Oerke, 2005. Effect of downy mildew development on transpiration of cucumber leaves visualized by digital infrared thermography. Phytopathology, 95: 233-240.
PubMed - Mandal, K., R. Saravanan, S. Maiti and I.L. Kothari, 2009. Effect of downy mildew disease on photosynthesis and chlorophyll fluorescence in Plantago ovata Forsk. J. Plant Dis. Protect., 116: 164-168.
Direct Link - Maust, B.E., F. Espadas, C. Talavera, M. Aguilar, J.M. Santamaria and C. Oropeza, 2003. Changes in carbohydrate metabolism in coconut palms infected with the lethal yellowing phytoplasma. Phytopathology, 93: 976-981.
PubMed - Monje, O.A. and B. Bugbee, 1992. Inherent limitations of non-destructive chlorphyll meters: A comparison of two meters. Hort. Sci., 27: 69-71.
PubMed - Ndikumana, I., R. Gasore, S. Issaka, G. Pinel-Galzi and A. Onasanya et al., 2011. Rice yellow mottle virus in rice in Rwanda: First report and evidence of strain circulation. New Dis. Rep., 23: 18-18.
CrossRefDirect Link - Poshtmasari, H.K., H. Pirdashti, M. Nasiri and M.A. Bahmanyar, 2007. Chlorophyll content and biological yield of modern and old rice cultivars in different urea fertilizer rates and applications. Asian J. Plant Sci., 6: 177-180.
CrossRefDirect Link - Rahoutei, J., I. García-Luque and M. Baron, 2000. Inhibition of photosynthesis by viral infection: Effect on PSII structure and function. Physiol. Plantarum, 110: 286-292.
CrossRef - Ochola, D. and G. Tusiime, 2011. Survey on incidence and severity of rice yellow mottle virus disease in Eastern Uganda. Int. J. Plant Pathol., 2: 15-25.
CrossRefDirect Link - Ramesh, K., B. Chandrasekaran, T.N. Balasubramanian, U. Bangarusamy, R. Sivasamy and N. Sankaran, 2002. Chlorophyll dynamics in rice (Oryza sativa) before and after flowering based on SPAD (chlorophyll) meter monitoring and its relation with grain yield. J. Agron. Crop Sci., 188: 102-105.
CrossRefDirect Link - Ryslava, H., K. Muller, S. Semo radova, H. Synkova and N. Eeoovska, 2003. Photosynthesis and activity of phosphoenolpyruvate carboxylase in Nicotiana tabacum L. leaves infected by potato virus A and potato virus. Photosynthetica, 41: 357-363.
Direct Link - Sere, Y., A. Onasanya, F.E. Nwilene, M.E. Abo and K. Akator, 2008. Potential of insect vector screening method for development of durable resistant cultivars to rice yellow mottle virus disease. Int. J. Virol., 4: 41-47.
CrossRefDirect Link - Shtienberg, D., 1992. Effects of foliar diseases on gas exchange processes: A comparative study. Phytopathology, 82: 760-765.
CrossRefDirect Link - Xie, X.J., S.H.H. Shen, Y.X. Li, X.Y. Xao, B.B. Li and D.F. Xu, 2011. Effect of photosynthetic characteristic and dry matter accumulation of rice under high temperature at heading stage. Afr. J. Agric. Res., 6: 1931-1940.
Direct Link - Nwilene, F.E., A.K. Traore, A.N. Asidi, Y. Sere, A. Onasanya and M.E. Abo, 2009. New records of insect vectors of Rice Yellow Mottle Virus (RYMV) in Cote d'Ivoire, West Africa. J. Entomol., 6: 189-197.
CrossRefDirect Link - Martines, D.E. and J.J. Guiamet, 2004. Distortion of the SPAD 502 chlorophyll meter readings by changes in irradiance and leaf water status. Agronomie, 24: 41-46.
CrossRefDirect Link