
M.A. El-Siddig
Depertment of Botany, Faculty of Science, University of Khartoum, P.O. Box 321, P.C. 11115, Sudan
A.A. El-Hussein
Depertment of Botany, Faculty of Science, University of Khartoum, P.O. Box 321, P.C. 11115, Sudan
M.M. Saker
Academy of Scientific Research and Technology, Egypt
International Journal of Agricultural Research
Year: 2011 | Volume: 6 | Issue: 4 | Page No.: 323-334







ABSTRACT
The aim of this study was to develop a protocol for transformation/regeneration of tomato plant (cv. Summer set) using a plant antifungal gene (defensin). Transformation was carried out using disarmed A. tumefaciens strain LBA4404 harbouring a binary vector pITB-AFP. Sequencing of plasmid DNA extracted from this strain indicated that it contains defesin gene (AFP), an antifungal protein-coding gene, under the control of a CaMV 35S promoter and Nopaline Synthase (NOS) terminator, hygromycin phosphotransferase (hpt) and β-glucuronidase (GUS) genes, as selectable and marker genes, respectively. The factors that affect transformation/regeneration protocols were optimized in a series of experiments. Results indicated that exposure of cotyledonary explants to Agrobacterium inoculums of 0.8 O.D600 for 30 min, selection on Hygromycin-containing medium (Hygromycin concentration of 25 mg L-1) and subsequent regeneration on MS medium supplemented with 2.5 mg L-1 BA as a cytokinin and 1.0 mg L-1 IAA as an auxin resulted in transformation efficiency of 7%. GUS expression was observed in transformed tomato shoots but never in the control plants. PCR amplification of DNA extracted from the transformed tissues demonstrated the generation of the expected amplicon, corresponding to AFP gene. This result strongly verifies the successful transformation of the tomato cultivar Summer set, an endeavour which is reported for the first time in Sudan. Moreover, this protocol paves the way for problem solving-applications encompassing other Sudanese crops of economic importance.
PDF Abstract XML References Citation
Received: February 15, 2011;
Accepted: April 13, 2011;
Published: May 21, 2011
How to cite this article
M.A. El-Siddig, A.A. El-Hussein and M.M. Saker, 2011. Agrobacterium-mediated Transformation of Tomato Plants Expressing Defensin Gene. International Journal of Agricultural Research, 6: 323-334.
DOI: 10.3923/ijar.2011.323.334
URL: https://scialert.net/abstract/?doi=ijar.2011.323.334
DOI: 10.3923/ijar.2011.323.334
URL: https://scialert.net/abstract/?doi=ijar.2011.323.334
INTRODUCTION
Tomato (Lycopersicon esulentum Mill.) is the major vegetable crop grown worldwide, with a production estimates of 95 million Mt (FAOSTAT, 2002) and its production is concentrated in semi-arid regions (Santa-Curz et al., 2002). Presently, tomato is becoming increasingly important in Sudan for local consumption and for export. It is cultivated throughout the year under irrigation in an area that exceeds 36540 hectares with an average yield of 17.57 tons per hectare (AOAD, 2007). The most important grown cultivars are the canning types such as Strain B, Strain C, Summer set, Peto86, Peto111 and CastleRock in addition to few local varieties (Mukhtar et al., 2009).
In Sudan, as in other parts of the world, cultivated tomatoes suffer from many diseases that are caused by viruses, bacteria and fungi. Among virus diseases, tomato mosaic root knot and tomato yellow leaf curl (Geneif, 1986) are considered as the most important. Bacterial diseases include bacterial wilt (Pseudomonas solanacearum) (Jinnah et al., 2002), Verticillium wilts (Verticillium dahliae), powdery mildews (Leveilula taurica) and early and late blights which are caused by Alternaria solani/alternata and Phytophthora infestans, respectively (Abbo et al., 2009).
While the use of disease resistant cultivars may present an effective way of controlling the above diseases, genetic engineering techniques continue to play a major role in the development of disease resistant cultivars (Bhatia et al., 2004). Various factors that affect the development of techniques for the isolation and identification of many genes involved in plant disease resistance, morphology and development have been studied (Ling et al., 1998). In addition, different factors such as Agrobacterium cell density (Murray et al., 1998), regeneration and co-cultivation conditions (Hu and Phillips, 2001), addition of acetosyringone and cell competence after wounding (Murray et al., 1998) and gene constructs (Krasnyanski et al., 2001) were found to play a major role in tomato transformation. The first report of tomato transformation was forwarded by McCormick et al. (1986) and since then, there have been numerous publications on transformation of various tomato cultivars (Vidya et al., 2000; Hu and Phillips, 2001; Raj et al., 2005; Sun et al., 2006; Shahriari et al., 2006; Saker et al., 2007). Standardization of tomato transformation procedures is, still, incomplete as different tomato cultivars vary in their response to specific treatment. The present study was undertaken to develop an efficient procedure for the production of fungal-resistant transgenic tomato (cv. CastleRock) plants expressing defensin gene.
MATERIALS AND METHODS
This study was conducted during June 2009 to Jan. 2010. Agrobacterium-mediated tomato transformation was performed in accordance to the protocol of Park et al. (2003) with some modifications.
Plant tissue: Seeds of tomato (Lycopersicon esculentum Mill.) cultivar Summer set were surface sterilized in 10% (v/v) sodium hypochlorite solution, with 2 drops of Tween-20, for 15 min followed by three rinses in sterile water. Seeds were then germinated on MS (Duchefa Biochemie) inorganic salt (MSO) medium (Murashige and Skoog, 1962) with 30 g L-1 sucrose, pH 5.7 and solidified using 7 g L-1 TC agar (GFS Chemicals and reagents, Texsas, USA). Hypocotyl explants and cotyledonary sections either with distal or proximal ends from 8-day-old seedlings were used. Explants sub-cuttings made in MSO liquid medium were blot-dried and placed on MS medium supplemented with 1 mg L-1 of both 6-benzyladenine (BA) and Naphthalene Acetic Acid (NAA) for one day before co-cultivation with Agrobacterium.
Bacterial strain and plasmid: A defensin (antifungal, AFP) gene cloned in Agrobacterium tumefaciens LBA4404 strain was used. This strain harbours the pITB-AFP plasmid vector which contains defensin (AFP) gene under the transcriptional control of cauliflower mosaic virus 35S promoter (CaMV-35S) and nopaline synthase, Nos, terminator, hygromycin phosphotransferase (hpt) gene and GUS-intron (uidA-β-glucuronidase) as a reporter gene. Bacteria were grown on LB medium supplemented with streptomycin (30 mg L-1) and kanamycin (100 mg L-1) with shaking at 200 rpm to an OD600 = 1.0. The bacterial suspension was then diluted with MSO medium in a ratio of 1: 5 and used for transformation experiments.
Transformation and regeneration protocol: The precultured explants were carefully submerged in an Agrobacterium inoculum in a Petri dish (Ø9 cm) for 30 min with gentle swinging.
The explants were then blot dried on sterile filter paper, transferred to Medium B and incubated in the dark for three days at 25°C. Explants were then transferred to plates containing a selection medium (Medium C) supplemented with 25 mg L-1 hygromycin. The plates were sealed with parafilm and explants were left to regenerate at 25°C, with a 16/8 h (light/dark) photoperiod in the culture room. Hygromycin-resistant calli obtained after a second round of selection were transferred to a fresh selection medium. Once in every two weeks, the fresh and healthy looking hygromycin-resistant calli were sub-cultured in a fresh selection medium for shoot regeneration. After approximately 6-8 weeks, shoots were excised and transferred to Medium D for shoot development. The shoots obtained were transferred to rooting medium (Medium E) for root development. A set of explants which was not co-cultivated with Agrobacterium was also prepared, as described above, as a negative control. The experiment was made in three replications in a completely randomized design. The number of surviving calli in each case was determined and the data was subjected to analysis of variance. Transformation frequency was expressed as a percentage of the number of shoots recovered from hygromycin-resistant calli relative to the total number of incubated hygromycin-resistent calli. Any rooting shoot on the selection medium containing 25 mg L-1 hygromycin was considered as a transformant.
Media composition: The culture media used in the transformation and regeneration experiments were solidified with 7g L-1 TC agar and its pH was adjusted to 5.7.
Medium B (Co-cultivation medium): MS salts and vitamins, 30 g L-1 sucrose, 1 mg L-1 BAP and 1 mg L-1 NAA.
Medium C (Selection and shoot regeneration medium): MS salts and vitamins, 30 g L-1 sucrose, 2.5 mg L-1 BAP, 0.1 mg L-1 Lndole-3-Acetic Acid (IAA), 250 mg L-1 cefotaxime and 25 mg L-1 hygromycin.
Medium D (Shoot eleongation medium): MS salts and vitamins, 30 g L-1 sucrose, 1 mg L-1 BAP, 0.1 mg L-1 IAA and 250 mg L-1 cefotaxime.
Medium E (Rooting medium): MS salts and vitamins, 30 g L-1 sucrose, 1 mg L-1 IAA and 250 mg L-1 cefotaxime.
Effect of Cefotaxime on callus growth and shoot regeneration: To examine the influence of cefotaxime on callus growth and shoot regeneration (before and after transformation), 100 cotyledonary explants (25 pieces per Petri dish) were separately cultured on MS medium containing 0, 50, 100, 200, 250 and 300 mg L-1 cefotaxime. Regeneration frequency was calculated as a percentage of the number of regenerated explants/total number of explants (Ling et al., 1998).
Effect of the inoculation period: To assess the effect of different periods of transfection (the period for which explants were immersed in the bacterial suspension) on the transformation frequency, explants were submerged in the bacterial broth for 10, 20, 30 and 35 min.
Effect of Agrobacterium density: Bacterial densities (OD600) ranging from 0.3 to 2.0 were tested to determine the optimum density for transformation.
Sensitivity of tomato explants to hygromycin: To determine the effect of hygromycin (water solution of hygromycin B) on the regeneration of tomato explants and to screen for an appropriate hygromycin concentration for transformants selection, cotyledonary and hypocotyl explants were transferred to the induction medium supplemented with different concentrations of hygromycin (0, 5, 10, 15, 20, 25, 30, 35 and 40 mg L-1) in Petri dishes (10 cm in diameter) each containing 100 explants. The explants were incubated for 2 weeks in the dark at 25°C and then transferred to a 16/8 h (light/dark) photoperiod of 80 mol m-2 sec-1 at 25°C. The number of regenerated shoots was recorded after 5 weeks.
Effect of plant growth hormones combination on regeneration: To evaluate the effect of growth regulators on transformation efficiency, different plant growth regulator combinations were examined. These combinations were in mg L-1: BA 1.0/NAA 0.1 for 4 days (a one-day preculture period and a 3-day co-cultivation period) followed by transfer to BA 2.5/IAA 0.1; Zeatin 1.0/IAA 0.1; Zeatin 2.0; or to Zeatin 2.0/IAA 0.1.
Histology and histochemical analysis: GUS activity was analyzed in putative transformed primary explants, shoots and leaves from regenerated plants according to Jefferson (1987). Plant cells were incubated at 37°C for 24 h in GUS-staining solution (0.5 mM of X-gluc, 10 mM EDTA, 0.5 mM ferricyanide, 0.5 mM ferrocyanide, in 0.1 M phosphate buffer, pH 7.0). GUS activity was visualized under stereoscope after washing the stained explants in 70% ethanol.
Molecular verification of the transformants: To confirm the presence of the AFP gene in the regenerated transformed plants, total DNAs were isolated from both transformed and untransformed (negative control) plant samples and were used as templates for PCR. Plasmid DNA was also isolated and used as a positive control.
Rapid plant DNA extraction: DNA extraction was carried out following CTAB (cetyl trimethyl ammonium bromide) protocol, which is a modification of the method of Doyle and Doyle (1987).
Polymerase Chain Reaction (PCR): The presence of AFP gene was investigated by PCR amplification. Specific oligonucleotide primers for AFP gene were ‘forward’: 5'- CGC GGA TCC ATG GCG AGG TGT GAG AAT TTG GCT-3' and ‘reverse’: 5'-TGC TCT AGA ATG GCG AGG TGT GAG AAT TTG GCT-3'.
Each PCR reaction was performed in 25 μL (total volume) of the reaction mixture that consisted of 1X reaction buffer, 10 ng plant DNA from a putative transgenic plant as a template, 200 mM dNTPs, 1.5 mM MgCl2, 2 mM of each primer and 0.5 unit of Taq DNA polymerase. Amplification was carried out in a Thermal Cycler (Biometra) under the following conditions: 94°C for 3 min for initial denaturation, 94°C for 1 min for denaturation, 54°C for 1.5 min for annealing, 72°C for 1 min for elongation, 7 min at 72°C final extension and 35 cycles of amplification.
Amplified DNA fragments were electrophoresed on 1.0% agarose gel and detected by ethidium bromide staining and photographed under ultraviolet light. The presence of the target band in the transformants and its absence in the untransformed plants is considered as a proof of successful transformation.
RESULTS
A total of 300 cotyledonary leaves and 300 hypocotyls of tomato plants (cv. Summer Set) were used for transformation. Trials revealed great variations in the transformation frequency depending on the explant type (cotyledon or hypocotyl). The results showed that cotyledonary explants were more efficient for regeneration and subsequent transformation than hypocotyl explants (Table 1). In both explant types, callus formation was observed in more than 85% of the cultured plant tissues within 15-25 days (Fig. 1).
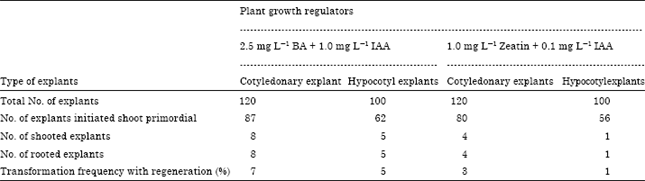
Detection of transformed cells was made, after 5 days of co-cultivating the explants with Agrobacterium, by placing individual explant pieces on Hygromycin-containing medium. Green calli were obtained, on average, after 4 weeks of incubation (Fig. 2). 47% and 35% of cotyledonary and hypocotyl explants, respectively, showed Hygromycin resistance after three selections. Transformed tissues were maintained on selection media until the appropriate size for transfer to shoot induction medium. Shoot induction was achieved in 6-8 weeks during which explants were transferred to fresh medium every two weeks. Well-grown shoots were cut off, vertically inserted into root induction medium and incubated for 10 days. A representative result is given in Fig. 3. The highest transformation frequencies were 7 and 3% for cotyledonary explants in media containing BA and Zeatin, respectively (Table 2). The results of survival of transformed and untransformed plantlets at various stages are summarized in Table 3.
| Table 1: | Effect of explants type on the rate of transformation |
 | |
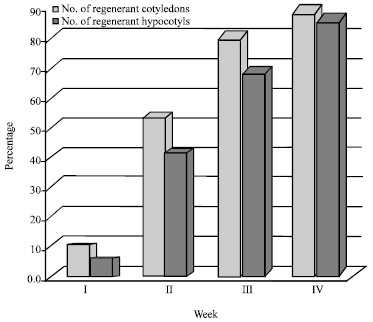 | |
| Fig. 1: | Percentages of regenerant tomato explants during 4 weeks |
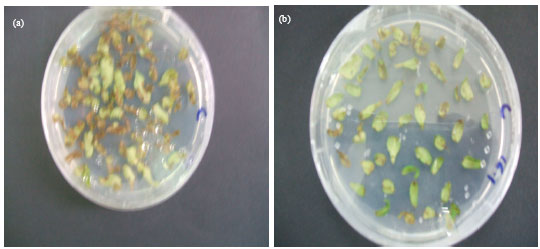 | |
| Fig. 2: | Recovery of Hygromycin resistant transgenic tomato explants on Hygromycin-containing medium. (a) Transformed cotyledonary explants culture on a hygromycin-containing medium and (b) transformed cotyledonary and hypocotyl explants cultured on a medium without hygromycin (note the browning of the untransformed tissues in between the resistant transformed tissues) |
 | |
| Fig. 3: | Various stages during regeneration of transgenic tomato plantlets: (a) callus induction and shoot initiation, (b) shoot elongation and (c) rooted plantlets |
| Table 2: | Effect of explants type and growth regulators on the transformation frequency of tomato |
 | |
| Table 3: | Survival percentage* of transformed (co-cultivated) and untransformed (control) tomato plants at various stages of development |
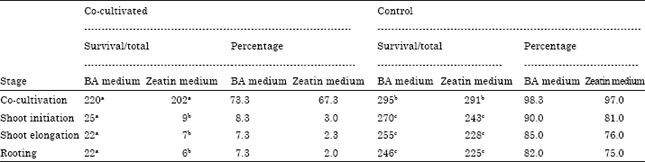 | |
| *Results based on pooled data of three experiments. Any two readings of the same stage having the same alphabetical letter are not significant at 0.05 level | |
The frequency of shoot initiation for transformed plantlets was found to be 2.0% (6/300) in the medium containing Zeatin and 7.3% (22/300) in the BA medium. For untransformed plantlets, shoot initiation frequency was 75.0% and 82.0% in Zeatin and BA media, respectively. The shoots were subsequently transferred on fresh selection medium containing BA for shoot elongation. Survival of the transformed shoots during shoot elongation period indicated successful transformation.
Verification of the transgenic plants
GUS histochemical assay: The histochemical GUS activity of selected regenerants showed a uniform dark blue color in the transformed tissues. GUS expression should also mimic the normal expression pattern of the gene of interest. On the other hand, non-transformed tissue did not exhibit such a color under identical assay conditions. GUS expression was found in some parts of the cotyledonary leaves and the hypocotyl pieces either at high (dark blue) or low levels (light blue). The possibility of false GUS positive was ruled out because the GUS gene was interrupted by an intron.
DNA isolation and PCR analysis: Genomic DNA from six randomly selected transformed plants as well as from untransformed ones were isolated and subjected to PCR analysis. The putative transgenic plants, when analysed by PCR using AFP-specific primers, gave the expected PCR amplicon (300 bp), whereas such an amplicon was not observed in untransformed (negative control) plants (Fig. 4). This result clearly demonstrates the presence of the AFP gene in tomato planlets co-cultivated with Agrobacterium containing the AFP-disarmed plasmid construct.
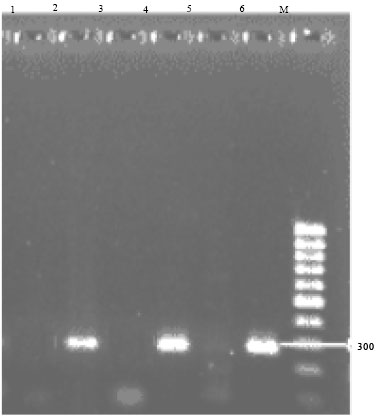 | |
| Fig. 4: | PCR confirmations of regenerated transformed plants using AFP primers. Lanes: 1: Negative control; 2 and 6: Transformed plants; 4: Positive control; 3 and 5: Untransformed plants; M: 1 kb marker |
DISCUSSION
Transformation of variuos tomato cultivars was previously reported by various authors. Transformation frequencies have ranged from 6% in cv. Pusa Ruby (Vidya et al., 2000) to 40% in cv. Micro-Tom (Sun et al., 2006). In spite of reported successes in tomato transformations, most of the transformation procedures were cumbersome and relied on feeder layers (petunia, tomato, or tobacco), time consuming media formulations or successive subcultures (Qiu et al., 2007).
No simple general procedure for tomato transformation exists due to many unknown factors that affect transformation and/or regeneration efficiencies. It has been reported that Agrobacterium tumefaciens-mediated gene transfer is influenced by strain (Stomp et al., 1990), genotype response to specific treatments (Park et al., 2003), explant type (Humara et al., 1999), temperature (Dillen et al., 1997), size of T-DNA (Park et al., 2003) and constitutive expression of the virulence genes (Hansen et al., 1994; Rossi et al., 1996). Van Roekel et al. (1993) reported significant effects of culture media and Agrobacterium strains on the transformation frequency of tomato. High-frequency transformation using A. tumefaciens depends not only on the efficiency of the plant in vitro regeneration system but also on the subsequent elimination of bacterial cells from transformed tissues (Tang et al., 2004).
A reliable plant regeneration protocol determines the successfulness of any transformation trial. Therefore, plant regeneration conditions have to be optimized for a given plant species/cultivar and type of explant (Farid Uddin et al., 2004). Results on regeneration trials indicated that callus induction and regeneration from tomato explants are quite permissive over a range of plant growth regulators. Previuos studies (McCormick et al., 1986; Van Roekel et al., 1993; Frary and Earle, 1996; Costa et al., 2000) have suggested specific types and combinations of plant growth regulators for effective regeneration of transformed tomato plants.
In this study, a reproducible transformation system for Summer set tomato cultivar using Agrobacterium tumefaciens and cotyledonary explants was developed. The results presented are the average of three independent experiments with 100 explants in each trial. Both explant types have shown high capacities for regeneration. However, cotyledons were found to be superior compared to the hypocotyls. In addition, the type and the concentration of phytohormones in the induction media were found to be crucial factors controlling tomato regeneration.
Compared to the transformed tissues, high frequency of shoot induction was observed in the untransformed tissues. This may be attributed to the competence of the cells for regeneration (Velcheva et al., 2005). Direct shoot initiation, rather than the callus, was observed at the edge of the cotyledonary proximal end. Similar observations were reported by Raj et al. (2005) and Peres et al. (2001) who attributed this to be due to hormonal metabolism and/or genetic background of organogenetic competence. The possibility of non-transformed shoot survival on the elongation medium is generally reported in similar transformation trials giving a false positive indication (Velcheva et al., 2005). The regeneration of non-transformed plants was explained by non-efficient selection due to the protection of non-transformed cells presumbly caused by the occurrence of chimeric plants (Ghorbel et al., 1999; De Almeida et al., 2003; Velcheva et al., 2005) or alternatively that the antibiotic used for selection (hygromycin) was partially or completely phosphorylated by cells expressing hpt gene (Bashir et al., 2004; Velcheva et al., 2005).
Previous studies aiming at improving shoot regeneration of transformed tissue from many tomato cultivars indicated the necessity of using a tobacco, petunia or tomato feeder layer for pre-culture and explant-Agrobacterium co-cultivation (McCormick et al., 1986; Fillatti et al., 1987; Delannay et al., 1989; Van Roekel et al., 1993; Agharbaoui et al., 1995; Frary and Earle, 1996; Ling et al., 1998; Tabaeizadeh et al., 1999). Hamza and Chupeau, (1993) showed that pretreatment with feeder cells stimulated tomato cell transformation but significantly reduced regeneration of transformed cells. Although, in this study, no feeder layer was used for pre-culture or Agrobacterium co-cultivation, an adequate transformation frequency (7%) was obtained. (Park et al., 2003) reported similar results on other tomato cultivars with a transformation frequency of 20%.
CONCLUSION
When grown off-season under tropical conditions of the Sudan, tomato is likely to suffer from fungal infections. We have established a transformation/regeneration system for successful insertion of an antifungal protein (plant defensin) into tomato cv. Summer set using A. tumefaciencs as a vector. The result is considered to be a significant step in the development of commercial fungal resistant transgenic tomato cultivars.
REFERENCES
- Abbo, A.S.H., M.O. Idris and M.M.A. El-Balla, 2009. The response of tea tree oil as a biofungicide against early blight disease in tomato crop (solanum lycopersicum) in Sudan. Proceedings of the Conference on International Research on Food Security, Natural Resource Management and Rural Development, Tropentag 2009, Oct. 6-8, University of Hamburg, pp: 1-9.
Direct Link - Agharbaoui, Z., A.F. Greer and Z. Tabaeizadeh, 1995. Transformation of the wild tomato Lycopersicon chilense dun. by Agrobacterium tumefaciens. Plant.Cell. Rep., 15: 102-105.
CrossRefDirect Link - De Almeida, W.A.B., F. de Assis Alves Mourao Filho, B.M.J. Mendes, A. Pavan, A.P.M. Rodriguez, 2003. Agrobacterium-mediated transformation of Citrus sinensis and Citrus limonia epicotyl segments. Scientia Agricola, 60: 23-29.
CrossRefDirect Link - Bhatia, P., N. Ashwanth, T. Senaratna and D. Midmore, 2004. Tissue culture studies of tomato (Lycopersicon esculentum). Plant Cell. Tissue Organ Cult., 78: 1-21.
CrossRefDirect Link - Costa, M.G.C., F.T.S. Nogueira, M.L. Figueira, W.C. Otoni, S.H. Brommonschenkel and P.R. Cecon, 2000. Influence of the antibiotic timentin on plant regeneration of tomato (Lycopersicon esculentum Mill.) cultivars. Plant Cell Rep., 19: 327-332.
CrossRefDirect Link - Delannay, X., B.J. LaVallee, R.K. Proksch, R.L. Fuchs and S.R. Sims et al., 1989. Field performance of transgenic tomato plants expressing the Bacillus thuringiensis var. kurstaki insect control protein. Nat. Biotechnol., 7: 1265-1269.
Direct Link - Dillen, W., J. de Clercq, J. Kapila, M. Zambre, M. van Montagu and G. Angenent, 1997. The effect of temperature on Agrobacterium tumefaciens-mediated gene transfer to plants. Plant J., 12: 1459-1463.
CrossRefDirect Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Fillatti, J.J., J. Kiser, R. Rose and L. Comai, 1987. Efficient transfer of a glyphosate tolerance gene into tomato using a binary Agrobacterium tumefaciens vector. Nat. Biotechnol., 5: 726-730.
CrossRefDirect Link - Frary, A. and E.D. Earle, 1996. An examination of factors affecting the efficiency of Agrobacterium-mediated transformation of tomato. Plant Cell Rep., 16: 235-240.
CrossRef - Ghorbel, R., J. Juarez, L. Navarro and L. Pena, 1999. Green fluorescent protein as a screenable marker to increase the efficiency of generating woody fruit plants. TAG Theor. Applied Genet., 99: 350-358.
CrossRef - Hamza, S. and Y. Chupeau, 1993. Re-evaluation of conditions for plant regeneration and Agrobacterium-mediated transformation from tomato (Lycopersicon esculentum). J. Exp. Bot., 44: 1837-1845.
CrossRefDirect Link - Hansen, G., A. Das and M.D. Chilton, 1994. Constitutive expression of the virulence genes improves the efficiency of plant transformation by Agrobacterium. Proc. Natl. Acad. Sci., 91: 7603-7607.
PubMedDirect Link - Hu, W. and G.C. Phillips, 2001. A combination of overgrowth-control and antibiotics improves Agrobacterium tumefaciens-mediated transformation efficiency for cultivated tomato (L. esculentum). In Vitro Cell. Dev. Biol. Plant, 37: 12-18.
Direct Link - Humara, J.M., M. Lopez and R.J. Ordas, 1999. Agrobacterium tumefaciens-mediated transformation of Pinus pinea L. cotyledons: An assessment of factors influencing the efficiency of uidA gene transfer. Plant Cell. Rep., 19: 51-58.
CrossRef - Jefferson, R.A., 1987. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep., 5: 387-405.
CrossRefDirect Link - Krasnyanski, S.F., J. Sandhu, L.L. Domier, D.E. Buetow and S.S. Korban, 2001. Effect of an enhanced CaMV 35S promoter and a fruit-specific promoter on uidA gene expression in transgenic tomato plants. In Vitro Cell. Dev. Biol. Plant, 37: 427-433.
CrossRef - Ling, H.Q., D. Kriseleit and M.W. Ganal, 1998. Effect of ticarcillin/potassium clavulanate on callus growth and shoot regeneration in Agerobacterium-mediated transformation of tomato (Lycopersicon esculentum Mill.). Plant Cell Rep., 17: 843-847.
CrossRefDirect Link - McCormick, S., J. Niedermeyer, J. Fry, A. Barnason, R. Horsch and R. Fraley, 1986. Leaf disc transformation of cultivated tomato (L. esculentum) using Agrobacterium tumefaciens. Plant Cell Rep., 5: 81-84.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Murray, S.L., R. Janse van Vuuren and D.K. Berger, 1998. Tomato transformation is influenced by acetosyringone and Agrobacterium tumefaciens cell density. J. Southern Afr. Soc. Hortic. Sci., 8: 60-64.
Direct Link - Park, S.H., J.L. Morris, J.E. Park, K.D. Hirschi and R.H. Smith, 2003. Efficient and genotype-independent Agrobacterium-mediated tomato transformation. J. Plant Physiol., 160: 1253-1257.
CrossRefPubMedDirect Link - Peres, L.E.P., P.G. Morgante, C. Vecchi, J.E. Kraus and M.A. van Sluys, 2001. Shoot regeneration capacity from roots and transgenic hairy roots of tomato cultivars and wild related species. Plant Cell Tissue Organ Cult., 65: 37-44.
CrossRefDirect Link - Qiu, D., G. Diretto, R. Tavarza and G. Giuliano, 2007. Improved protocol for Agrobacterium mediated transformation of tomato and production of transgenic plants containing carotenoid biosynthetic gene CsZCD. Sci. Hortic., 112: 172-175.
CrossRef - Raj, S.K., R. Singh, S.K. Pandey and B.P. Singh, 2005. Agrobacterium-mediated tomato transformation and regeneration of transgenic lines expressing tomato leaf curl virus coat protein gene for resistance against TLCV infection. Curr. Sci., 88: 1674-1679.
Direct Link - Rossi, L., B. Hohn and B. Tinland, 1996. Integration of complete transferred DNA units is dependent on the activity of virulence E2 protein of Agrobacterium tumefaciens. Proc. Natl. Acad. Sci. USA., 93: 126-130.
Direct Link - Santa-Cruz, A., M.M. Martinez-Rodriguez, F. Perez-Alfocea, R. Romero-Aranda and M.C. Bolarin, 2002. The rootstock effect on the tomato salinity response depends on the shoot genotype. Plant Sci., 162: 825-831.
CrossRefDirect Link - Saker, M.M., H.A. Hussein, N.H. Osman and M.H. Soliman, 2007. In vitro production of transgenic tomatoes expressing defensin gene using newly developed regeneration and transformation system. Arab J. Biotech., 11: 59-70.
Direct Link - Stomp, A.M., C. Loopstra, W.S. Chilton, R.R. Sederoff and L.W. Moore, 1990. Extended host range of Agrobacterium tumefaciens in the genus Pinus. Plant Physiol., 92: 1226-1232.
Direct Link - Sun, H.J., S. Uchii, S. Watanabe and H. Ezura, 2006. A highly efficient transformation protocol for Micro-Tom, a model cultivar for tomato functional genomics. Plant Cell Physiol. 47: 426-431.
CrossRefPubMedDirect Link - Tabaeizadeh, Z., Z. Agharbaoui, H. Harrak and V. Poysa, 1999. Transgenic tomato plants expressing a Lycopersicon chilense chitinase gene demonstrate improved resistance to Verticillium dahliae race 2. Plant Cell Rep., 19: 197-202.
CrossRef - Tang, W., H. Luo and R.J. Newton, 2004. Effects of antibiotics on the elimination of Agrobacterium tumefaciens from loblolly pine (Pinus taeda) zygotic embryo explants and on transgenic plant regeneration. Plant Cell Tissue Organ Cult., 79: 71-81.
CrossRef - Van Roekel, J.S.C., B. Damm, L.S. Melchers and A. Hoekema, 1993. Factors influencing transformation frequency of tomato (Lycopersicon esculentum). Plant Cell Rep., 12: 644-647.
CrossRefDirect Link - Velcheva, M., Z. Faltin, M. Flaishman, Y. Eshdat and A. Perl, 2005. A liquid culture system for Agrobacterium-mediated transformation of tomato (Lycopersicon esculentum L. Mill.). Plant Sci., 168: 121-130.
CrossRefDirect Link - Vidya, C.S.S., M. Manoharan, C.T.R. Kumar, H.S. Savithri and G.L. Sita, 2000. Agrobacterium-mediated transformation of tomato (Lycopersicon esculentum var. pusa ruby) with coat-protein gene of Physalis mottle tymovirus. J. Plant Physiol., 156: 106-110.
Direct Link - Bashir, K., M. Rafiq, F. Tahira, T. Husnain and S. Riazuddin, 2004. Hygromycin based selection of transformants in a local inbred line of Zea mays (L.). Pak. J. Biol. Sci., 7: 318-323.
CrossRefDirect Link - Uddin, M.F., S.M.A. Islam, S. Naznin and M.H.K. Shiragi, 2004. Effect of variety and plant growth regulators in ms medium on callus proliferation from virus infected tomato plant. Biotechnology, 3: 181-186.
CrossRefDirect Link - Geneif, A.A., 1986. Attempts to control tomato leafcurl virus on tomato in the sudan. Acta Hort., 190: 145-158.
Direct Link - Jinnah, M.A., K.M. Khalequzzaman, M.S. Islam, M.A.K.S. Siddique and M. Ashrafuzzaman, 2002. Control of bacterial wilt of tomato by Pseudomonas fluorescens in the field. Pak. J. Biol. Sci., 5: 1167-1169.
CrossRefDirect Link - Mukhtar, S.K., A. Hashim, M. Peterschmitt and M.K. Abdrahman, 2009. Field screening and molecular identification of tomato leaf curl virus in Sudan. Arab J. Plant Protec., 27: 95-98.
Direct Link - Shahriari, F., H. Hashemi and B. Hosseini, 2006. Factors influencing regeneration and genetic transformation of three elite cultivars of tomato (Lycopersicon esculentum L.). Pak. J. Biol. Sci., 9: 2729-2733.
CrossRefDirect Link