
Virgilio Gavicho Uarrota
Plant Science Center, Plant Morphogenesis and Biochemistry Laboratory, Federal University of Santa Catarina 88040-900, P.O. BOX 476, SC, Florianopolis, Brazil
Ricardo Brasil Severino
Plant Science Center, Plant Morphogenesis and Biochemistry Laboratory, Federal University of Santa Catarina 88040-900, P.O. BOX 476, SC, Florianopolis, Brazil
Marcelo Maraschin
Plant Science Center, Plant Morphogenesis and Biochemistry Laboratory, Federal University of Santa Catarina 88040-900, P.O. BOX 476, SC, Florianopolis, Brazil
International Journal of Agricultural Research
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 218-226







ABSTRACT
Maize (Zea mays L.) is a staple food for millions of people in the world and its grain is converted into well-accepted local food products, including gruels, porridges, pastes and infant weaning food. Maize landraces synthesize a myriad of secondary metabolites and these compounds play important roles throughout the plant’s life cycle as mediators in the interaction of the plant with its environment such as plant-insect, plant-microorganism and plant-plant interaction. Secondary metabolites determine important aspects in human food quality. Moreover, several plant secondary metabolites are used for the production of medicines, dyes, insecticides, flavors and fragrances. The extensive genetic and chemical diversity of maize results from a long domestication process carried out since Pre-Colombian civilizations. More than 250 maize varieties are known worldwide and their origin is linked directly or indirectly to the maize domestication and breeding processes performed by ancient civilizations. One of the properties of maize is the spectacular diversity in morphology among its races, which is seemingly paralleled by an extensive allelic variation as detected by molecular methods. On the other hand, despite the fact that modern farming techniques have greatly increased the yield of maize, the genetic breeding programs adopted have led to a greater genetic uniformity and a meaningful loss of diversity. In fact, nowadays very few of the world’s maize germplasm consist of local and creole varieties (landraces), showing the genetic vulnerability of that species. In this context, small farmers in some regions of the world still cultivate maize landraces that are populations with high genetic variability and represent a valuable source of potentially useful traits such as resistance or tolerance to biotic and abiotic stress factors. In this review, a few of the major issues that should be taken into consideration in approaching maize landraces as source of secondary metabolites of interest for human health and nutrition will be briefly discussed.
PDF Abstract XML References Citation
Received: December 20, 2010;
Accepted: January 17, 2011;
Published: March 28, 2011
How to cite this article
Virgilio Gavicho Uarrota, Ricardo Brasil Severino and Marcelo Maraschin, 2011. Maize Landraces (Zea mays L.): A New Prospective Source for Secondary Metabolite Production. International Journal of Agricultural Research, 6: 218-226.
DOI: 10.3923/ijar.2011.218.226
URL: https://scialert.net/abstract/?doi=ijar.2011.218.226
DOI: 10.3923/ijar.2011.218.226
URL: https://scialert.net/abstract/?doi=ijar.2011.218.226
INTRODUCTION
Maize (Zea mays L.) is a widely consumed cereal worldwide and contains carotenoids, phenolics and anthocyanins which are phytochemicals synthesized in the plant by secondary metabolite pathways (Lopez-Martinez et al., 2009). These compounds form a large reservoir of natural chemical diversity as they have a variety of unique carbon skeletons and functional group modifications (Kutchan and Dixon, 2005). By definition, secondary metabolites are not essential for the growth and development of a plant, but rather are required for the interaction of a plant with its environment. The multiple levels at which a plant can communicate with other organisms, either in competition or synergy, are reflected in the functional variety found in secondary metabolites. The obvious demands of a dynamic environment confer a natural plasticity to secondary metabolism and drive the evolution of genetic diversification, which has resulted in an arsenal of natural products (Kutchan and Dixon, 2005). Because much of secondary metabolism is genus or even species-specific, as well as many of the confirmed functions of plant secondary metabolites are quite varied or, at worse, unknown, it is well recognized that this field has not been fully explored in its potential for medicinal purposes, for instance (Maraschin and Verpoorte, 1999). This scenario has been gradually changed with the increase availability of powerful molecular tools for profiling transcripts and metabolites on an unprecedented scale such as Nuclear Magnetic Resonance (NMR), Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight Mass Spectrometry (MALDI-TOF MS), Gas Chromatography (GC) and High Performance Liquid Chromatography (HPLC)-(Maraschin et al., 2001), as well as with the realization that secondary metabolism, although complex, has evolved from primary metabolism in a way that can be understood and exploited for the creation of novel bioactive molecules. Once an area of research reserved for organic chemists, the study of plant secondary metabolism has expanded from the isolation of natural products and the elucidation of their structures to an analysis of enzymes, genes, regulation and genetic engineering.
Technological advances in analytical chemistry, in particular in the development of high-field nuclear magnetic resonance spectroscopy, mass spectrometry (electrospray and MALDI-MS, e.g.,) and Fourier transform-ion cyclotron mass spectrometry, have facilitated the identification and elucidation of the structures of new compounds present even at low levels within a plant. An enormous quantity of information has been gained on chemical structure of plant secondary metabolites in the past decades, but how can one exploit more this chemo (bio) diversity? This study illustrates modern insights and how useful maize landraces might be as prospected as sources of bioactive compounds.
Mankind has been using plant secondary metabolites for the past few thousand years for multiple purposes such as dyes, flavors, stimulants, hallucinogens, vertebrate and human poisons, as well as therapeutic agents. The interest in secondary metabolites has increased in recent years since many investigations with respect to human nutrition pointed out that modest and long-term intake of certain secondary metabolites will have a major impact on preventing incidences of cancers and many chronic diseases (Kutchan and Dixon, 2005). In this context, maize landraces are thought to be potential sources of bioactive compounds and could be more intensively exploited since it is known that those genotypes are rich in chemical diversity (Kuhnen et al., 2010).
PHARMACOLOGICAL ROLES OF SECONDARY METABOLITES EXTRACTED FROM MAIZE LANDRACES
Maize landraces contain, among others, carotenoids, anthocyanins and phenolic compounds. These secondary metabolites play important roles in human health. Epidemiological and in vitro researches suggest an inverse relationship between consumption of fruits and vegetables and the incidence of various chronic and degenerative diseases that come with aging such as cancer, cardiovascular diseases, cataracts and brain and immune dysfunction (Lopez-Martinez et al., 2009). The health beneficial properties of plant metabolites, carotenoids and (poly) phenols, for instance, have been related not only to their high antioxidant and anti-radical activities, but also to several other biological properties, e.g., anti-mutagenic, pro-estrogenic, anti-angiogenic and induction of detoxification enzymes such as glutathione transferase and quinine reductase (Lopez-Martinez et al., 2009; Kuhnen et al., 2009).
The xanthophylls (oxycarotenoids) lutein and zeaxanthin (Fig. 1) the main carotenoids found in maize grains and derived products, carry desirable health-related properties, e.g., enhancement of immune function, blockage of mammary tumor growth and protection against blindness caused by age-related macular degeneration. These and other human diseases such as psoriasis, diabetes-related blindness and tumor growth were already referred to angiogenic diseases characterized by the uncontrolled formation of blood vessels (i.e., neovascularization). Studies have demonstrated that the xanthophyll-rich seed extract of maize displays anti-vasculo/angiogenic properties, suggesting a potential role of compounds such as zeaxanthin and lutein in the prevention of diseases related to uncontrolled process of vessel formation. The results found shed light on the importance of chemical prospective approaches as an add-value strategy to raw materials of maize genotypes, since some of their secondary metabolites (e.g., lutein and zeaxanthin) are quite valuable compounds. In fact, some ongoing researchers in maize landraces have been confirming these potentialities (Kuhnen et al., 2009, 2010).
NEW INSIGHTS IN MAIZE LANDRACES AND EMERGING OPPORTUNITIES
In recent years, the use of plants as bioreactors has emerged as an exciting area of research and significant advances have created new opportunities. The driving forces behind the rapid growth of plant bioreactors include low production cost, product safety and easy scale up. As the yield and concentration of a compound is crucial for commercial viability, several strategies have been developed to boost up the biosynthesis of some target secondary metabolites (Maraschin et al., 2002; 2000a).
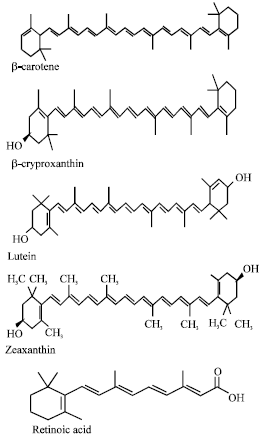 | |
| Fig. 1: | Chemical structures of carotenoids found in maize landraces seeds, silks and flowers. Retinoic acid is an oxidized and more stable form of retinol, i.e., vitamin A |
Maize landraces usually present a well adapted phenotype in their sites of cultivation resulting from a peculiar genetic variability positively interacting with (a) biotic factors, e.g., soil, climate, pests and diseases. For that, an expressive number of secondary metabolites are thought to be involved regulating the ecological relationships of a given maize landrace population and its environment. This way, a better understanding of the effects of ecological factors on the adaptation response of those genotypes seems to be relevant as one aims at to improve the yield of a target bioactive compound in populations of Z. mays landraces. Additionally, such an approach has increased in importance since maize landraces are claimed to be low-energy input crops as they allow obtaining a certain yield with minimum ecological impact (i.e., eco-friendly). On the other hand, despite the great potential as source of already known or new active biomolecules, studies on the chemical diversity of maize landraces are lacking.
In a second approach, Ethnopharmacological studies from various regions of the world on the use of maize landraces have been published showing pharmacological activities that include among others mild diuretic, tonic and urinary demulcent, antispasmodic, anti inflammatory and antioxidant properties. Aqueous extracts of maize female flowers, i.e., stigma and styles, have also been used to pass kidney stones, to cure bladder ailments, gout, benign prostatic hyperplasia, edema (water retention), lowering blood pressure and even helping rheumatism symptoms or hypertensions (Kuhnen et al., 2010).
Discovery of a single new pharmaceutical agent originated from plant material might be a time-cost and extremely expensive process. In a random collection for pharmacological screening, most part of the plant extracts tested (95% or more) is inactive and great part of those active extracts contain already known compounds. If Ethnopharmacological or Ethnobotanical research is used to provide initial information, success in screening is significantly improved; with 20-60% of plant extracts tested showing some pharmacological activity (Soejarto et al., 2005). One valuable strategy used to improve the identification of bioactive compounds from plants was known as ‘ethno-directed’ screening and encompasses field working with traditional healers (“curanderos”). Taking into account that maize landraces are well known genotypes in their native regions, one can find information on the traditional knowledge associated to their usage for both nutritional and human health treatments. However, despite the considerable research activity in natural products identification and activity analysis, this field has not been enough explored and this is also true for maize landraces.
ECOLOGICAL FACTORS OF INFLUENCE ON THE CONTENT OF SECONDARY METABOLITES
Since secondary metabolites represent a chemical interface between plants and surrounding environment, their syntheses are frequently affected by environmental conditions. Thus, variations in the total content and/or of the relative proportions of secondary metabolites in plants can take place. The main environmental factors that can streamline or alter the production or concentration of secondary metabolites in plants are seasonality, circadian rhythm, developmental stage and age, temperature, water stress, UV radiation, soil nutrients, altitude, atmospheric composition, tissue damage (Gobbo-Neto and Lopes, 2007) and phosphorus nutrition (Uarrota, 2010). Figure 3 shows some examples of secondary metabolites found in maize landraces tissues (Kuhnen et al., 2010) that are affected by the factors above.
In recent years, efforts are underway to improve the carotenoid levels in staple food crops to overcome vitamin A deficiency in areas with limited access to animal products, fruits and vegetables. An increasing number of studies have been performed showing the bioactive potential of secondary metabolites (Maraschin et al., 2000b, 2003). Maize landraces are sources of three carotenoids, namely β-carotene, β-cryptoxanthin and α-carotene that are pro-vitamin A compounds. Besides, maize is also a good source of nonprovitamin A carotenoids, including lutein and zeaxanthin, which play beneficial roles in human health (Menkir et al., 2008). The consumption of carotenoid-rich foods is associated with reduced risks of developing cardiovascular diseases, enhanced immune responses, improved vision and prevention of night blindness, as well as the maintenance of healthy skin and gastrointestinal and respiratory systems. Increased dietary intake of lutein and zeaxanthin has been associated with lowering of the risk of cataracts, age-related macular degeneration and other degenerative diseases (Menkir et al., 2008). Since the various carotenoids have distinct roles in human metabolism, further enhancement of their levels in staple food crops may have a positive health impact in areas where maize is consumed. Of interest, maize is a staple food crop of utmost importance in less favored countries where, no rare, chronic vitamin A deficiency in childhood is a major concern. The stimulus to low-energy input crops cultivation (maize landrace, for instance) in such countries is claimed to be relevant for obvious economical, social and ecological reasons.
MAJOR IMPACT OF CARBON, NITROGEN AND SULFUR RESOURCES ON THE PRODUCTION OF SECONDARY METABOLITES
Nutrient supply plays a major role regulating biosynthesis, accumulation and degradation pathways in plants. For example, the content of the metabolically linked carbon and nitrogen compounds has a major quantitative impact on the extent of production of secondary metabolites in plants. The carbon-nitrogen balance hypothesis claims that under conditions of limited nitrogen availability as usually found in production systems of maize landraces, secondary metabolism is directed toward producing carbon-rich metabolites and vice versa (Coley et al., 1985). Yet, this idea seems to be correct for some cases but not all. This is most probably due to the fact that changes in carbon and nitrogen availability (such as photosynthesis and nitrogen fertilization) do not always lead to analogous changes in the relative levels of carbon-rich and nitrogen-rich primary metabolites, but rather the opposite (Matt et al., 2002). On the other hand, the relative endogenous pools of carbon-rich or nitrogen-rich primary metabolites seem to have a strong correlation with the levels of carbon-rich or nitrogen-rich secondary metabolites (Fritz et al., 2006). Matt et al. (2002) showed that an increase in the sugar/amino acid ratio resulted in the elevation of carbon-rich phenylpropanoids and a decline in the levels of the nitrogen-containing alkaloid nicotine. Sulfur (S) is also an essential primary compound for plants, taking part in enzyme functions (e.g., formation of disulfide bonds and S-containing cofactors), redox regulation (e.g., glutathione and Fe-S clusters), donation of methyl groups in numerous reactions, including DNA methylation and mRNA capping through S-adenosylmethionine (i.e. SAM), as well as in the synthesis of polyamines (Rausch and Wachter, 2005). However, S is also incorporated into an array of secondary metabolites (Hirai and Saito, 2008). Catabolic recycling of S-containing secondary metabolites occurring upon plant growth in S-deficient environments is crucial for the response of the plants to various stresses. In cruciferous plants, for example, including the model plant Arabidopsis, glucosinolates are the major S-containing secondary metabolites that are required for plant defense against pathogens (Sonderby et al., 2010). Indeed, the activation of sulfate acquisition and repression of glucosinolates production occur in parallel under S limitation. Under S-limiting conditions, plants generally enhance S assimilation through activating S transporters, while at the same time also activating a putative thioglucosidase, thought to be involved in the hydrolytic degradation of glucosinolates for catabolic sulfur recycling, as well as in negatively regulating glucosinolates biosynthesis genes (Aharoni and Galili, 2010).
Expanding this subject to maize landraces, it is worth mentioning that those genotypes usually show a good adaptation to poor soils, even guaranteeing interesting grain yields under limiting soil conditions for hybrid varieties. Such a phenotype is thought to be in connection with the synthesis and accumulation of secondary metabolites, i.e., an ecological strategy of survival of Z. mays landraces populations, with eventual meaningful production of valuable compounds for human health. Unfortunately, such an issue seem not to be enough understood, avoiding a more intensive and rational exploitation of the maize landrace’s potential.
METABOLIC PATHWAYS OF SECONDARY METABOLITES
Although in the past 10 years quite some research has been done aiming at to increase secondary metabolites production, a major constraint has been the poor characterization of plant secondary metabolic pathways at the level of biosynthetic intermediates and enzymes. Consequently, very few genes from plant secondary metabolism are available, hampering the improvement of yield of a given compound of interest through genetic manipulation, for example. The best studied pathway at the genetic level seems to be the one leading to the formation of flavonoids and anthocyanins. Because of their antioxidant activity, high levels of anthocyanins in foods, for example, are an interesting trait and this is especially relevant for some maize landraces where prominent amounts of anthocyanins and their aglycone forms (e.g., cyanidin, delphinidin and malvidin) are found in their grains and female flowers (Kuhnen et al., 2010). Figure 2 shows an overview of the biosynthetic pathways of plant secondary metabolites and their main intermediary compounds.
More often it has been aimed to increase the production of certain compounds in the normal producing plant species by epigenetic manipulation (Maraschin and Verpoorte, 1999). However, when the results are unsatisfactory, techniques of genetic manipulation have been used to transfer (part of) a pathway to other plant species (i.e., heterologous transformation) or microorganisms (Verpoorte and Memelink, 2002). For that, from a metabolic engineering point of view, two general approaches have been followed to increase the production of target compounds. Firstly, methods have been employed to change the expression of one or few genes, thereby overcoming specific rate limiting steps in the pathway, to shutdown competitive pathways and to decrease catabolism of the product of interest. Secondly, attempts have been made to change the expression of regulatory genes that control multiple biosynthesis genes (Verpoorte and Memelink, 2002). The tight link between metabolic fluxes of primary metabolites and the accumulation of secondary metabolites renders the engineering of the latter compounds quite complex as it demands the consideration of the entire metabolic network in order to redirect primary metabolites into secondary metabolites with respect to the primary-secondary interface (Aharoni and Galili, 2010). Indeed, it seems to be consensus that for a successful genetic manipulation of secondary metabolites biosynthetic pathways of plant species, deeper studies are urgent regarding the dynamic interactions between primary and secondary metabolic pathways and the effect of intrinsic and ecological regulator factors, as one envisage technological applications thereof (Maraschin and Verpoorte, 1999). On this subject, maize landraces, thanks to their high genetic variability, have been claimed as important sources of genes for genetic breeding programs aiming at to improve nutritional and health traits of Z. mays grains and derived products.
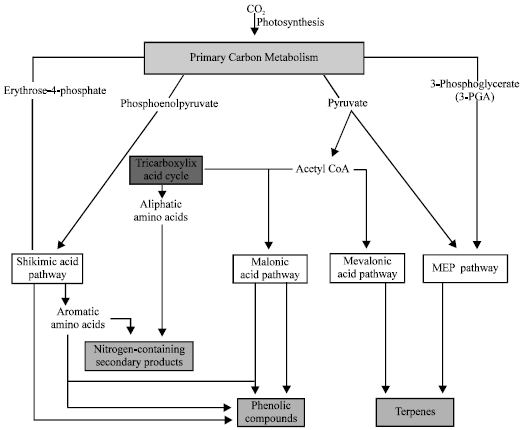 | |
| Fig. 2: | The production of secondary metabolites is tightly associated with pathways of primary/central metabolism, such as glycolysis, shikimate and production of AAA and aliphatic amino acids |
 | |
| Fig. 3: | Secondary metabolites whose occurrence is affected by seasonality, circadian rhythm, water availability and tissue damage in maize landraces |
CONCLUSIONS
In past few years, an increase number of studies have been revealing maize landraces as useful sources of raw materials for secondary metabolite production such as carotenoids, anthocyanins and phenolics compounds which are thought to play important roles in human health. Thus, maize landraces constitute a prospective germplasm for improving or increasing the production of those compounds with direct application in food, pharmaceutical and cosmetical industries. In fact, there seem to be a wide range of secondary metabolite profiles among individuals and varieties analyzed under the same growing condition, allowing the exploitation of a prominent chemical diversity of those genotypes. Besides, within this framework of ideas, heirloom maize landraces varieties are a still important resource against genetic erosion and as livelihood for uncountable small farmers in several countries. Otherwise, in comparison with the genetic diversity of maize landraces available world wide, studies focusing on the chemical characterization of those genotypes are still scarce in the literature. In fact, more studies are needed to understand the regulation and integration of secondary metabolism pathways with primary ones as well as the (a) biotic factors affecting the synthesis of health-valuable compounds and how to optimize it in maize landraces. Finally, the extinguishment of such germplasm resulting from the adoption of modern agricultural techniques, i.e., hybrid and transgenic maize varieties, is a continuous concern and its potential as source of valuable bioactive molecules might shed new insights into its usage, corroborating to preserve that invaluable genetic resource.
ACKNOWLEDGMENTS
This study was carried out under CNPq-Brazil/MCT-Mozambique Postgraduate fellowship program. The authors wish to thank all anonymous reviewers.
REFERENCES
- Aharoni, A. and G. Galili, 2010. Metabolic engineering of the plant primary-secondary metabolism interface. Curr. Opin. Biotechnol., Vol. 22.
CrossRef - Coley, P.D., J.P. Bryant and F.S. Chapin, 1985. Resource availability and plant antiherbivore defense. Science, 230: 895-899.
PubMed - Gobbo-Neto, L. and N.P. Lopes, 2007. Medicinal plants: Factors of influence on the content of secondary metabolites. Quím. Nova, 30: 374-381.
CrossRefDirect Link - Matt, P., A. Krapp, V. Haake, H. Mock and M. Stitt, 2002. Decreased Rubisco activity leads to dramatic changes of nitrate metabolism, amino acid metabolism and the levels of phenylpropanoids and nicotine in tobacco antisense RBCS transformants. Plant J., 30: 663-677.
PubMed - Fritz, C., N. Palacios-Rojas, R. Feil and M. Stitt, 2006. Regulation of secondary metabolism by the carbon-nitrogen status in tobacco: Nitrate inhibits large sectors of phenylpropanoid metabolism. Plant J., 46: 533-548.
PubMed - Rausch, T. and A. Wachter, 2005. Sulfur metabolism: A versatile platform for launching defence operations. Trends Plant Sci., 10: 503-509.
CrossRef - Hirai, M.Y. and K. Saito, 2008. Analysis of systemic sulfur metabolism in plants using integrated omics strategies. Mol. BioSyst., 4: 967-973.
CrossRef - Sonderby, I.E., F. Geu-Flores and B.A. Halkier, 2010. Biosynthesis of glucosinolates-gene discovery and beyond. Trends Plant Sci., 15: 283-290.
PubMed - Verpoorte, R. and J. Memelink, 2002. Engineering secondary metabolite production in plants. Curr. Opin. Biotechnol., 13: 181-187.
CrossRefDirect Link - Menkir, A., W. Liu, W.S. White, B. Maziya-Dixon and T. Rocheford, 2008. Carotenoid diversity in tropical-adapted yellow maize inbred lines. Food Chem., 109: 521-529.
CrossRef - Kutchan, T. and R.A. Dixon, 2005. Physiology and metabolism: Secondary metabolism: Natures chemical reservoir under deconvolution. Curr. Opin. Biotechnol., 8: 227-229.
CrossRef - Kuhnen, S., J.B. Ogliari, P.F. Dias, M. da Silva Santos and A.G. Ferreira et al., 2010. Metabolic fingerprint of Brazilian maize landraces silks (stigmata/styles) using NMR spectroscopy and chemometric methods. J. Agric. Food Chem., 58: 2194-2200.
PubMed - Kuhnen, S., P.M.M. Lemos, L.H. Campestrini, J.B. Ogliari, P.F. Dias and M. Maraschin, 2009. Antiangiogenic properties of carotenoids: A potential role of maize as functional food. J. Functional Foods, 1: 284-290.
CrossRef - Lopez-Martinez, L.X., R.M. Oliart-Ros, G. Valerio-Alfaro, C.H. Lee, K.L. Parkin and H.S. Garcia, 2009. Antioxidant activity, phenolic compounds and anthocyanins content of eighteen strains of Mexican maize. LWT Food Sci. Technol., 42: 1187-1192.
CrossRef - Maraschin, M., S.G. Carobrez, D. Persike, M.L. Peixoto and A.G. Ferreira et al., 2000. Cell wall polysaccharides from Mandevilla velutina (Apocynaceae) cultured cells: Extraction and chemical structure. Carbohydrate Polymers, 41: 55-60.
CrossRef - Maraschin, M., J.A. Sugul, K.V. Wood, C. Bonham and D.F. Buchi et al., 2002. Somaclonal variation: A morphogenetic and biochemical analysis of Mandevilla velutina cultured cells. Braz. J. Med. Biochem. Res., 35: 633-643.
Direct Link - Maraschin, M., C. Goncalves, R. Passos, P.F. Dias, R.M.R. Valle, J.D. Fontana and M.L. Pessatti, 2000. Isolation, chemical characterization and biological activities of cell wasll polysaccharides of Laurencia microcladia (Rhodomelaceae, Ceramiales). Tech. News Facimar, 4: 37-41.
Direct Link - Maraschin, M., J.A. Sugul, K.V. Wood, C. Bonham and F.M. Lancas et al., 2001. Supercritical CO2 extraction of velutinol a from Mandevilla velutina (Apocynaceae) cultured cells and MALDI-TOF MS analysis. Biotechnol. Lett., 23: 77-82.
CrossRef - Uarrota, V.G., 2010. Response of cowpea (Vigna unguiculata L. Walp.) to water stress and phosphorus fertilization. J. Agron., 9: 87-91.
CrossRefDirect Link - Soejarto, D.D., H.H. Fong, G.T. Tan, H.J. Zhang and C.Y. Ma et al., 2005. Ethnobotany/ethnopharmacology and mass bioprospecting: Issues on intellectual property and benefit-sharing. J. Ethnopharmacol., 100: 15-22.
CrossRefPubMedDirect Link