
Ravindra Kumar
Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005 (UP), India
Asha Sinha
Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005 (UP), India
Seweta Srivastava
Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005 (UP), India
Manisha Srivastava
Department of Botany, Harish Chandra (PG) College, Varanasi-221001 (UP), India
International Journal of Agricultural Research
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 172-179







ABSTRACT
The aim of the present study was to determine the decomposition rate of Sesbania aculeata L. by CO2 evolution and to estimate Soluble Crude Protein (SCP) production by dominant decomposing mycoflora of Sesbania aculeata L. Eight dominant decomposing mycobiota were selected for the study. In the substrate induced respiration the significant difference was observed in both sterilized and unsterilized substrate with the test fungi. The maximum CO2 evolution was observed with Aspergillus niger in sterilized (16.04 μg day-1) and unsterilized green manure (18.92 μg day-1). In other experiment conducted for the estimation of soluble crude protein production Penicillium citrinum has produced maximum SCP (26.54%) at 25°C followed by Trichoderma harzianum, Aspergillus niger and Curvularia lunata whereas minimum soluble crude protein production was observed in Penicillium rubrum (8.46%) at 35°C. The maximum per cent biomass reduction observed by Aspergillus niger (28.60%) at 25°C and minimum was found in Penicillium rubrum (3.80%) at 35°C. Among seven different nitrogen sources tested against Penicillium citrinum, the highest producer of SCP, potassium nitrate was found to be the best for maximum SCP production (26.54%) whereas the least suitable nitrogen source for SCP production by Penicillium citrinum was recorded to be sodium nitrate (14.85%).
PDF Abstract XML References Citation
How to cite this article
Ravindra Kumar, Asha Sinha, Seweta Srivastava and Manisha Srivastava, 2011. Substrate Induced Respiration and Soluble Crude Protein Production by Soil Mycobiota Involved in the Decomposition of Sesbania aculeata L. International Journal of Agricultural Research, 6: 172-179.
DOI: 10.3923/ijar.2011.172.179
URL: https://scialert.net/abstract/?doi=ijar.2011.172.179
DOI: 10.3923/ijar.2011.172.179
URL: https://scialert.net/abstract/?doi=ijar.2011.172.179
INTRODUCTION
Green manure crops are of vital concern particularly in the context of sustainable agriculture (Nayyar and Chhibba, 2000). Use of Sesbania as a green manure crop is a common practice in South Asia. Like most of the green manure crops, Sesbania belongs to the family Leguminosae and its subfamily is Papilionoideae. Species of the genus Sesbania are known for exceptionally fast growth rates as well as a very high affinity for association with several nitrogen-fixing Rhizobia in the soil that cause formation of numerous and large nodules in the plant roots (Hafeez et al., 2007).
The rates of CO2 evolution from soil are generally considered as a measure of metabolic activity of the soil biota. This soil respiration generally means the CO2 produced by soil fauna, microbes (Soil inhabiting fungi, bacteria and actinomycetes) and flora including root respiration. The relationship of rate of decomposition and CO2 evolution is well established phenomenon (Witkamp, 1966a, 1969; Okeke and Omaliko, 1992). When fresh organic matter is added to the soil it invariably undergoes decomposition to yield carbon dioxide as its major product. The rate of degradation of organic material depends largely on its chemical composition. Usually organic matter with higher nitrogen content and narrow C/N ratio undergoes faster decomposition than the one with poor nitrogen content. The addition of excess nitrogen to the organic residues with wider C/N ratio increased their rate of decomposition and CO2 evolution (Pernas, 1975). Bajpai et al. (1980) studied decomposition of paddy straw, Dhaincha and their mixture (1:1) in vitro under low temperature conditions of winter in normal and saline sodic soil. A burst in CO2 evolution occurred after 3 days of incubation with all plant materials in both soil types. Decomposition in terms of CO2 evolution and nutrient release was highest in normal soil. Maximum CO2 evolution occurred after 90 days in mixture, Dhaincha+paddy straw (1:1) decomposed at slowest rate in their study. Zaccheo et al. (2002) during their studies on decomposition of organic residues in soil used DRIFT (diffuse reflectance infrared Fourier-transform) spectroscopy to follow the early transformations that take place after the incorporation of organic materials in soil. Lucerne (A), Dried Maize (DM), laboratory-composted maize (CM) and two commercial composts (YWC and MWC) confined into glassfibre bags were incubated by them in sand with and without planting with lettuce in their study. DRIFT spectra of these materials before and after incubations were correlated with CO2-C evolution and mass, carbon and nitrogen balances. Joseph et al. (2006) conducted a field experiment to study the decomposition rate of green manure (Sesbania aculeata) by various stages and methods of its incorporation under intercropped wet-sown rice. Bedi et al. (2009) have studied on CO2 evolution in soils from green manures (including Sesbania aculeata L.), compost and nitrogen fertilizer under aerobic and anaerobic conditions. They reported higher cumulative CO2 evolution was shown by green manures dhaincha+50% N+ compost and lowest CO2 evolution was found in case of recommended N, under both the conditions.
Kahlon et al. (1980) reported the Soluble Crude Protein (SCP) production by different cellulolytic fungi viz., Chaetomium cellulolyticum, Rhizoctonia betaticola, Chaetomium globosum and Myrothecium verrucaria while using agricultural waste (wheat straw) as substrate. It was recorded that Chaetomium cellulolyticum gave highest percentage of protein (19.30%) whereas the highest carboxymethyl cellulose (CMC) activity was observed in case of Rhizoctonia betaticola. Sekhon (1975) reported that Chaetomium thermophile was found to produce maximum SCP when grown on delignified wheat straw. Bioconversion of delignified rice straw by eight different fungal species viz., Myrothecium verrucaria, Curvularia lunata, Sclerotium rolfsii, Fusarium equiseti, Helminthosporium maydis, Helminthosporium tetramera, Chaetomium globosum and Chaetomium cellulolyticum was studied by Dhillon et al. (1980). During their study highest SCP was observed by Chaetomium cellulolyticum (32.1), Hobbie et al. (2003) also studied SCP production using delignified agricultural by-products as substrate. Vibha and Sinha (2005) evaluated six cellulolytic fungi viz., Trichoderma harzianum, Penicillium citrinum, Curvularia lunata, Aspergillus flavus, Alternaria alternata and Aspergillus niger on rice stubble for SCP production. Trichoderma harzianum gave highest SCP production which was significantly superior to other fungi.
MATERIALS AND METHODS
This study has been conducted from 2008 to 2010.
For the study the decomposition of Sesbania aculeata L. in experimental conditions by soil mycobiota, the material was collected after harvesting of the Sesbania crop from the experimental site (Agricultural farm, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005). The whole study was categorized under two steps:
Study about the role of dominant decomposing mycoflora
Determination of decomposition rate of green manure by some dominant mycoflora by CO2 evolution: The rate of decomposition of the green manure by selected dominant fungi in terms of CO2 evolution was determined by the method described by Witkamp (1966b). The green manure was obtained from the experimental field and collected in sterilized polythene bags and brought into laboratory for the experiments. Green manure was cut in to small pieces (2 cm) and mixed properly. Six grams of green manure pieces were transferred into a 500 mL Erlenmeyer flasks containing 100 g moist sand (25-30% moisture) and were mixed together to sterilized at 121°C. Another batch of flasks containing moist sand only was sterilized; in these flasks 6 g unsterilized green manure pieces were mixed. The flasks inoculated individually with 1 mL standardized spore suspension (1.25x104 spores/cell mL-1) of the selected test fungi in triplicate, control were prepared by inoculating the flasks with sterilized distilled water. The flasks were incubated at room temperature (28±2°C) for four weeks and thereafter CO2 evolution was measured by the following method.
Cotton wool plugs were removed from mouth of the flasks and a small test tube (10x1.1 cm) containing 5 mL of 0.5 N KOH solution was inserted into a flask under aseptic conditions. The mouth of the flasks was tightly closed by rubber cork and CO2 absorbed at every 48 h interval was titrated with 0.1 N HCl using phenolphthalein as an indicator. The estimation of CO2 evolution, from both sterilized and unsterilized green manure substrates, was done continuously for 10 days at 2 days (48 h) interval. After every estimation fresh 0.5 N 5 mL KOH was inserted to the flasks. The results were expressed as μg CO2 evolution per day.
Estimation of Soluble Crude Protein (SCP) production by dominant mycoflora: The soluble crude protein production by dominant decomposing mycoflora was estimated by the procedure described by Dhillon et al. (1980). Air dried green manure (100 g), was cut in 2-3 cm in length and autoclaved with 1.8 L of 1% Sodium hydroxide (NaOH) at 121°C between 15-20 lbs for 1 h. After squeezing through nylon cloth the green manure was thoroughly washed with distilled water till neutral and dried at 60°C . The pre treated and dry green manure ground to 60 mesh and was used as sole source of carbon.
Basal synthetic medium described by Chahal and Grey (1969) was used and selected dominant fungi were grown in 50 mL of the basal synthetic medium and 500 mg delignified green manure. Four 250 mL Erlenmeyer flasks of each culture were incubated on rotary shaker at five different temperatures viz., 15, 20, 25, 30 and 35°C to test the effect of different temperatures on soluble crude protein production by dominant decomposing fungi. Five days after incubation the content of the flasks was filtered through tarred Whatmann filter No. 1 to determine the weight of the fungal mycelium and undigested cellulosic material. The dried biomass was analyzed for its nitrogen content using Kel plus auto-analyser. The nitrogen content was multiplied by 6.25 factor for the calculation of soluble crude protein. Seven different sources viz., Ammonium nitrate, ammonium sulphate, ammonium chloride, ammonium dihydrogen phosphate, potassium nitrate, sodium nitrate and urea were tested against highest SCP producing test fungus to find out the most suitable nitrogen source for soluble crude protein production. Each of the nitrogen sources was added in equivalent to 400 mg of nitrogen/L of medium used during the study.
Statistical analysis: The data obtained from CO2 evolution was analyzed using CRD design and the data for the effect of different temperatures on soluble crude protein production and per cent biomass reduction during soluble crude protein production was analysed using factorial RBD (Two factors fungixtemperature) and the results were expressed in terms of LSD (least significant difference).
RESULTS
Rate of Green manure decomposition by some dominant mycoflora in terms of CO2 evolution: Table 1 and 2 showed that the CO2 evolution was high in unsterilized green manure substrate as compared to sterilized substrate. The significant difference was observed in both sterilized and unsterilized substrate with the test fungi. The maximum CO2 evolution was observed with Aspergillus niger in sterilized (16.04 μg day-1) and unsterilized green manure (18.92 μg day-1).
| Table 1: | Estimation of rate of decomposition of Sesbania aculeata L. sterilized green manure by dominant decomposing fungi in terms of CO2 evolution (μg day-1) |
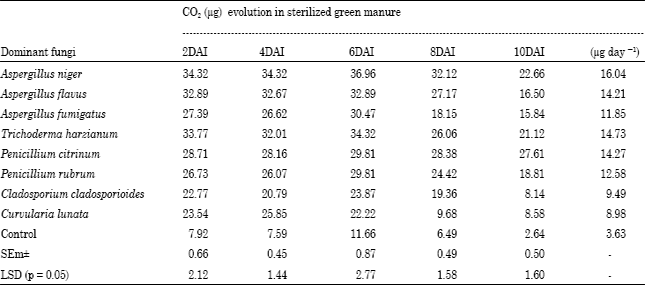 | |
DAI: Days After Insertion of KOH | |
| Table 2: | Estimation of rate of decomposition of Sesbania aculeata L. un-sterilized green manure by dominant decomposing fungi in terms of CO2 evolution (μg day-1) |
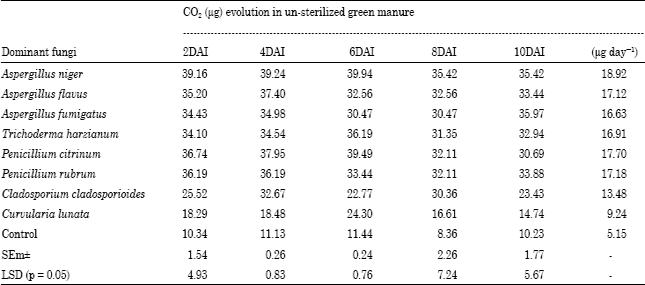 | |
DAI: Days After Insertion of KOH | |
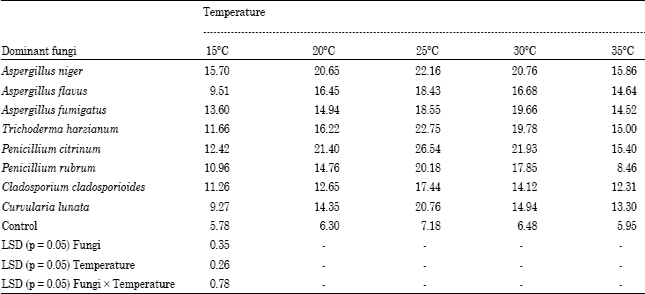
Soluble crude protein production by dominant decomposing fungi: The data on soluble crude protein production by eight selected dominant decomposers is presented in Table 3. Eight cellulolytic fungi were tested on delignified cellulose of green manure as a carbon source. The interaction between test fungi (dominant decomposers) and temperature was found significant. The maximum SCP production was recorded at 25°C by the dominant fungi. Maximum soluble crude protein was produced by Penicillium citrinum (26.54%) at 25°C and minimum was found in Penicillium rubrum (8.46%) at 35°C. The SCP production capacity of cellulolytic fungi was found in order of Penicillium citrinum (26.54%) >Trichoderma harzianum (22.75%) > Aspergillus niger (22.16%) > Curvularia lunata (20.76%) > Penicillium rubrum (20.18%)>Aspergillus fumigatus (18.55%)> Aspergillus flavus (18.43%)> Cladosporium cladosporioides (17.44%) at 25°C.
The data presented in Table 4 shows the maximum per cent biomass reduction observed by Aspergillus niger (28.60%) at 25°C and minimum was found in Penicillium rubrum (3.80%) at 35°C. The order of the per cent biomass reduction by dominant decomposing fungi was recorded at 25°C as Aspergillus niger (28.60%)>Penicillium citrinum (27.80%)>Trichoderma harzianum (22.80%)>Cladosporium cladosporioides (21.20%)> Aspergillus flavus (20.80%)>Penicillium rubrum (20.40%)> Curvularia lunata (19.30%)>Aspergillus fumigatus (17.50%).
| Table 3: | Effect of different temperatures on soluble crude protein (%) production by dominant green manure decomposing fungi |
 | |
| Table 4: | Per cent biomass reduction of Sesbania aculeata L. substrate during soluble crude protein production by dominant decomposing fungi |
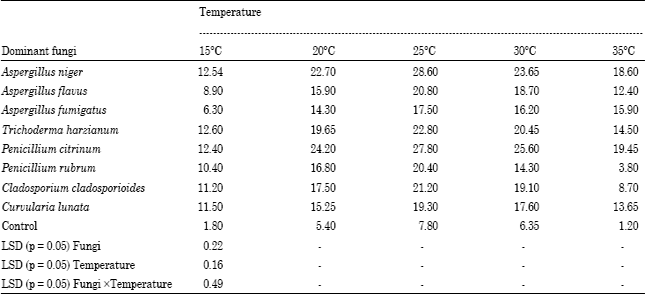 | |
1Dry weight of substrate + fungal mycellium | |
| Table 5: | Effect of different nitrogen sources on SCP* production by Penicillium citrinum |
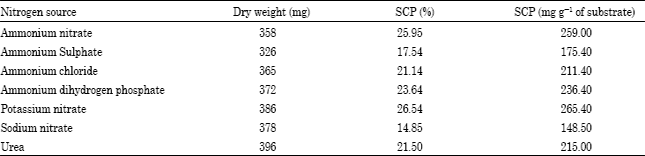 | |
*Soluble crude protein | |
Among seven different nitrogen sources tested against Penicillium citrinum, the highest producer of SCP, potassium nitrate was found to be the best for maximum SCP production (26.54%) whereas the least suitable nitrogen source for SCP production by Penicillium citrinum was recorded to be Sodium nitrate (14.85%) (Table 5) Similar results were reported by Dhillon and Chahal (1978) and Vibha and Sinha (2005).
DISCUSSION
Table 1 and 2 suggested that the more CO2 release took place in un-sterilized condition perhaps due to the presence of mixed species of green manure decomposers. The CO2 evolution was significantly higher in Aspergillus niger in both conditions due better adaptability and utilization pattern of the substrate. CO2 flux is an indicator of decomposition rates and activities of the microbes involved in the decomposition of substrate. Amendment with corn stover increases the flux and cumulative CO2 release compared with the un-amended (Pangga et al., 2000; Saha et al., 2003; Johnson et al., 2004; Saha and Hajra, 2004; Chandra et al., 2007). CO2 evolution in soil from green manures (Sesbania aculeata L.) has been investigated by Bedi et al. (2009). They found highest cumulative CO2 evolution by Sesbania aculeata + 50% nitrogen + compost and lowest CO2 release was found in case of recommended dosages of nitrogen under both aerobic and un-aerobic conditions.
Table 3 suggested that the Penicillium citrinum was potential producer of soluble crude protein at 25° while Penicillium rubrum produced minimum per cent of soluble crude protein at 35°C. Similar results were earlier reported by Chahal and Grey (1969), Dhillon et al. (1980) and Vibha and Sinha (2005). Rapid and higher production of SCP by fungi from delignified cellulose may be due to availability of amorphous form of cellulose owing to delignification with sodium hydroxide. Punj et al. (1971) from their detailed studies of decomposition of delignified cellulose reported that delignification of residue with sodium hydroxide results in the increase in amorphous form of cellulose which is readily attacked by fungi. Dhillon et al. (1980) reported similar reason for increase in SCP from delignified cellulose with sodium hydroxide. Iyayi (2004) reported highest increase in per cent protein obtained with Aspergillus niger, A. flavus and Penicillium sp. when agro-industrial by products were separately inoculated with these fungi.
The data presented in Table 4 shows that the fungi have different ability of production of fungal biomass and utilization of green manure substrate. The maximum per cent biomass reduction observed by Aspergillus niger (28.60%) at 25°C and minimum was found in Penicillium rubrum (3.80%) at 35°C. This is consistent with the finding of Chahal and Grey (1969) and Vibha and Sinha (2005). The data given in the Table 5 revealed the effect of different nitrogen sources on the soluble crude protein production by Penicillium citrinum. During the experiment potassium nitrate was recorded best nitrogen source whereas sodium nitrate was found poorest nitrogen source for soluble crude protein by Penicillium citrinum. Similar results were reported by Dhillon and Chahal (1978) and Vibha and Sinha (2005). In their experiment on soluble crude protein production, Vibha and Sinha (2005) tested eight different nitrogen sources against Trichoderma harzianum and reported almost similar results.
ACKNOWLEDGMENTS
Authors thank Head, Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi for laboratory facilities. Their sincere thanks are also due to University Grant Commission, Govt. of India, for the financial support.
REFERENCES
- Chandra, R., N. Kumar and A.K. Tyagi, 2007. Nutrient dynamics and decomposition rates during composting of sulphitation pressmud by different methods. J. Environ. Sci. Eng., 49: 183-188.
Direct Link - Hafeez, F.Y., F. Naeem, N. Shaheen and K.A. Malik, 2007. Nodulation of Sesbania spp. by introduced Rhizobia in competition with naturalized strains in different soil types. Pak. J. Bot., 39: 919-929.
Direct Link - Hobbie, E.A., L.S. Watrud, S. Maggard, T. Shiroyama and P.T. Rygiewicz, 2003. Carbohydrate use and assimilation by litter and soil fungi assessed by carbon isotopes and BIOLOG (R) assays. Soil Biol. Biochem., 35: 303-311.
CrossRef - Iyayi, E.A., 2004. Changes in the cellulose, sugar and crude protein contents of agro-industrial by-products fermented with Aspergillus niger, Aspergillus flavus and Penicillium sp. Afr. J. Biotechnol., 3: 186-188.
Direct Link - Johnson, J.M.F., D. Reicosky, B. Sharratt, M. Lindstorm, W. Voorhess and L.C. Boggs, 2004. Characterization of soil amendment with by-product of corn stover fermentation. Soil Sci. Soc. Am. J., 68: 139-147.
Direct Link - Nayyar, V.K. and I.M. Chhibba, 2000. Effect of Green Manuring on Micronutrient Availability in Rice-Wheat Cropping System of Northwest India. In: Long-Term Soil Fertility Experiments in Rice-Wheat Cropping Systems, Abrol, I.P., K.F. Bronson, J.M. Duxbury and R.K. Gupta (Eds.). Rice-Wheat Consortium for the Indo-Gangetic Plains, New Delhi, India, pp: 68-72.
- Okeke, A.I. and C.P.E. Omaliko, 1992. Leaf litter decomposition and carbon di oxide evolution of some agroforestry fallow species in southern Nigeria. For. Ecol. Manage., 50: 103-116.
CrossRef - Pangga, G.V., G. Blair and R. Lefroy, 2000. Measurement of decomposition and associated nutrient release from straw (Oryza sativa L.) of different rice varieties using a perfusion system. Plant Soil, 223: 1-11.
Direct Link - Pernas, H., 1975. Model for decomposition of organic matter by microorganisms. Soil Biol. Biochem., 7: 161-169.
CrossRef - Witkamp, M., 1969. Cycles of temperature and carbon dioxide evolution from litter and soil. Ecology, 50: 922-924.
Direct Link - Witkamp, M., 1966. Decomposition of leaf litter in relation to environment, microflora, and microbial respiration. Ecology, 47: 194-201.
Direct Link - Witkamp, M., 1966. Rates of carbon dioxide evolution from the forest floor. Ecology, 47: 492-494.
Direct Link - Zaccheo, P., G. Cabassi, G. Ricca and L. Crippa, 2002. Decomposition of organic residues in soil: Experimental technique and spectroscopic approach. Organic Geochem., 33: 327-345.
CrossRef