
S. Angra
Department of Botany, Panjab University, Chandigarh 160 014, India
S. Kaur
Department of Botany, Panjab University, Chandigarh 160 014, India
K. Singh
Department of Botany, Panjab University, Chandigarh 160 014, India
D. Pathania
Department of Botany, Panjab University, Chandigarh 160 014, India
N. Kaur
Department of Botany, Panjab University, Chandigarh 160 014, India
S. Sharma
Department of Botany, Panjab University, Chandigarh 160 014, India
H. Nayyar
Department of Botany, Panjab University, Chandigarh 160 014, India
International Journal of Agricultural Research
Year: 2010 | Volume: 5 | Issue: 6 | Page No.: 328-345







ABSTRACT
The objective of the present study was to elucidate the metabolic bases of sensitivity of soybean genotypes during seed filling to water stress that are otherwise least understood. Here, two contrasting soybean genotypes Pb1 (stress-tolerant) and Bragg (stress-sensitive) were water stressed by holding 50% of the water compared to controls for 10 days during the onset of seed filling stage (R5 stage). These genotypes were assessed for profiles of osmolytes and antioxidants during the course of water stress in the leaves. While, both the genotypes did not differ much till 4th or 6th day of stress, the differences appeared thereafter and were prominent at 10 day (last day) of stress. In general, Pb1 genotypes experienced lesser extent of stress injury and retained greater amount of water. This was associated with higher accumulation of sucrose, free amino acids, soluble proteins and proline by this genotype as compared to Bragg, which accumulated greater content of reducing sugars. The oxidative damage as lipid peroxidation was similar in both the genotypes but Bragg genotype had higher hydrogen peroxide levels. The antioxidants like ascorbic acid, catalase and ascorbate peroxidase were expressed greatly in Pb1 genotype than in Bragg genotype while superoxide dismutase activity did not differ between the two genotypes. It was indicated that Pb 1 genotype was affected to a lesser extent by water stress due to its superior capacity to generate osmolytes and antioxidants. Additionally, the seed components such as starch and proteins of the stressed plants experienced less damage in Pb1 genotype.
PDF Abstract XML References Citation
Received: March 18, 2010;
Accepted: May 10, 2010;
Published: June 10, 2010
How to cite this article
S. Angra, S. Kaur, K. Singh, D. Pathania, N. Kaur, S. Sharma and H. Nayyar, 2010. Water-Deficit Stress During Seed Filling in Contrasting Soybean Genotypes: Association of Stress Sensitivity with Profiles of Osmolytes and Antioxidants. International Journal of Agricultural Research, 5: 328-345.
DOI: 10.3923/ijar.2010.328.345
URL: https://scialert.net/abstract/?doi=ijar.2010.328.345
DOI: 10.3923/ijar.2010.328.345
URL: https://scialert.net/abstract/?doi=ijar.2010.328.345
INTRODUCTION
Water-deficit is a major abiotic stress limiting soybean production (Desclaux et al., 2000; Lobato et al., 2008). The growth stages in soybean can be characterized precisely and hence offer an excellent model to study the impact of climatic factors on its production. The various growth stages of soybean respond differently to water stress to affect its yield potential (Desclaux et al., 2000;Egli and Bruening, 2004). The reproductive stage is reported to be the most sensitive to water stress and hence detrimental for soybean yield (Liu et al., 2004a; Dogan et al., 2007). The reproductive stage encompasses early flowering, full bloom, beginning pod, full pod, beginning seed and full seed (Desclaux et al., 2000). Previous findings indicate greater damage to yield (up to 46%) in soybean plants stressed at seed filling stage compared to stress at other reproductive stages (Eck et al., 1987). Subsequently, Brevedan and Egli (2003) reported that short periods of water stress during seed filling caused substantial yield reduction (39%) due to fewer and smaller seeds in soybean. These authors also reported decrease in chlorophyll, carbon exchange rate and carbohydrates in water-stressed soybean plants that limited the seed yields. Liu et al. (2004b) associated reduction in carbohydrate concentration in drought-stressed soybean plants with decreased pod set.
The metabolic reasons governing the water stress sensitivity in soybean at reproductive stage are least known. Turgor maintenance is a primary mechanism to deal with water stress that involves osmoregulation (Chaves and Oliveira, 2004). Osmolytes like proline, sugars and amino acids that contribute towards osmoregulation in crops including soybean (Adejare and Umebese, 2008) are reported to govern the stress sensitivity (Ashraf and Foolad, 2007; Nayyar and Walia, 2004; Kocsy et al., 2005). Thus, in response to drought stress, significant variations were observed in expression of osmolytes such as proline, sugars and amino acids in various crops such as pea (Sanchez et al., 1998) and wheat (Nayyar and Walia, 2004; Kameli and Losel, 1995; Kerepesi and Galiba, 2000) and many other species (Ashraf and Foolad, 2007). Water stress also evokes oxidative stress (Turkan et al., 2004); to counter it the plants activate a variety of antioxidants (Noctor and Foyer, 1998). The expression of these antioxidants determines the stress response that varies among the genotypes of various crop species (Sairam et al., 1998). Thus, in drought-genotypes, the activities of enzymatic antioxidants such as superoxide dismutase, catalase, ascorbate peroxidase and levels of non-enzymatic antioxidants such as ascorbic acid and glutathione were significantly higher tha their sensitive counterparts (Sairam et al., 1998; Kocsy et al., 2005; Turkan et al., 2004; Nayyar and Gupta, 2006; Zhou et al., 2006).
Understanding these mechanisms indicates the defense strategy of a crop species in dealing with water stress. Information on status of osmolytes and antioxidants in soybean genotypes subjected to water stress during seed filling is lacking. One of the ways of probing the stress response is to compare the differentially sensitive genotypes at metabolic levels assuming to get clues about the traits governing the stress sensitivity. Thus, the present study was conducted on the contrasting soybean genotypes (Pb1-stress tolerant) and Bragg (stress-sensitive), which were water stressed at seed filling stage, to elucidate the mechanisms related to tolerance against water stress. It was hypothesized that variations in the generation of osmolytes and antioxidants might determine the stress sensitivity of these genotypes.
MATERIALS AND METHODS
The experiment was conducted in the year 2008 (June to October) at Panjab University, Chandigarh. The seeds of soybean genotypes Pb1 (drought-tolerant) and Bragg (drought-sensitive) were raised in earthen pots (height 30 cm , diameter 25 cm) having a mixture of air dry soil and 5 g of a compound N6P20K10 fertilizer to remove any nutrient deficiency as a limiting factor (Cissé et al.,1996). The seeds were inoculated with Bradyrhizobium japonicum; four seeds were planted in each pot in November and, after emergence, the plants were thinned to two per pot. The plants were grown under natural conditions and protected from rains. At R5 (beginning seed) stage, the plants were subjected to short-term water stress for 10 days by withholding 50% of the water compared to the amount provided to the control plants that continued to get irrigation. The addition of water in the pots was adjusted by their regular weighing. Upon completion of the stress period, the fully-irrigated pots had a soil moisture content of 35-39% compared to water stressed ones which had 17-20% moisture at the completion of stress period (10 day). The plants were examined at different intervals for the following parameters:
Stress Injury
The leaves of the water-stressed plants were examined for stress injury as membrane damage (Lutts et al., 1996), chlorophyll content (Arnon, 1949) and relative leaf water content (Barrs and Weatherly, 1962) as elaborated elsewhere (Nayyar and Gupta, 2006).
Stress Index
Stress index was also obtained from the relationship as follows (Fischer and Maurer, 1978): Is = 1-(Ystr/Ycont), where, Ycont is the mean total yield (seed number/plant) at harvest under well-watered conditions and YStr the mean total yield under water-stress conditions. Stress index is important in translating the effect of water stress on plant yield. The more it approaches unity (1), the more depressive the effect of stress on the yield component (Ndunguru et al., 1995). On the other hand, as Is approaches zero, the higher the resistance capacity of a plant to drought.
Sucrose
For the sucrose assay, samples were extracted with 80% ethanol at 80°C three times each for 1.5 h. The extracts were combined, evaporated at 40°C in an air-circulating oven and used for the sucrose assay. Enzymatic assays for sucrose were performed using the spectrophotometric method of Jones et al. (1977). Aliquots of 200 μL from standard sucrose and samples were added to 1 mL of reaction mixture composed of imidazole buffer 100 mm (pH 6.9), (40 mM imidazole base, 60 mM imidazole-HCI), 0.4 mM NADP+, 1 mM ATP, 5 mM MgC12, 0.5 mM dithiothreitol, 0.02% (w/v) BSA, 20 μg mL-1 yeast invertase (EC3.2.1.26), 2 μg mL-1 yeast hexokinase (EC 2.7.1.1), 1 μg mL-1 yeast P-glucoisomerase (EC 5.3.1.9) and incubated for 30 min at 25°C to allow conversion of glucose and fructose to glucose 6-P. The absorption was read at 340 nm. Eighty five microliters of glucose-6-P dehydrogenase (70 U mL-1) added, mixed well and re-read after about 5 min when absorbance became constant. Blanks were run with 200 μL of extract and 1 mL of reaction mixture without invertase. Readings obtained from each sample were converted to sucrose concentration employing a standard curve.
Reducing Sugars
The oven-dried plant material was homogenized in hot ethanol (80%) and centrifuged at 2000 rpm for 10 min. Supernatant was clearly decanted off. Three milliliter of ethanol (80%) was added to the residue and recentrifuged. The extraction was repeated twice to ensure the complete recovery of sugars. The residue was kept for further analysis of starch. The supernatant was pooled and evaporated to dryness in China dish on a boiling water bath. The residue was eluted with 5 mL of 20% ethanol and subjected to analysis for reducing sugars and free amino acids. For estimation of reducing sugars, to 1 mL of ethanol extract (prepared as above) 1 mL of DNSA reagent was added. The reaction mixture was boiled for 12 min. Two molliter of distilled water was added and absorbance was recorded at 560 nm against a blank containing 80% ethanol in place of ethanol extract. The concentration of reducing sugars (mg g-1 DW) was calculated from a standard curve plotted with known concentration of glucose (Sumner and Howell, 1935). The starch content was measured by acid hydrolysis method given by McCready et al. (1950). The residue of ethanol extract (used in above estimations) washed with 80% ethanol to remove all the traces of soluble sugars. To the residue, 5 mL of distilled water and 6.5 mL of 52% perchloric acid was added to extract the starch by placing the samples at 0°C for 20 min. The mixture was centrifuged and retained the extract. The process was repeated 3-4 times using fresh perchloric acid and diluted to final volume 100 mL. To 0.5 mL of diluted extract, 4.5 mL of distilled water was added followed by addition of 10 mL of cold anthrone sulfuric acid reagent in ice bath. The sample mixture was heated at 100°C for 8 min and cooled rapidly to room temperature. The absorbance was measured at 630 nm. The final content of starch was calculated from a standard curve plotted with known concentration of glucose. The estimation of free amino acid content was done by method of Lee and Takahashi (1966). Ninhydrin reagent (3.8 mL) was added to 1 mL of ethanol extract and the contents were shaken vigorously. The mixture was heated in boiling water bath for 12 min and cooled in running tap water to room temperature. The absorbance of the coloured solution was read at 570 nm against a blank containing 80% ethanol. The concentration of free amino acid (mg g-1 DW) was calculated from a standard curve plotted with known concentration of glycine.
Proline and Soluble Proteins
The proline and soluble proteins content was examined using the methods of Bates et al. (1973) and Lowry et al. (1951), elaborated elsewhere (Nayyar and Gupta, 2006).
Lipid Peroxidation and Antioxidants
The lipid peroxidation was measured in terms of thiobarbituric acid-reactive substances (TBARS), the products of lipid peroxidation according to the method of Heath and Packer (1968). The leaf samples (1 g) were homogenized in 10 mL of 0.1% trichloroacetic acid. The homogenate was centrifuged at 15,000 g for 5 min. Four milliliter of 0.5% thiobarbituric acid in 20% trichloroacetic acid was added to a 1 mL aliquot of the supernatant. The mixture was heated at 95°C for 30 min and then quickly cooled in an ice bath. After centrifugation at 10,000 g for 10 min, the absorbance was recorded at 532 nm. The value for non-specific absorption at 600 nm was subtracted. The content was calculated using its absorption coefficient of 155 mmol-1 cm-1 and expressed as μmol g-1 weight.
Enzymes Assays
For enzyme extracts and assays, the leaves were frozen and then ground in 4 mL solution containing 50 mM phosphate buffer (pH 7.0), 1% (w/v) polyvinylpolypyrrolidone and 0.2 mM ascorbic acid (ASA). The homogenate was centrifuged at 15,000 g for 30 min and supernatant was collected and used for enzyme assays. The activity of superoxide dismutase was measured according to the method of Giannopolitis and Ries (1977). The assay medium contained 50 mM phosphate buffer (pH 7.8), 13 mM methionine, 75 mM p-nitro blue tetrazolium chloride (NBT), 2 mM riboflavin, 0.1 mM EDTA and 5 mL enzyme extract. One unit of enzyme activity was determined as the amount of the enzyme to reach an inhibition of 50% NBT reduction rate by monitoring the absorbance at 560 nm. The activity of catalase was determined as a decrease in absorbance at 240 nm for 1 min following the decomposition of H2O2 (Chance and Maehly, 1955). The reaction mixture contained 50 mM phosphate buffer (pH 7.0) and 15 mM H2O2. The activity of ascorbate peroxidase was measured as a decrease in absorbance at 290 nm for 1 min (Nakano and Asada, 1981). The assay mixture consisted of 0.5 mM ascorbic acid, 0.1 mM H2O2, 0.1 mM EDTA, 50 mM sodium phosphate buffer (pH 7.0) and 0.15 mL enzyme extract. Ascorbic acid was measured by the method of Mukherjee and Choudhuri (1985). Leaf material was extracted with 10 mL of 6% trichloroacetic acid. The extract was mixed with 2 mL of 2% dinitrophenylhydrazine (in acidic medium) followed by addition of 1 drop of 10% thiourea (in 70% ethanol). The mixture was boiled for 15 min in a water bath and after cooling at room temperature, 5 mL of 80% (v/v) H2SO4 was added to the mixture at 0°C. The absorbance was recorded at 530 nm. The concentration of ascorbic acid was calculated from a standard curve plotted with known concentration of ascorbic acid. For measurement of glutathione content, fresh tissue was homogenized in 2 mL of 2% metaphosphoric acid and centrifuged at 17,000 g for 10 min. The aliquots of the supernatant were neutralized by adding 0.6 mL of 10% sodium citrate to 0.9 mL of the extract. A total volume of 1 mL of assay containing 700 μL NADPH (0.3 mmol L-1), 100 μL DTNB (6 mmol L-1), 100 μL distilled water and 100 μL of extract was prepared and stabilized at 25°C for 3-4 min. Later 10 μL of glutathione reductase was added and the absorbance was measured at 412 nm. Glutathione was calculated from a standard graph as described by Griffifh (1980). The activities of amylases was assayed using the method of Shuster and Gifford (1962) while that of invertase was measured using the method of Nygaard (1977).
Seed Composition
The developing seeds after 5th day of stress collected from control and stressed plants of both the genotypes and analyzed for enzymes according to the methods described above. The seed reserves were estimated from mature seeds according to AOAC (1990) procedures.
The observations were replicated (n = 3) and standard errors were worked out, mean values±SE are presented. Data was subjected to ANOVA and Critical Difference (CD) was worked out for genotypes and days of stress, using AGRISTAT software.
RESULTS
Yield
Water stress resulted in greater reduction in seed yield in Bragg (46% over control) relative to Pb1 genotype (28% over control) (Fig. 1). Consequently, the stress sensitivity index was appreciably greater in Bragg (0.45) compared to Pb1 genotype (0.22).
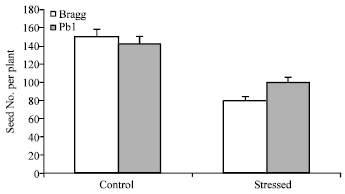 | |
| Fig. 1: | Seed number per plant in control and water stressed plants of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 day. Mean values±SE (n = 3) (CD (5%): 2.64 |
Stress Injury
The stress injury was measured as increase in Electrolyte Leakage (EL), Relative Leaf Water Content (RLWC) and chlorophyll loss during stress period (Fig. 2). The Bragg genotype experienced 18% higher EL than Pb1 genotype on 10th day of stress (Fig. 2). The relative leaf water content was 19% more in Pb1 genotype than Bragg genotype on this day. The Bragg genotype lost about 50% Chl over control compared to 34% loss in Pb1 genotype on last day of stress.
Carbohydrates
Enzymes
The activity of amylases (α- and β) and invertase (Fig. 3) showed increase as the stress duration increased. Pb1 genotypes possessed about 23 and 20% greater activity of amylases and invertase, respectively than Bragg genotype during 6th day onwards.
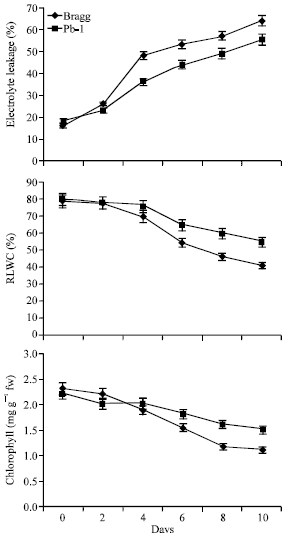 | |
| Fig. 2: | Electrolyte leakage (EL), Relative Leaf Water Content (RLWC) and chlorophyll (Chl) content in leaves of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 d. Mean values ±SE (n = 3), (CD (5%): for Electrolyte leakage-Genotypes (2.6) for days (2.3); for RLWC-Genotypes (3.5), days (2.6); Chlorophyll- Genotypes (0.18) for days (0.16) |
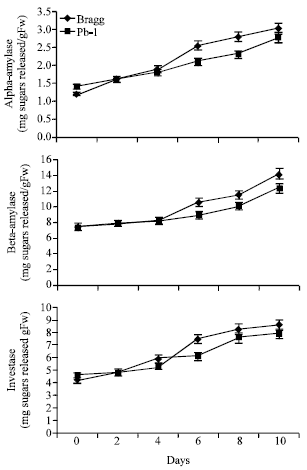 | |
| Fig. 3: | Amylases and invertase activity in leaves of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 d. Mean values±SE (n = 3), (CD (5%): α-amylase-Genotypes (0.2) for days (0.16); β-amylase-Genotypes (0.31), days (0.52); Invertase- Genotypes (0.35) for days (0.26) |
Starch
The starch content of the leaves began to decrease on 4th day of stress in both the genotypes (Fig. 4). The extent of decrease was greater in Bragg genotype from 4th day onwards. On last day, the stressed plants of Pb1 possessed 23% greater starch content than Bragg genotype.
Sucrose
The sucrose content decreased due to stress in both the genotypes with Bragg genotype showing greater reduction than Pb 1 genotype (Fig. 4). The differences were significant between the two genotypes from 6th day onwards. On 10th day the Pb1 genotype possessed about 21% greater sucrose than Pb 1 genotype.
Reducing Sugars
The content of reducing sugars showed increase in the stressed plants of both the genotype; it increased with duration of the stress (Fig. 4). The Bragg genotype showed greater accumulation of reducing sugars from 6th day of stress and onwards. On 10th day, the Bragg genotype possessed 20% greater content of reducing sugars.
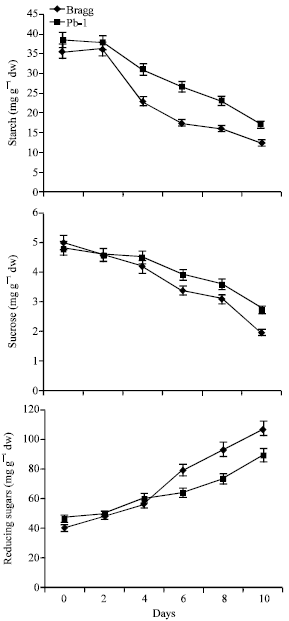 | |
| Fig. 4: | Starch, sucrose and reducing sugars in leaves of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 d. Mean values±SE (n = 3). (CD (5%): Starch-Genotypes (2.6), days (2.3); Sucrose-Genotypes (2.5), days (1.9); Reducing sugars-Genotypes-(0.21) for days (0.18) |
Non Structural Carbohydrates
The non-structural carbohydrates involving sucrose, reducing sugars and starch showed increase with progress of starch in both the genotypes (Fig. 5). The differences between the two genotypes surfaced on 6th day and Bragg genotype possessed higher content till the last day of stress. Though sucrose and starch were lesser in Bragg than Pb1 genotype, the higher content of reducing sugars in the former genotype made the difference.
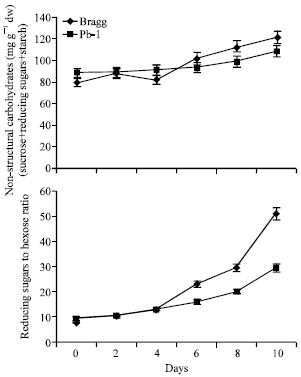 | |
| Fig. 5: | Non structural carbohydrates and reducing sugars to hexose ratio in leaves of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 day. Mean values±SE (n = 3). (CD (5%): Non structural carbohydrates-Genotypes (2.6), days (1.8); Reducing sugars to hexose ratio-Genotypes (2.7), days (2.1) |
Reducing Sugars to Sucrose Ratio
The ratio of reducing sugars to sucrose increased with stress duration that was concomitant with decrease in sucrose and increase in reducing sugars (Fig. 5). The differences were negligible between the two genotypes till 4th day of stress. Thereafter, the Bragg genotype had markedly greater ratio and on 10th day where it was observed to be 52 in Bragg compared to 30 in case of Pb1 genotype.
Nitrogenous Components
Soluble Proteins
The soluble proteins increased with stress duration in both the genotypes. The extent of increase was greater in Pb1 genotype than in Bragg genotype (Fig. 6). The Pb1 genotype possessed higher soluble proteins from 4th day onwards. In this genotype, the increase occurred till 8th day of stress while in Bragg genotype, the proteins began to decrease from 6th day onwards.
Free Amino Acids
The content of free amino acids increased with progression of the stress in both the genotypes (Fig. 6). The increase was noticeable till 8th day of stress in Pb1 while in Bragg it occurred till 6th day. Thereafter, the content decreased and reached markedly lower levels on 10th day. Pb 1 genotype had higher content of free amino acids than Bragg on this day.
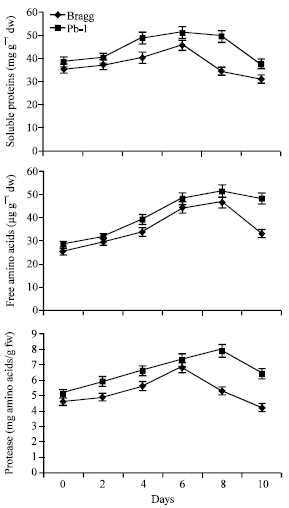 | |
| Fig. 6: | Soluble proteins, free amino acids and proteases in leaves of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 d. Mean values±SE (n = 3). (CD (5%): Proteins-Genotypes (1.3) for days (1.8); Free amino acids- Genotypes (1.6), days (1.9). Protease-Genotypes (0.63), Days (0.58) |
Protease
The activity of protease showed increase with stress exposure and reached its peak on 6th day in Bargg and 8th day in Pb1 genotype thereafter a decease was observed (Fig. 6) Pb1 genotype possessed significantly greater activity than Brag genotype on 8th and 10th day of stress.
Oxidative Damage
The oxidative damage was measured in terms of malondialdehyde (MDA) and hydrogen peroxide (H2O2) content (Fig. 7). The oxidative damage became significant on 4th day of stress and increased subsequently in both the genotypes. The differences between the two genotypes for MDA content were negligible but for H2O2 content, the Bragg genotype had greater content especially from 6th day onwards during stress indicating greater oxidative damage in this genotype.
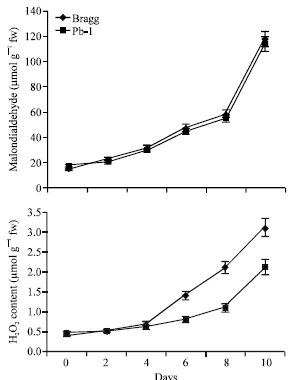 | |
| Fig. 7: | Malondialdehyde and hydrogen peroxide content in leaves of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 day. Mean values±SE (n = 3). (CD (5%): Malondialdehyde-Genotypes (1.6), days (2.4); Hydrogen peroxide-Genotypes (0.82), days (0.91) |
Anti-Oxidants
The antioxidants, superoxide dismutase (SOD; converts the superoxide radicals into hydrogen peroxide), catalase (CAT; catalyses the removal of hydrogen peroxide) and ascorbic acid (ASC) were analyzed in both the genotypes (Fig. 8). The activity of these enzymes showed increase in both the genotypes from 2nd day of stress onwards and remained high till 8th day. It began to decrease thereafter. The differences for SOD were insignificant between the two genotypes but the activity of CAT was greater in Pb1 genotype than in Bragg genotype from 6thday of stress onwards. Ascorbate peroxidase (APO) activity like other antioxidants increased in both the genotypes and its activity was greatest on 6th day of stress with Pb1 genotype showing higher activity level than Bragg. The activity declined on 10th day of stress in both the genotypes with Pb1 genotype maintaining higher levels (Fig. 8).
Ascorbic acid (ASC) content increased with stress duration till 8th day and decreased thereafter in both the genotypes. ASC was about 19% higher in Pb1 genotype than in Bragg genotype on 8th day of stress (Fig. 9). The proline content began to increase with stress in both the genotypes (Fig. 9). The differences were insignificant between the two genotypes till 4th day of stress. After that, it increased to a markedly greater extent in Pb1 genotype than in Bragg genotype and on 10th day of stress, Pb1 genotype possessed about 26% greater proline content than the Bragg genotype.
Seed Composition
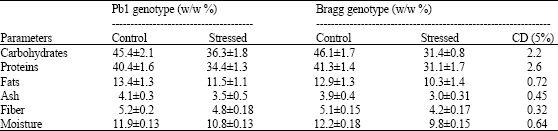
The proximate analysis of seeds collected from control and stressed plants indicated a significant reduction in carbohydrates, proteins, fats, ash, fiber content and seed moisture in both the genotypes (Table 1). In the seeds of the stressed plants, carbohydrates decreased to a greater extent in Bragg (31.4% W/w) compared to Pb1 genotype (36.3% W/w). The proteins also showed larger decrease in Bragg (31.4% W/w) than in Pb1 (34.4% W/w) genotype.
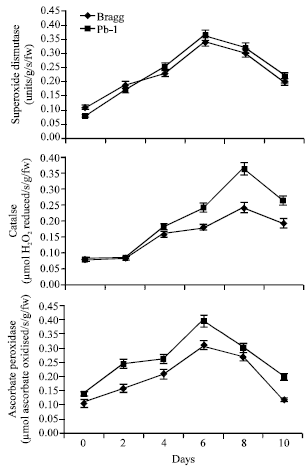 | |
| Fig. 8: | Superoxide dismutase, catalase and ascorbate peroxidase activity in leaves of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 day. Mean values±SE (n = 3). (CD. (5%): Superoxide dismutase-Genotypes (0.13) for days (0.11); Catalase- Genotypes (0.48) for days (0.56); Ascorbtae peroxidase-genotypes (0.09), days-0.061) |
| Table 1: | Proximate composition of seeds harvested at physiological maturity from plants water-stressed at R5 stage. Mean values ± SE (n = 3) |
 | |
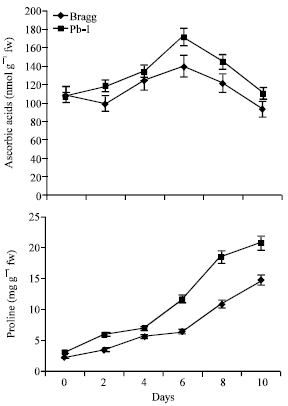 | |
| Fig. 9: | Ascorbic acid and proline content in leaves of Bragg (drought sensitive) and Pb1 (drought tolerant) soybean genotypes. The plants were water stressed during seed filling for 10 days. Mean values±SE (n = 3). (CD (5%): Ascorbic acid-Genotypes (3.7), days (4.1); Proline-Genotypes (1.3), days (1.6) |
The fat content reduced largely in Bragg genotype as compared to Pb1 genotype. The Bragg genotype also possessed lesser amount of ash and fiber than Pb1 genotype. The Bragg genotype had relatively less moisture content than Pb1 genotype.
DISCUSSION
The findings revealed that water-deficit during seed development considerably limited the soybean yield, which is in agreement with earlier findings on this crop (Desclaux and Roumet, 1996; Brevedan and Egli, 2003). The decrease due to stress in seed yield occurred due to reduction in seed size and number that was in agreement with some previous studies (Brevedan and Egli, 2003). Out of the two genotypes, Bragg experienced greater reduction in seed yield than Pb 1 genotype suggesting higher stress sensitivity of the former genotype. This was corroborated by greater stress injury (measured as electrolyte leakage) and loss of chlorophyll in Bragg. Pertinently, during stress treatment, Bragg also retained less relative leaf water content compared to Pb1 that might be a vital reason contributing towards greater stress-induced damage to the former genotype. Thus, the differences in the seed yield between the two genotypes could primarily be related to variation in their leaf water content.
The metabolic reasons associated with differential water stress sensitivity among soybean genotypes are not known that were examined in the present study. We worked on the expression of endogenous level of osmolytes like proline, free amino acids and sugars and their related enzymes, which have a role in turgor generation or its maintenance during water stressed situation. Additionally, we compared the oxidative damage due to water stress between the two genotypes. It was hypothesized that the differential sensitivity of the two contrasting genotypes might be related to variations in osmolytes and antioxidants.
Water stress decreased the sucrose and starch content in both the genotypes, which is in accordance with the previous findings of Brevedan and Egli (2003) and Liu et al. (2004a, b) in drought-stressed soybean. The extent of decrease in these molecules was higher in Bragg genotype than in Pb1 genotype. The content of sucrose and starch in leaves depends upon the photosynthetic ability during stress (Praxedes et al., 2006) and in this context, Pb1 genotype appeared to be superior to Bragg genotype. This was also evident from the presence of greater chlorophyll and sucrose content in Pb1 genotype during stress period. These findings get support from our previous studies on wheat (Nayyar and Kaushal, 2002) that indicated that the stress-tolerant genotypes maintain higher Pn under stressed situations than their counterparts owing to greater stability of their photosynthetic apparatus or due to their ability to maintain higher chlorophyll content. Alternatively, the faster breakdown of starch and sucrose might also reduce their levels. We noticed here elevation in activities of starch and sucrose hydrolyzing enzymes like amylases and invertases during stress period. Relatively, their activity levels were greater in Bragg genotype than in Pb1 genotype, which explains larger reduction in content of these molecules in stressed plants of the Bragg genotype. The resultant products-reducing sugars (glucose and fructose) have a significant role in the osmotic adjustment (Morgan, 1992) and contribute towards improvement in turgor status (Morgan, 1992; Kerepesi and Galiba, 2000). Here, we found in spite of higher content of reducing sugars in Bragg genotype, its leaf water content was lesser than in Pb1 genotype. This implied that osmolytes other than reducing sugars might have greater role towards turgor generation in Pb1 genotype. There are contrasting observations on the status of reducing sugars in differentially sensitive genotypes of other crop species. While some studies report that the tolerant genotypes of a crop species possessed higher content of reducing sugars (Nayyar and Walia, 2004; Kerepesi and Galiba, 2000), converse was true for other cases (Kameli and Losel, 1995). Moreover, stomatal controls might be better in Pb1 genotype thus minimising water loss, which needs to be examined. Nevertheless, the elevation of sugars might have implications in energy requirement (Baena-Gonzalez et al., 2007).
The content of free amino acids was greater in Pb1 genotype than in Bragg genotype which coincided with greater protease activity in this genotype (Fig. 6). Additionally, the proline content was also significantly greater in Pb1 genotype than in Bragg genotype. Both free amino acids and proline have major involvement in turgor generation (Sánchez et al., 1998). These findings are in agreement with earlier studies where the tolerant genotypes of a crop species have been reported to possess greater contents of the free amino acids and proline than the sensitive ones (Nayyar and Walia, 2004; Kocsy et al., 2005). The superior water status in Pb1 genotype during stress might possibly be attributed to higher content of these molecules in this genotype than in Bragg. The level of soluble proteins showed increase initially and decrease subsequently in both the genotypes but their values were greater in Pb1 genotype. It seems that the initial increase in total soluble proteins during drought stress might be due to the expression of new stress proteins, while the decrease may be attributed to their hydrolysis and degradation (Hajheidari et al., 2005). These findings are in concurrence with the previous studies reporting the increase or decrease in soluble proteins under stress conditions (Riccardi et al., 1998). Here, the protein levels did not show decrease with rise in proteases activity during stress conditions in both the genotypes. The increased protein levels might occur due to induction of biosynthesis of stress proteins that act as chaperones and protect the membranes and enzymes (Jiang and Huang, 2002). In this regard, the Pb1 genotype was superior in having higher content of soluble proteins than Bragg that is in agreement with previous studies reporting greater soluble proteins in tolerant genotypes (Labhilili et al., 1995).
The oxidative damage in the leaves of the stressed plants was observed as rise in levels of malondialdehyde (MDA) and hydrogen peroxide (H2O2) content that is in concurrence with previous such studies (Chaves and Oliveira, 2004). While MDA content did not vary significantly during stress between the two genotypes, the H2O2 content was significantly higher in Bragg genotype on 10th day of stress. Our findings get support from an earlier study by Sairam et al. (1998) who also observed H2O2 content to be higher in a drought-susceptible wheat genotype. The non-enzymatic antioxidant, ascorbic acid was in greater content in Pb 1 genotype than its counterpart. Ascorbic acid constitutes a component in ascorbate-glutathione cycle for scavenging hydrogen peroxide (Noctor and Foyer, 1998); its greater level in the Pb1 genotype reflects a stronger antioxidant system in this genotype. These findings are in similar to some previous ones reporting greater content of ascorbate and glutathione in the tolerant genotypes of different plant species growing under water stress compared to the susceptible ones (Zhou et al., 2006). The enzyme superoxide dismutase (SOD), which converts superoxide radicals to hydrogen peroxide, did not vary significantly. These observations were similar to those of Saruyama and Tanida (1995) and Sairam et al. (1998) in contrasting genotypes of other crop species. The catalase and ascorbate peroxidase were significantly higher in Pb1 genotype than in Bragg genotype. This further corroborated that the former genotype had better ability to deal with oxidative stress, which is in accordance with our earlier findings on oxidative response of differentially sensitive genotypes (Nayyar and Gupta, 2006). In this context, our findings are similar to Oswald et al. (1992) who suggested that catalase and ascorbate peroxidase were more important in imparting tolerance against oxidative stress than SOD activity alone.
It is pertinent to mention here that though both the genotypes elevated the levels of osmoregulants and antioxidants during the stress period, Pb1 genotype was able o sustain their levels to a higher degree and for a longer duration that might be primary governing reasons for its greater drought tolerance. These observations are in agreement with findings by Sairam et al. (1998) in wheat and Nayyar and Gupta (2006) in wheat and maize.
It was further observed that the seeds of Pb1 genotype experienced less damage to their composition than Bragg genotype, which might be related to greater levels of sucrose content in the leaves of Pb1 genotype that possibly also maintained its higher mobilization to the developing seeds. The reduction in accumulation of proteins, fats and minerals in the stressed seeds possibly is associated with either impairment in the availability of their precursors or direct effects of stress on their biosynthetic enzymes (Ahmadi and Baker, 2001). Our observations are in accordance with previous ones reporting the impaired filling of seeds due to drought stress leading to reduction in their quality (Dornbos et al., 1989; Franca-Neto et al., 1993; Zalewski et al., 2001).
Thus, present findings indicated that genotype Pb1 is able to retain higher leaf turgor due to greater content of sucrose, free amino acids, proline and soluble proteins. Additionally, Pb1 genotype was able to deal with oxidative stress in a effective way than Bragg genotype. Manipulation of components of osmoregulation like proline and antioxidants like ascorbic acid may lead to induction of tolerance against water stress in soybean genotypes.
ACKNOWLEDGMENT
The partial funding from UGC, New Delhi (No. 3-213/2001/SRII) to conduct this work is gratefully acknowledged.
REFERENCES
- Adejare, F.B. and C.E. Umebese, 2008. Water stress induces cultivar dependent changes in stomatal complex, yield and osmotic adjustments in Glycine max L. Int. J. Agric. Res., 3: 287-295.
CrossRefDirect Link - Ahmadi, A. and D.A. Baker, 2001. The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat. Plant Gr. Regl., 35: 81-91.
CrossRef - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Baena-Gonzalez, E., F. Rolland, J.M. Thevelein and J. Sheen, 2007. A central integrator of transcription networks in plant stress and energy signalling. Nature, 448: 938-943.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Brevedan, R.E. and D.B. Egli, 2003. Short periods of water stress during seed filling, leaf senescence and yield of soybean. Crop Sci., 43: 2083-2088.
Direct Link - Chance, B. and A.C. Maehly, 1955. Assay of catalases and peroxidases: Catalase: 2H2O2 → 2H2O + O2 (1), Catalase and peroxidase: ROOH + AH2 → H2O + ROH + A (2). Methods Enzymol., 2: 764-775.
CrossRefDirect Link - Chaves, M.M. and M.M. Oliveira, 2004. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot., 55: 2365-2384.
CrossRefPubMedDirect Link - Desclaux, D. and P. Roumet, 1996. Impact of drought stress on the phenology of two soybean (Glycine max L. Merr) cultivars. Field Crops Res., 46: 61-70.
CrossRef - Desclaux, D., T.T. Huynh and P. Roumet, 2000. Identification of soybean plant characteristics that indicate the timing of drought stress. Crop Sci., 40: 716-722.
CrossRefDirect Link - Dogan, E., H. Kirnak and O. Copur, 2007. Effect of seasonal water stress on soybean and site specific evaluation of CROPGRO-Soybean model under semi-arid climatic conditions. Agric. Water Manage., 90: 56-62.
CrossRef - Dornbos, D.L., R.E. Mullen and R.M. Shibles, 1989. Drought stress effects during seed fill on soybean seed germination and vigor. Crop Sci., 29: 476-480.
Direct Link - Eck, H.V., A.C. Mathers and J.T. Musick, 1987. Plant water stress at various growth stages and growth and yield of soybean. Field Crops Res., 17: 1-16.
Direct Link - Egli, D.B. and W.P. Bruening, 2004. Water stress, photosynthesis, seed sucrose levels and seed growth in soybean. J. Agric. Sci., 142: 1-8.
CrossRefDirect Link - Fischer, R.A. and R. Maurer, 1978. Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust. J. Agric. Res., 29: 897-912.
CrossRefDirect Link - Franca-Neto, J.B., F.C. Krzyzanowski, A.A. Henning, S.H. West and L.C. Miranda, 1993. Soybean seed quality as affected by shriveling due to heat and drought stresses during seed filling. Seed Sci. Technol., 21: 107-116.
Direct Link - Giannopolitis, C.N. and S.K. Ries, 1977. Superoxide dismutases. I. Occurrence in higher plants. Plant Physiol., 59: 309-314.
CrossRefPubMedDirect Link - Griffith, O.W., 1980. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem., 106: 207-212.
CrossRefPubMedDirect Link - Hajheidari, M., M. Abdollahian-Noghabi, H. Askari, M. Heidari and S.Y. Sadeghian et al., 2005. Proteome analysis of sugar beet leaves under drought stress. Proteomics, 5: 950-960.
CrossRef - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - AOAC, 1990. Official Methods of Analysis. 15th Edn., Association of Official Analytical Chemists, Washington, DC., USA., pp: 200-210.
Direct Link - Jiang, Y. and B. Huang, 2002. Protein alterations in tall fescue in response to drought stress and abscisic acid. Crop Sci., 42: 202-207.
PubMed - Jones, M.G.K., W.H. Jr. Outlaw and O.H. Lowry, 1977. Enzymic assay of 10−7 to 10−14 moles of sucrose in plant tissues. Plant Physiol., 60: 379-383.
Direct Link - Kameli, A. and D.M. Losel, 1995. Contribution of carbohydrates and other solutes to osmotic adjustment in wheat leaves under stress. J. Plant Physiol., 145: 363-366.
Direct Link - Kerepesi, I. and G. Galiba, 2000. Osmotic and salt stress induced alteration in soluble carbohydrate content in wheat seedling. Crop Sci., 40: 482-487.
Direct Link - Kocsy, G., R. Laurie, G. Szalai, V. Szilagyi, L. Simon-Sarkadi, G. Galiba and J.A. de Ronde, 2005. Genetic manipulation of proline levels affects antioxidants in soybean subjected to simultaneous drought and heat stresses. Physiol. Plant., 124: 227-235.
CrossRef - Labhilili, M., P. Joudrier and M.F. Gautier, 1995. Characterization of cDNAs encoding Triticum durum dehydrins and their expression patterns in cultivars that differ in drought tolerance. Plant Sci., 112: 219-230.
CrossRef - Lee, Y.P. and T. Takahashi, 1966. An improved colorimetric determination of amino acids with the use of ninhydrin. Anal. Biochem., 14: 71-77.
CrossRefDirect Link - Liu, F., C.R. Jensen and M.N. Andersen, 2004. Drought stress effect on carbohydrate concentration in soybean leaves and pods during early reproductive development: Its implication in altering pod set. Field Crops Res., 86: 1-13.
Direct Link - Liu, F., C.R. Jensen and M.N. Andersen, 2004. Pod set related to photosynthetic rate and endogenous ABA in soybeans subjected to different water regimes and exogenous ABA and BA at early reproductive stages. Ann. Bot., 94: 405-411.
Direct Link - Lobato, A.K.S., R.C.L. Costa, C.F.O. Neto and B.G.S. Filho, 2008. Morphological changes in soybean under progressive water stress. Int. J. Bot., 4: 231-235.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Lutts, S., J.M. Kinet and J. Bouharmont, 1996. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 78: 389-398.
CrossRefDirect Link - McCready, R.M., J. Guggolz, V. Silviera and H.S. Owens, 1950. Determination of starch and amylose in vegetables. Anal. Chem., 22: 1156-1158.
CrossRefDirect Link - Morgan, J.M., 1992. Osmotic components and properties associated with genotypic differences in osmoregulation in wheat. Funct. Plant Physiol., 19: 67-76.
Direct Link - Mukherjee, S.P. and M.A. Choudhuri, 1985. Implication of hydrogen peroxide-ascorbate system on membrane permeability of water stressed Vigna seedlings. New Phytol., 99: 355-360.
CrossRefDirect Link - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Nayyar, H. and D. Gupta, 2006. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot., 58: 106-113.
CrossRefDirect Link - Nayyar, H. and D.P. Walia, 2004. Genotypic variation in wheat in response to water stress and abscisic acid induced accumulation of osmolytes in developing grains. J. Agron. Crop Sci., 190: 39-45.
Direct Link - Nayyar, H. and S.K. Kaushal, 2002. Chilling induced oxidative stress in germinating wheat grains as affected by water stress and calcium. Biol. Plant., 45: 601-604.
CrossRefDirect Link - Nygaard, P., 1977. Utilization of exogenous carbohydrates for tube growth and starch synthesis in pine pollen suspension culture. Physiol. Plant., 39: 206-210.
CrossRefDirect Link - Ndunguru, B.J., B.R. Ntare, J.H. Williams and D.C. Greenberg, 1995. Assessment of groundnut cultivars for end-of-season drought tolerance in a Sahelian environment. J. Agric. Sci., 125: 79-85.
CrossRefDirect Link - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Mol. Biol., 49: 249-279.
CrossRefPubMedDirect Link - Oswald, W.E., R. Kraus, S. Hippelli, B. Benz, R. Volpert and E.F. Elstener, 1992. Comparison of enzymatic activities of dehydroascorbic acid reductase, glutathione reductase, catalase, peroxidase and superoxide dismutase of healthy and damaged spruce needles (Picea abies L. Karst). J. Plant Physiol., 139: 742-748.
Direct Link - Praxedes, S.C., F.M. DaMatta, M.E. Loureiro, M.A.G. Ferrao and A.T. Cordeiro, 2006. Effects of long-term soil drought on photosynthesis and carbohydrate metabolism in mature robusta coffee (Coffea canephora Pierre var. kouillou) leaves. Environ. Exp. Bot., 56: 263-273.
Direct Link - Sairam, R.K., P.S. Deshmukh and D.C. Saxena, 1998. Role of antioxidant systems in wheat genotypes tolerance to water stress. Biologia Plantarum, 41: 387-394.
CrossRefDirect Link - Sanchez, F.J., M. Manzanares, E.F. de Andres, J.L. Tenorio and L. Ayerbe, 1998. Turgor maintenance, osmotic adjustment and soluble sugar and proline accumulation in 49 pea cultivars in response to water stress. Field Crops Res., 59: 225-235.
CrossRefDirect Link - Saruyama, H. and M. Tanida, 1995. Effect of chilling on activated oxygen-scavenging enzymes in low temperature-sensitive and-tolerant cultivars of rice (Oryza sativa L.). Plant Sci., 109: 105-113.
Direct Link - Sumner, J.B. and S.F. Howell, 1935. A method for determination of saccharase activity. J. Biol. Chem., 108: 51-54.
Direct Link - Shuster, L. and R.H. Gifford, 1962. Changes in 3-nucleaotidases during the germination of wheat embryo. Arch. Biochem. Biophys., 96: 532-540.
CrossRef - Turkan, I., B. Melike, F. Ozdemir and H. Koca, 2005. Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Sci., 168: 223-231.
CrossRefDirect Link - Zalewski, K., L.B. Lahuta and M. Horbowicz, 2001. The effect of soil drought on the composition of carbohydrates in yellow lupin seeds and triticale kernels. Acta Physiol. Plant., 23: 73-78.
CrossRefDirect Link - Zhou, Y.H., J.Q. Yu, W.H. Mao, L.F. Huang, X.S. Song and S. Nogues, 2006. Genotypic variation of rubisco expression, photosynthetic electron flow and antioxidant metabolism in the chloroplasts of chill-exposed cucumber plants. Plant Cell Physiol., 47: 192-199.
CrossRefDirect Link
Frank sinota Reply
nice study. can be extended to differential gene display using these cultivars to corroborate the expression of these molecules