
A.E. De Silva
School of Arts and Sciences, Monash University Malaysia, P.O. Box 8975, 46780 Kelana Jaya, Selangor, Malaysia
M.A. Kadir
Department of Agricultural Biotechnology, University Putra Malaysia (UPM), 43400 Serdang, Selangor, Malaysia
M.A. Aziz
Department of Agricultural Biotechnology, University Putra Malaysia (UPM), 43400 Serdang, Selangor, Malaysia
S. Kadzimin
Department of Crop Science, Faculty of Agriculture, University Putra Malaysia (UPM), 43400 Serdang, Selangor, Malaysia
International Journal of Agricultural Research
Year: 2008 | Volume: 3 | Issue: 4 | Page No.: 261-267







ABSTRACT
The induction of callus from Meristemic Globular Bodies (MGB) of two pineapple cultivars, namely Moris and Josapine, under six concentration levels of auxin NAA and six concentration levels of 2,4-D in Murashige and Skoog solid media, was investigated. 2,4-D auxin treatments failed to induce calli in both cultivars. However, 53.71, 75.19 and 85.93 μM levels of auxin NAA caused calli induction in Moris while levels 32.22, 53.71 and 75.19 μM also induced calli Josapine. The percentage of MGB calli formation increased with increasing time of culturing. At 6 weeks of culturing, 83% of Moris MGB explants formed calli on 85.93 μM NAA, while 50% of Josapine MGB explants formed calli on 53.71 μM NAA. Calli cultures have been an essential tool in the in vitro selection of desirable plants under manipulated conditions and from in vitro mutations via somaclonal variation. More importantly, calli are increasingly used for the application of cellular level genetic modification techniques such as the Agrobacterium-mediated transformation, particle bombardment and protoplast isolation and fusion. In this study, auxin NAA successfully initiated and proliferated calli in Moris and Josapine globular meristemic cultures.
PDF Abstract XML References Citation
How to cite this article
A.E. De Silva, M.A. Kadir, M.A. Aziz and S. Kadzimin, 2008. Callus Induction in Pineapple (Ananas comosus L.) cv. Moris and Josapine. International Journal of Agricultural Research, 3: 261-267.
DOI: 10.3923/ijar.2008.261.267
URL: https://scialert.net/abstract/?doi=ijar.2008.261.267
DOI: 10.3923/ijar.2008.261.267
URL: https://scialert.net/abstract/?doi=ijar.2008.261.267
INTRODUCTION
Moris and Josapine pineapple cultivars are most popular as freshly consumed fruits in Malaysia. The Moris pineapple has the advantage of producing fruits that are high in sugar (13-18 ° Brix), attractive for their golden yellow flesh colour, crispy and spongy in texture, while their spineless leaves (except for the tip) are an advantageous trait as a canning variety (especially during harvesting). However, its average fruit weight is smaller (1.5-1.8 kg) than the Smooth Cayenne (2-4 kg) and tapers in shape. The plant is also highly susceptible to the split peduncle phenomena, which is associated with copper deficiency (Selamat, 2002). The Josapine fruit is also high in sugar content (16-17 ° Brix) with balanced acidity, dry and crispy in texture, cylindrical in shape and has storage duration of up to 5 days under room temperature and two weeks if kept below 10 °C. However, its average fruit weight (1.2-1.5 kg) is smaller than the Moris (Chan and Lee, 1996). Low average fruit weight strongly influences the total crop yield a farmer may harvest. As such both Moris and Josapine are not suitable cultivars for the canning industry. Smooth Cayenne cultivar, is more economical in this regard.
As a result of many years of continued and intense monitoring and screening, the Josapine cultivar was released in 1996 since breeding efforts began in 1984 by the Malaysian Agricultural Research and Development Institute (MARDI). The limited success in producing a marketable variety is due to recombinants that inadvertently inherit defects from their parents (Chan and Lee, 1996; Chan, 1993). An attempt to generate somatic hybrids between fusariosis resistant and susceptible varieties was carried out, as protoplast isolation from the pineapple Perolera cultivar (a fusariosis resistant cultivar), was successful. However, an efficient plant regeneration system from these protoplasts was unsuccessful (Pinho Guedes et al., 1997).
Genetic manipulation allows specific changes to target genes without rearranging the entire genome (Botella et al., 2000). Five transgenic plants possessing herbicide tolerant gene have been successfully regenerated from transformed leaves of a commercial Thai pineapple (Ananas comosus cv. Phuket) via microprojectile-mediated delivery (Sripaoraya et al., 2001). Transformed Smooth Cayenne calli via Agrobacterium tumefaciens regenerated 56 herbicides tolerant transgenic plants (Espinosa et al., 2002) and blackheart resistant transgenic plants via microprojectile and Agrobacterium-mediated delivery (Graham et al., 2000).
Regenerating pineapple plants from callus have been a strong disadvantage for micropropagation with regards to the issue of somaclonal variation. In contrast, somaclonal variation generated valuable traits in various crop species such as sugarcane (Liu and Chen, 1976), sorghum (Maralappanawar et al., 2000), wheat (Guenzi et al., 1992), tomato (Van den Bulk et al., 1990), rice (Yamagishi et al., 1996; Yang et al., 1999) and in garlic (Al-Zahim et al., 1999). In pineapple, calli of a commercial cultivar regenerated one completely white plantlet and another possessed green-streaked leaves (Matthews and Rangan, 1981), the Red Spanish calli regenerated a dwarf plant and one with less spiny leaves (Garcia et al., 2000), while the Smooth Cayenne calli regenerated plants with leaf spininess between 0-14% (Smith et al., 2002). Furthermore, RAPD analysis successfully distinguished the variants possessing spineless leaves characteristics from cultures of normal spined phenotype (Soneji et al., 2002; Feuser et al., 2003).
The importance and usefulness of pineapple callus tissues inevitably serves as a prerequisite to various crop improvement techniques. This study investigated the effects of auxins 2,4-D and NAA at various concentrations on the induction of calli in Moris and Josapine pineapple cultivars.
MATERIALS AND METHODS
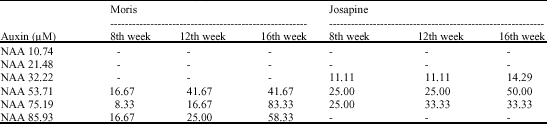
All experiments were carried out at the Cell Biology Laboratory, Faculty of Agriculture Biotechnology, University Putra Malaysia, Malaysia. A 0.5 cm3 of MGB from laboratory maintained pineapple plantlets of both the Josapine and Moris cultivars, were excised and used as explants. The basal medium used in this study was MS media (Murashige and Skoog, 1962) supplemented with 3% (w/v) sucrose and solidified with 0.4% (w/v) Gelrite™. The pH of the medium was adjusted to 5.8 prior to the addition of gelling agent (Matthews and Rangan, 1981). For calli induction, auxins [α-naphthaleneacetic acid (NAA) and 2,4-dichlorophenoxyacetic acid (2,4-D)] treatments were selected based on previous reports on callus proliferation of pineapple (Firoozabady and Moy, 2004; Akbar et al., 2003; Garcia et al., 2000; Benega et al., 1997; De Wald et al., 1988; Matthews and Rangan, 1981, 1979; Apte et al., 1979) and on the advantage that these auxins were relatively more economical to other common auxins such as IAA and IBA (Sigma-Adrich, 2007). The basal medium was modified by adding auxins NAA and 2,4-D, at six different concentrations of NAA (10.74, 21.40, 32.22, 53.71, 75.19 and 85.93 μM) and six different concentrations of 2,4-D (2.26, 4.52, 18.10, 27.15, 45.25 and 54.30 μM). The levels tested for NAA and 2,4-D were selected based on a preliminary study performed in our laboratory (Cell Biology Laboratory, University Putra Malaysia). All the cultures were incubated at 25 ± 2 °C with 16 h light (at an irradiance of 37.5 μmol m-2 sec-1)/8-h light cycle and relative humidity of 50-60% for 16 weeks without subculturing (Matthews and Rangan, 1981; Osei-Kofi and Adachi, 1993). A total of 26 treatments (including controls for both cultivars on MS hormone-free media) were studied. The treatments were replicated 12 times in a Randomized Complete Block Design. Percentage of explants forming calli from the 12 replications, were recorded at the end of the 16th week (Maralappanawar et al., 2000). Data were not statistically analyzed because of the high mortality rate but pictures have been provided to substantiated treatment effect.
RESULTS AND DISCUSSION
In contrast to NAA, all the 2,4-D concentrations (2.26-54.30 μM) failed to induce calli from MGB of both Moris and Josapine, but shoot and root organogenesis were observed similar to controls in (Fig. 1, 2). As a result, the effect of 2,4-D on calli induction was not recorded in Table 1. This observation was consistent with the work of Matthews and Rangan (1981) who also found that 2,4-D could not induce calli from in vitro lateral buds of a commercial pineapple cultivar but rather favoured calli proliferation. However, these workers and others observed that NAA caused calli induction and proliferation in various pineapple varieties such as the Moris (De Silva et al., 2006), the Serrana Smooth Cayenne and Perolera hybrid embryos (Benega et al., 1997) and a commercial cultivar (Matthews and Rangan, 1981). Furthermore, although various studies report that pineapple calli was induced via various combinations of plant growth regulators (Firoozabady and Moy, 2004; Akbar et al., 2003; Garcia et al., 2000; Benega et al., 1997; DeWald et al., 1988; Matthews and Rangan, 1981, 1979; Apte et al., 1979), it is apparent that the common auxin inducer is NAA, rather than 2,4-D (Garcia et al., 2000). In this study, the independent effect of these two auxins on the calli proliferation of the local Moris and Josapine cultivars, was clearly differentiated.
| Table 1: | Calli induction from MGB of the Moris and Josapine cultivar with different concentrations of NAA and 2,4-D without subculture for 16 weeks (%) |
 | |
| (%): No. of responding explants/12 explants (replicates), (-): Calli formation was not observed | |
 | |
| Fig. 1: | Control for Moris MGB, at the sixteenth week |
| |
| Fig. 2: | Control for Josapine MGB, at the sixteenth week |
Recently, Campanoni and Nick (2005) tested the effects of 2,4-D and NAA on the response of tobacco pith tissues. Contrary to pineapple, 2,4-D was a potent inducer of cell division in tobacco even at high concentrations, while NAA caused cell elongation at low concentrations and only caused cell division at high levels. It was explained that there may be two different auxin receptors specifically binding to each of the auxins regulated by their degree of affinity which subsequently launched two different signaling pathways resulting in different physiological functions [one such example is the NAA auxin binding protein (ABP1)]. This suggests a different auxin receptor or signaling pathway with regards to the induction of cell division in pineapple, from that of the tobacco model system.
In another study conducted on tobacco, Chen (2001) discussed that there were two different auxin receptors that bind to NAA (dual-auxin pathway). One auxin receptor, identified as ABP1 was reported to possess a high-affinity binding site which causes cell elongation under low auxin concentration levels. The other receptor (unidentified) is believed to possess a low-affinity binding site which causes cell division under high auxin concentration levels. In our study, calli induction was observed in Moris at 57.31, 75.19 and 85.93 μM NAA but not at levels lower than 57.31 μM NAA (Table 1, Fig. 3). Calli induction was observed in Josapine at 32.22, 57.31 and 75.19 μM NAA but not lower than 32.22 μM NAA (Table 1, Fig. 4). The calli induction response of MGB to their respective optimal NAA levels is therefore believed to be due to the presence of two auxin binding proteins, each binding protein responding to different auxin concentrations which controls cell elongation and division independently. Nonetheless, it is essential that future studies identify and characterize auxin receptors in pineapple, as this would elucidate the developmental pathway that differs from that of the tobacco model pathway.
Calli induction was not observed in Josapine when NAA level increased to 85.93 μM. Furthermore, the percentage of responding explants decreased with increasing NAA levels for both Moris and Josapine. These explants did not show any signs of further growth or development (arrested growth), but rather gradually became brown (necrotic). According to Campononi and Nick (2005), the Virginia Bright Italia (VBI-O) tobacco cells that had optimal cell division activity at 10 μM auxins(5.5 μM NAA and 4.5 μM 2,4-D), demonstrated a sharp decrease in cell division when auxin concentration deviated from the optimal concentration. This was not due to decrease in cell viability but rather the morphology. Cells under 1 μM of auxin were more elongated and the divided cells were subject to precocious disintegration. Cells under 100 μM were not pluricellular (many cells arranged in a file) but bicellular (two cells in a file). The bicellular cells had very small basal cell and an extremely elongated polar cell. These cells also contain extraordinarily bundled cytoplasmic strands. Although this report may suggest a similar degenerative response for the pineapple explants in this study, future studies are essential to elucidate the effect of high auxin concentrations on the molecular, biochemical and morphological condition of pineapple cells.
 | |
| Fig. 3: | Calli induction from Moris MGB under different NAA levels, at the sixteenth week (2.5 cm diameter vials) |
 | |
| Fig. 4: | Calli induction from Josapine MGB under different NAA levels, at the 16th week |
It was also observed that Josapine responded to a lower NAA concentration at 32.22 μM, than Moris at 53.71 μM. Although their difference in sensitivity to auxin could be generally explained by their genetic differences, we suggest three possible reasons based on the tobacco model system (Campanoni and Nick, 2005; Chen, 2001): (a) Moris and Josapine possess the same auxin receptors but in different quantities; (b) Josapine possesses the same auxin receptor but an alternative pathway was activated; (c) Josapine possesses an additional auxin receptor which has high affinity to NAA, but low affinity to another auxin (other than 2,4-D and NAA).
The percentage of MGB forming calli increased with time for both varieties (Table 1). By the end of the 16th week, 83.33% Moris MGB formed calli at 14 mg L-1, while 50% of Josapine MGB formed calli at 10 mg L-1. Cultures that were maintained beyond the 16th week gradually became necrotic.
CONCLUSION
2,4-D inhibited calli induction in both Moris and Josapine cultivar. NAA concentrations of 53.71, 75.19 and 85.93 μM NAA cause calli induction in Moris while for Josapine the concentrations that caused calli induction were 32.22, 53.71 and 85.93 μM. The percentage of MGB calli formation increased with increasing time of culturing for both Moris and Josapine. By the 16th week of culture, 83.33% of Moris MGBs formed calli on 75.79 μM NAA and 50% of Josapine MGBs formed calli on 53.71 μM. We also emphasized that NAA is generally the potent inducer of cell division or proliferation for pineapple rather than 2,4-D as observed in the tobacco model system. Future in vitro culture investigations on the comparison and elucidation of the molecular, biochemical and physiological characteristics between pineapple and the tobacco model system is therefore expected to provide useful and in depth contributions to the understanding of plant cell division and differentiation.
REFERENCES
- Al-Zahim, M.A., B.V. Ford-Lloyd and H.J. Newbury, 1999. Detection of somaclonal variation in garlic (Allium sativum L.) using RAPD and cytological analysis. Plant Cell Rep., 18: 473-477.
CrossRefDirect Link - Apte, P.V., G.S. Kaklij and M.R. Heble, 1979. Proteolytic enzymes (bromelains) in tissue cultures of Ananas sativus (pineapple). Plant Sci. Lett., 14: 57-62.
CrossRefDirect Link - Benega, R., M. Isidron, E. Arias, A. Cisneros, J. Martinez, I. Torres, M. Hidalgo and C.G. Borroto, 1997. In vitro germination and callus formation in pineapple hybrid seeds (Ananas comosus (L.) Merr.). Acta Hortic., 425: 243-246.
CrossRefDirect Link - Botella, J.R., A.S. Cavallaro and C.I. Cazzonelli, 2000. Towards the Production of Transgenic Pineapple to Control Flowering and Ripening. In: Proceedings of the 3rd International Pineapple Symposium, Subhadrabandhu, S. and P. Chairidchai (Eds.). Acta Hortic., Pattaya, Thailand, pp: 115-122.
Direct Link - Campanoni, P. and P. Nick, 2005. Auxin-dependent cell division and cell elongation. 1-Naphthaleneacetic acid and 2,4-dichlorophenoxyacetic acid activate different pathways. Plant Physiol., 137: 939-948.
CrossRefDirect Link - Chen, J.G., 2001. Dual auxin signaling pathways control cell elongation and division. J. Plant Growth Regul., 20: 255-264.
CrossRefDirect Link - De Silva, A.E., M.A. Kadir, M.A. Aziz and S. Kadzimin, 2006. Proliferation potential of 18-month-old callus of Ananas comosus L. cv. Moris. Sci. World J., 6: 169-175-175.
Direct Link - DeWald, M.G., G.A. Moore, W.B. Sherman and M.H. Evans, 1988. Production of pineapple plants in vitro. Plant Cell Rept., 7: 535-537.
CrossRefDirect Link - Espinosa, P., J. Lorenzo, A. Iglesias, L. Yabor and E. Menendez et al., 2002. Production of pineapple transgenic plants assisted by temporary immersion bioreactors. Plant Cell Rep., 21: 136-140.
CrossRef - Feuser, S., K. Meler, M. Daquinta, M.P. Guerra and R.O. Nodari, 2003. Genotypic fidelity of micropropagated pineapple (Ananas comosus) plantlets assessed by isozyme and RAPD markers. Plant Cell Tiss. Organ Cult., 72: 221-227.
Direct Link - Firoozabady, E. and Y. Moy, 2004. Regeneration of pineapple plants via somatic embryogenesis and organogenesis. In vitro Cell Dev. Biol. Plant, 40: 67-74.
Direct Link - Graham, M., L. Ko, V. Hardy, S. Robinson and B. Sawyer et al., 2000. The development of blackheart resistant pineapple through genetic gngineering. Acta Hortic. (ISHS), 529: 133-138.
Direct Link - Mathews, V.H. and T.S. Rangan, 1979. Multiple plantlets in lateral bud and leaf explant in vitro culture of pineapple. Sci. Hortic., 11: 319-328.
CrossRef - Liu, M.C. and W.H. Chen, 1976. Tissue and cell culture as aids to sugarcane breeding. I. Creation of genetic variation through callus culture. Euphytica, 25: 393-403.
CrossRefDirect Link - Maralappanawar, M.S., M.S. Kuruvinashetti and C.C. Harti, 2000. Regeneration, establishment and evaluation of somaclones in Sorghum bicolor (L.) Moench. Euphytica, 115: 173-180.
Direct Link - Mathews, V.H. and T.S. Rangan, 1979. Multiple plantlets in lateral bud and leaf explant in vitro culture of pineapple. Sci. Hortic., 11: 319-328.
CrossRef - Mathews, V.H. and T.S. Rangan, 1981. Growth and regeneration of plantlets in callus cultures of pineapple. Scient. Hortic., 14: 227-234.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Pinho Guedes, N.M., J. Maria, L. Zambolim and J.A. Ventura, 1997. Protoplast isolation of Ananas comosus (L.) Merr. cv 'Perolera'. Acta Hortic., 425: 259-266.
CrossRefDirect Link - Smith, M.K., H.L. Ko, S.D. Hamill and G.M. Sanewski, 2002. Pineapple transformation: Mangging somaclonal variation. Acta Hortic., 575: 69-74.
Direct Link - Soneji, J.R., P.S. Rao and M. Mhatre, 2002. Suitability of RAPD for analyzing spined and spineless variant regenerants of pineapple (Ananas comosus L., Merr.). Plant Mol. Biol. Rep., 20: 307-307.
CrossRefDirect Link - Sripaoraya, S., R. Marchant, J.B. Power and M.R. Davey, 2001. Herbicide-tolerant transgenic pineapple (Ananas comosus) Produced by microprojectile bombardment. Ann. Bot., 88: 597-603.
Direct Link - Bulk., R.W., H.J.M. Loffler, W.H. Lindhout and M. Koornneef, 1990. Somaclonal variation in tomato: Effect of explant source and a comparison with chemical mutagenesis. Theor. Applied Genet., 80: 817-825.
CrossRef - Yamagishi, M., M. Otani and T. Shimada, 1996. A comparison of somaclonal variation in rice plants derived and not derived from protoplasts. Plant Breed., 115: 289-294.
CrossRefDirect Link - Yang, H., Y. Tabei, H. Kamada, T. Kayano and F. Takaiwa, 1999. Detection of somaclonal variation in cultured rice cells using digoxigenin-based random amplified polymorphic DNA. Plant Cell Rep., 18: 520-526.
CrossRefDirect Link