
Hou-Yang Kang
Triticeae Research Institute, Sichuan Agricultural University, Dujiangyan City 611830, Sichuan, People`s Republic of China
Yi Wang
Triticeae Research Institute, Sichuan Agricultural University, Dujiangyan City 611830, Sichuan, People`s Republic of China
Hong-Jun Yuan
Triticeae Research Institute, Sichuan Agricultural University, Dujiangyan City 611830, Sichuan, People`s Republic of China
Yun Jiang
Triticeae Research Institute, Sichuan Agricultural University, Dujiangyan City 611830, Sichuan, People`s Republic of China
Yong-Hong Zhou
Triticeae Research Institute, Sichuan Agricultural University, Dujiangyan City 611830, Sichuan, People`s Republic of China
International Journal of Agricultural Research
Year: 2008 | Volume: 3 | Issue: 4 | Page No.: 252-260







ABSTRACT
By colchicine treatment of the hybrid plants between Triticum polonicum L. from Xinjiang in China and Aegilops tauschii Cosson. from Middle East, a new synthetic hexaploid wheat (SHW-DPW) was artificially obtained for the fist time. The average seed set of the intergeneric hybrids was 9.71% (8.27-15.31%) with colchicines treatment. The morphology of the SHW-DPW plants was comparable to that of the primary F1 plants and the SHW-DPW plants appeared more robust. All the SHW-DPW plants were uniform in morphology and had some obvious traits inherited from T. polonicum and Ae. tauschii. Especially, the glumes were very soft and the rachis internodia was short, which was different from other synthetic hexaploid wheat. Their spikes were morphologically similar to those of Triticum petropavlovskyi Udacz. et Miguch. The SHW-DPW plants were fertile, with 58.95% selfed seed set for the euhexaploids and 45.63% for the aneuploids, respectively. The meiosis of the SHW-DPW plants was quite regular, which showed a pairing configuration of 0.43 univalents, 20.77 bivalents and 0.01 trivalents. The rate of chromosome association of the aneuploids was comparatively lower than that in the euploid individuals. The potential utilization for wheat improvement and study for the origin of T. petropavlovskyi are discussed in the present study.
PDF Abstract XML References Citation
How to cite this article
Hou-Yang Kang, Yi Wang, Hong-Jun Yuan, Yun Jiang and Yong-Hong Zhou, 2008. A New Synthesized 6x-Wheats, Derived from Dwarfing Polish Wheat (Triticum polonicum L.) and Aegilops tauschii Cosson. International Journal of Agricultural Research, 3: 252-260.
DOI: 10.3923/ijar.2008.252.260
URL: https://scialert.net/abstract/?doi=ijar.2008.252.260
DOI: 10.3923/ijar.2008.252.260
URL: https://scialert.net/abstract/?doi=ijar.2008.252.260
INTRODUCTION
Allopolyploidy has played a major role in the evolution of crop plants. One of the best-known allopolyploid complexes is Triticum-Aegilops in Triticeae (Poaceae). Among the roughly 30 species of Triticum and Aegilops, 75% of them are natural allotetraploids (2n = 4x = 28) or allohexaploids (2n = 4x = 42) (Sakamoto, 1973; Yen et al., 2005). Bread wheat (Triticum aestivum L., 2n = 6x = 42, AABBDD), one of the most remarkable allopolyploids, also called common wheat, has no direct hexaploid wild progenitor (Morris and Sears, 1967). It possesses three sets of homologous chromosomes, whose A genome stems from Triticum urartu Thum., the D genome stems from the wild diploid Aegilops tauschii Cosson. and the B genome donor is probably Aegilops speltoides L. or its related species in the section Sitopsis of Aegilops (Kihara, 1944; Heun et al., 1997; Feldman, 2001). Bread wheat has undergone two hybridizations and polyploidizations during its origin and evolution. Tetraploid Triticum wheat (Triticum turgidum L., 2n = 4x = 28, AABB) was formed in the first intercrossing between T. urartu and Ae. speltoides or its related species, followed by chromosome doubling. Then bread wheat was formed by the second polyploidization after the intercrossing between the cultivated T. turgidum (maternal) and Ae. tauschii (paternal) in ca. 10000 years ago (Feldman, 2001; Huang et al., 2002).
Since only a few accessions of tetraploid wheat or Ae. tauschii genotypes were involved in the evolutionary origin of bread wheat, the genetic diversity of bread wheat were largely decreased in comparison with that of its donor species (Miller, 1987; Feldman, 2001). Numerous genetic variations in the ancestral tetraploid wheat or Ae. tauschii were not represented at the hexaploid level due to the evolution bottleneck (Dhaliwal et al., 1993; Ayal et al., 2005; Warburton et al., 2006). Close evolutionary relationship and extensive genetic diversity for desirable traits of tetraploid wheat and Ae. tauschii were especially interesting for bread wheat improvement. The genetic diversity may be introgressed into bread wheat by the bridge of synthetic hexaploid or amphidiploids derived from artificial synthesis of hexaploid wheat (tetraploid wheat x Ae. tauschii), which was a manner analogous to the evolution of hexaploid wheat. Ever since the reports by McFadden and Sears (1944) and Kihara and Lilienfeld (1949) on the artificial synthesis of hexaploid wheat between existing T. turgidum with Ae. tauschii, many synthetic hexaploids have been obtained from the hybridizations between Ae. tauschii and the tetraploid wheat, such as T. durum, T. dicoccon, T. carthlicum, T. paleocolchicum, T. dicoccoides (Tanaka, 1961; Lange and Jochemsen, 1992; Mujeeb-Kazi et al., 1996; Lage et al., 2003; Warburton et al., 2006). Furthermore, there is no report of the successful synthesis of the amphidiploid between T. polonicum and Ae. tauschii.
Dwarfing polish wheat (Triticum polonicum L., 2n = 4x = 28, AABB, DPW) is a tertraploid species that was collected from Tulufan, Xingjiang, China. It is the only one T. polonicum with the trait of dwarfing in China, which was characterized as dwarfing, more tillers, more length of spike, more spikelets per spike and length of glume (Yang et al., 2001; Liu et al., 2002). In 2005, we successfully carried out the intergeneric hybridization between T. polonicum (DPW) and Ae. tauschii (AS60) and obtained the F1 hybrids without the use of embryo rescue. The production, morphology, meiotic behavior and fertility of a new synthetic hexaploid wheat (2n = 6x = 42, AABBDD, SHW-DPW) derived from T. polonicum (DPW) x Ae. tauschii (AS60) by chromosome doubling are described in the present study.
MATERIALS AND METHODS
Plant Materials
Dwarfing polish wheat (Triticum polonicum L., 2n = 4x = 28, AABB; DPW) was collected in Tulufan, Xingjiang, China by Prof. C. Yen and J.L. Yang. Aegilops tauschii Cosson. (AS60) was originated from Middle East. Triticum petropavlovskyi Udacz. et Miguch (2n = 6x = 42, AABBDD) (AS356 and AS358) was originated from Xingjiang, China. All the materials were deposited in Triticeae Research Institute, Sichuan Agricultural University, Sichuan, China. This research was conducted at the Triticeae Research Institute of Sichuan Agricultural University, China in 2005-2007.
The Production of Synthetic Hexaploid Wheat
To produce artificially synthetic wheat, T. polonicum (DPW) as female was crossed to Ae. tauschii (AS60) and F1 seeds were obtained in 2005. The emasculation and pollination techniques were carried out as described by Liu et al. (2002). No embryo rescue techniques or gibberellic acid solution were applied to the embryos after pollination. The F1 seeds were germinated in petri dishes and the seedlings were then planted in the field. To double the chromosome, colchicine treatment was carried out as per the following procedure. When the F1 seedlings grew to the six-seven leaf stage, they were removed from soil and the roots were washed and cut to a length of 25 mm. These were then placed in 30 mm diameter glass tubes and a solution of 0.05% colchicine was added such that the crown was submerged to a depth of 50 mm. The plants (in tubes) were then placed at 20 °C for 22 h. Then they were removed from the tubes, rinsed with water and returned to the tubes containing only water. The water in the tubes was changed every 20 min for 8 h to ensure complete removal of residual colchicines. After the leaves were trimmed to approximately half their original length to reduce transpiration, the plants were transplanted into field. To maintain a humid environment, the plants were covered by clear plastic covers. After seven days, the plastic covers were removed. Before flowering, spikes on well developed tillers of the treated plants were bagged to ensure self-pollination. At maturity, seeds (F2) were harvested from all the treated plants. The F2 plants were bagged and selfed and the F3 seeds were obtained. Crossability percentages and seedset were estimated.
Morphology and Meiosis Analysis
Eight morphological characters including plant height, tillering, length of spike, number of spikelets per spike, length of glume, length of flag leaf, length of internodes of spike and length of awn of the amphidiploid plants (SHW-DPW) obtained from the hybrids T. polonicum (DPW) x Ae. tauschii (AS60) and the parental species were measured and calculated.
For mitotic studies, the root tips (1-3 cm long) were cut and pretreated in ice water at 0-4 °C for 20-24 h and fixed in Carnoy`s fixative I (a 3:1 mixture of 95% ethanol and glacial acetic acid) overnight. The root tips were hydrolyzed in 1 M HCl at 60 °C for 10 min, stained following the Feulgen procedure. Somatic chromosomes were counted. Analyses of meiosis behavior in the synthetic hexaploid wheat (SHW-DPW) were performed by the conventional aceto-carmine squash method. Stages of meiosis were determined in aceto-carmine squashes of one of three anthers per flower. If appropriate stages were present, the remaining two anthers were fixed in a mixture of absolute ethanol, chloroform and acetic acid (6:3:1, v/v) and kept in a refrigerator for 24 h, then stored under refrigeration (4 °C) in 70% alcohol until use. Cytological observations and documentation were made with an Olympus BX-51 microscope coupled with a Photometrics SenSys CCD camera.
Pollen fertility was measured for pollen grains sampled from mature anthers at flowering. The grains were stained in a 2% aceto-carmine solution with glycerin.
RESULTS
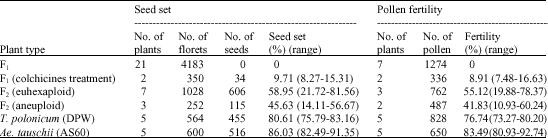
Without using embryo rescue technique, F1 hybrid seeds between T. polonicum (DPW) and Ae. tauschii (AS60) were obtained and the crossability was 1.67%. The F1 seeds of the T. polonicum (DPW) x Ae. tauschii were germinated and 24 plants were obtained. The morphological characteristics of these F1 plants were similar to those of bread wheat. The F1 hybrid plants have 2n = 21 chromosomes. Three F1 hybrid plants were used for colchicines treatment, one was killed and the other two plants showed various degrees of regrowth. After vernalization, the two plants were grown to flowering and formed four and eight spikes per plant, respectively. A total number of 11 and 23 seeds were obtained and the rates of selfed seed set were 15.31 and 8.27%, respectively (Table 1). All the spikes of the other 21 F1 plants were bagged to avoid unexpected pollination and no seeds were obtained under self-pollination.
| Table 1: | The seedset and pollen fertility of F1 and F2 hybrid of T. polonicum (DPW) with Ae. tauschii (AS60) |
 | |
| Table 2: | Comparison of morphological characters among T. polonicum (DPW), Ae. tauschii (AS60) and synthetic hexaploid wheat (SHW-DP) |
 | |
 | |
| Fig. 1: | Spikes morphology of T. polonicum (DPW), Ae. tauschii (AS60) and the synthetic hexaploid wheat (SHW-DPW) (A) Spike: a. T. polonicum (DPW); b. SHW-DPW; c. Ae. tauschii (AS60) (B) Spikelets: a. T. polonicum (DPW); b. SHW-DPW; c. Ae. tauschii (AS60) |
Ten well-developed F2 seeds were selected and germinated. All of the seeds were germinated and the plants grew vigorously. The morphology of the SHW-DPW plants was similar to that of the F1 plants. But the SHW-DPW plants appeared more robust, such as plant height, number of spikelets per spike, length of spikes and length of flag leaf. As expected, all the SHW-DPW plants were uniform in morphology and had some obvious traits inherited from T. polonicum (DPW), such as number of tillers, number of spikelets per spike and length of awn and internodes of spike, while non-waxiness, tough rachis and hairy auricles resembled Ae. tauschii (Table 2, Fig. 1). Compared with T. polonicum (DPW), the SHW-DPW plants had short glumes and long length of spikes. It is remarkable that the SHW-DPW plants have soft glume and short rachis internodes. Furthermore, the spike morphology of SHW-DPW was very similar to T. petropavlovskyi (Fig. 2). While the powdery mildew disease and budworm occurred heavily on T. polonicum (DPW), the SHW-DPW plants were immune. The results of the pollen fertility of F1 hybrids, synthetic hexaploid wheat (SHW-DPW) and their parents are listed in Table 1. Spikes on well developed tillers of SHW-DPW plants were bagged to avoid unexpected pollination and the average selfed seedset was 58.95% for the euhexaploids and 45.63% for the aneuploids (Table 1).
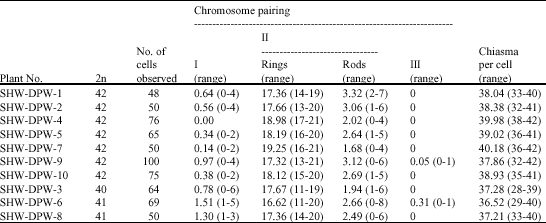
| Table 3: | The configuration of meiotic MI in synthetic hexaploid wheat (SHW-DPW) obtained from the hybrids of T. polonicum (DPW) x Ae. tauschii (AS60) |
 | |
 | |
| Fig. 2: | Spikes morphology of the synthetic hexaploid wheat (SHW-DPW) and T. petropavlovskyi (A) Spike: a. T. petropavlovskyi (AS356); b. SHW-DPW; c. T. petropavlovskyi (AS358) (B) Spikelets: a. T. petropavlovskyi (AS356); b. SHW-DPW |
Of the ten synthetic hexaploid wheat (SHW-DPW) plants, seven were euhexaploids with 42 chromosomes, the other three plants were aneuploids with 40 (one plants) and 41 (two plants) chromosomes, respectively (Fig. 3a, b). The results of the chromosome pairing in meiotic metaphase I (MI) of the SHW-DPW plants are summarized in Table 3. The chromosome pairing of the euploid plants was quite regular with average 0.43 univalents, 20.77 bivalents and 0.01 trivalents per cell (Table 3, Fig. 3c, d). Little variation was observed among the plants. Plant SHW-DPW-4 showed the highest rate of chromosome association, with 21 bivalents per cell, while plant SHW-DPW-9 was characterized by the lowest rate of chromosome association (20.44). At anaphase I, lagging chromosome could be observed in some cells analyzed and some of them undergo precocious
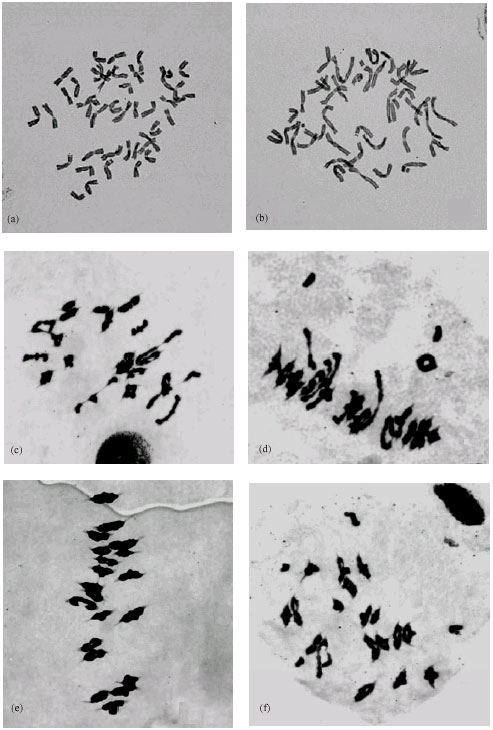 | |
| Fig. 3: | Chromosome observation for the synthetic hexaploid wheat (SHW-DPW) (a) The somatic chromosomes of SHW-DPW with 2n = 42; (b) The somatic chromosomes of SHW-DPW with 2n = 41; (c-f) The meiotic chromosomes of SHW-DPW: C. 2n = 42 = 15 II (ring) + 6 II (rod); D. 2n = 42 = 2 I + 15 II (ring) + 5 II (rod); E. 2n = 40 = 20 II (ring); F. 2n = 41 = 4 I + 17 II (ring) + 1 III |
disjunction. In the aneuploid individuals, the rate of chromosome association was comparatively lower than that in euploid individuals, which was ascribed to the increment of univalents (Table 3, Fig. 3e, f).
DISCUSSION
Production of Synthetic Hexaploid Wheat (SHW-DPW) Between T. polonicum and Ae. tauschii
Ae. tauschii (AS60) has a unique genotype which facilitate hybrid seed development and viability and which is the important donor material in cross with tetraploid wheat and bread wheat (Liu et al., 2002). After a few years` efforts, we have successfully obtained the F1 hybrids between T. polonicum (DPW) and Ae. tauschii (AS60) without using embryo rescue for the first time in 2005. The seed set of the cross was 1.67%. In the present study, the synthetic hexaploid wheat (SHW-DPW) between T. polonicum (DPW) and Ae. tauschii (AS60) were obtained with colchicine treatment for the first time. The average seed set of the intergeneric hybrids was 9.71% (8.27-15.31%), which was much lower than 44% (on the basis of plants) observed for the wheat haploids-double (De Buyser and Henry, 1986). The selfed seed set of SHW-DPW were 58.95% for the euhexaploids and 45.63% for the aneuploids, respectively, which were much lower than the rates (87.40-100%) of synthetic hexaploid wheat between Ae. tauschii and the tetraploid wheat, T. turgidum, T. durum, T. carthlicum, T. paleocolchicum, T. dicoccon and T. dicoccoides (McFadden and Sears, 1944; Lange and Jochemsen, 1992; Warburton et al., 2006).
In this study, we successfully obtained the F1 hybrids and synthetic hexaploid wheat (SHW-DPW) by a lot of crosses between T. polonicum (DPW) and Ae. tauschii (AS60). Therefore, the genotypes of different the tetraploid wheat and Ae. tauschii, the process of chromosome doubling with colchicines treatment and the field management are very important in hybridization and chromosome doubling of F1 hybrids between the tetraploid wheat and Ae. tauschii.
Morphology and Cytology Assay of Synthetic Hexaploid Wheat (SHW-DPW)
Morphologically, the traits of tillers, spikelets per spike and length of awn and internodes of spike in SHW-DPW plants were similar to those in T. polonicum (DPW), while non-waxiness, hairy auricles and tough rachis in SHW-DPW plants were similar to those in Ae. tauschii. All the hybrid plants were resistant to powdery mildew and budworm. In the previous studies, the synthetic hexaploid wheats between Ae. tauschii and T. turgidum, T. durum, T. dicoccon and T. dicoccoides were reported to have a relatively lax spikes, which resulted from the long rachis internodes (Lange and Jochemsen, 1992; Lage et al., 2003; Yoshihiro and Shuhei, 2004). In the present study, the SHW-DPW plants have a comparatively short rachis internode, which in general was similar to that in the F1 hybrids of T. polonicum (DPW) x Ae. tauschii (AS60) and T. petropavlovskyi. Compared with the other synthetic hexaploid wheat, SHW-DPW has a longer and softer glume.
Chromosome association at metaphase I of meiosis of SHW-DPW was quite regular, with an average of 0.43 univalents, 20.77 bivalents and 0.01 trivalents per cell. The trivalent configurations only appeared in two SHW-DPW plants. Three out of 10 of SHW-DPW plants were aneuploids. The differences of chromosome pairing among the plants were small and the average values of chromosome association were similar to those of synthetic hexaploid wheat between T. turgidum, T. durum, T. carthlicum, T. dicoccon and T. dicoccoides and Ae. tauschii (Tanaka, 1961; Lange and Jochemsen, 1992; Warburton et al., 2006). The chromosome behaviour suggests that the SHW-DPW was cytologically stable and regular.
The Potential Value of Synthetic Hexaploid Wheat (SHW-DPW) for Studying the Origin of Triticum petropavlovskyi
The origin of Triticum petropavlovskyi Udacz. et Migusch. (2n = 6x = 42, AABBDD) has remained an issue (Chen et al., 1985; Yang et al., 1992; Akond et al., 2007). Based on previous studies, there were three hypotheses: (1) a single mutation in T. aestivum; (2) independent allopolyploidization between T. polonicum and Ae. tauschii; (3) introgression from T. polonicum to T. aestivum. In this study, we successfully obtained the synthetic hexaploid wheat (SHW-DPW) between T. polonicum (DPW) and Ae. tauschii through chromosome doubling. The spike morphology of SHW-DPW was similar to T. petropavlovskyi, especially the long and soft glume (Fig. 2). Meanwhile, the hybrids of bread wheat x T. polonicum and a set of T. aestivum-T. polonicum introgression lines with long glume have also been obtained. These materials will be significant to the study of the origin of T. petropavlovskyi. Further studies are in progress in our laboratory.
Implications of Synthetic Hexaploid Wheat (SHW-DPW) for the Wheat Breeding
It is advantageous for wheat improvement through the production of synthetic hexaploid wheat. Since it allows not only the Ae. tauschii resistance to be exploited but also incorporates the genetic diversity of the A and B genomes of the respective tetraploid wheat. In the present study, a new synthetic hexaploid wheat (SHW-DPW) between T. polonicum (DPW) and Ae. tauschii (AS60) were obtained successfully. As expected, the SHW-DPW plants inherited the traits of T. polonicum (DPW), such as dwarfing (about 75 cm), more tillers (about 10), more length of spike (about 19 cm) and more spikelets per spike (about 23) and the characters of resistance to powdery mildew and budworm of Ae. tauschii were also expressed. Meanwhile, we successfully obtained a set of hybrids and many derivatives between SHW-DPW and bread wheat. These hybrids, SHW-DPW and the derivatives could be important germplasms for the genetic improvement of bread wheat.
ACKNOWLEDGMENTS
The authors thank Profs. You-Liang Zheng, Deng-Cai Liu and Xiu-Jin Lan of Triticeae Research Institute, Sichuan Agricultural University for several useful suggestions. This work was supported by the National Natural Science Foundation of China (No. 30470135, 30670150), the Program for Changjiang Scholars and Innovative Research Teams in University of China (IRT0453) and the Education Bureau and Science and Technology Bureau of Sichuan Province, China.
REFERENCES
- Akond, A.S.M.G.M., W. Nobuyoshi and F. Yoshihiko, 2007. Exploration of genetic diversity among Xinjiang Triticum and Triticum polonicum by AFLP markers. J. Applied Genet., 48: 25-33.
PubMedDirect Link - De Buyser, J. and Y. Henry, 1986. Wheat: Production of haploids, performance of doubled haploids and yield trials. Biotechnol. Agric. For., 2: 73-88.
CrossRefDirect Link - Heun, M., R. Schafer-Pregi, D. Klawan, R. Castagna, M. Accerbi, B. Borghi and F. Salamini, 1997. Site of the einkorn wheat domestication identified by DNA fingerprinting. Science, 278: 1312-1314.
CrossRefDirect Link - Huang, S., A. Sirikhachornkit, X. Su, J. Faris, B.S. Gill, B. Haselkorn and P. Gomicki, 2002. Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of the Triticum/Aegilops complex and the evolutionary history of wheat. Proc. Natl. Acad. Sci. USA., 99: 8133-8138.
PubMedDirect Link - Lage, J., M.L. Warburton, J. Crossa, B. Skovmand and S.B. Andersen, 2003. Assessment of genetic diversity in synthetic hexaploid wheats and their Triticum dicoccon and Aegilops tauschii parents using AFLPs and agronomic traits. Euphtytica, 134: 305-317.
CrossRef - Lange, W. and G. Jochemsen, 1992. Use of the gene pools of Triticum turgidum sp. dicoccoides and Aegilops squarrosa for the breeding of common wheat (T. aestivum) through chromosome-doubled hybrids. I. Two strategies for the production of the amphiploids. Euphytica, 59: 197-212.
CrossRef - Liu, G.X., Y.H. Zhou and Y.L. Zheng, 2002. Morphological and cytological studies of dwarfing polish wheat (Triticum turgidum concv. polonicum) from Xinjiang China. J. Sichuan Agric. Univ., 20: 189-193.
Direct Link - Mujeeb-Kazi, A., V. Rosas and S. Roldan, 1996. Conservation of the genetic variation of Triticum tauschii (Coss.) Schmalh. (Aegilops squarrosa auct. non L.) in synthetic hexaploid wheats (T. turgidum L. s. lat. xT. tauschii; 2n = 6x = 42, AABBDD) and its potential utilization for wheat improvement. Genet. Resour. Crop Evol., 43: 129-134.
CrossRefDirect Link - Warburton, M.L., J. Crossa, J. Franco, M. Kazi, R. Trethowan, S. Rajaram, W. Pfeiffer, P. Zhang, S. Dreisigacker and V.M. Ginkel, 2006. Bringing wild relatives back into the family: Recovering genetic diversity in CIMMYT improved wheat germplasm. Euphytica, 149: 289-301.
CrossRef - Yen, C., J.L. Yang and Y. Yen, 2005. Hitoshi Kihara, Áskell Löve and the modern generic concept of the genera in the tribe Triticeae (Poaceae). Acta Phytotax. Sin., 43: 82-93.
Direct Link - Yoshihiro, M. and N. Shuhei, 2004. Durum wheat as a candidate for the unknown female progenitor of bread wheat: An empirical study with a highly fertile F1 hybrid with Aegilops tauschii Cosson. Theor. Applied Genet., 109: 1710-1717.
CrossRefDirect Link