
J. Masaka
Department of Land and Water Resources Management, Faculty of Natural Resources Management and Agriculture, Midlands State University, Private Bag 9055, Gweru, Zimbabwe
N. Khumbula
Department of Land and Water Resources Management, Faculty of Natural Resources Management and Agriculture, Midlands State University, Private Bag 9055, Gweru, Zimbabwe
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 7 | Page No.: 581-589







ABSTRACT
In nursery coffee seedling production, farmers often subject potted soil media to casual compaction without considering the effects of such practices on the physical properties of growing media, which significantly influence seedling emergence and biomass characteristics. A 32-week study was carried out at Chipinge Coffee Research Station (20°13’ S; 32°36’ E, elev. >1500 m above sea level), Zimbabwe. The main objective of the study was to determine the effect of different compaction levels in the nursery on the emergence and biometric characteristics of coffee. The first seedling emergence counts in the 5th week after sowing showed that seemingly insignificant bulk density increases of 5.7% (6.6 g cm-3) and 37.5% (43.28 g cm-3) in T2 and T3 potted soils above no-compaction levels triggered uncorrespondingly massive reductions of 75 and 90% in coffee seed germination, respectively. The no-compaction seedlings were 30.33 and 56.27 mm taller than those in the semi and full soil compaction pots respectively in the 21st week after sowing. No-compactioncoffee seedlings recorded 0.689-0.911 mm thicker stems than those observed in semi and full compaction pots. Semi compaction simulations reduced root mass per plant by a modest 1.88 g while full compaction in potted soil media dwarfed root biomass by a massive 5.34 g.
PDF Abstract XML References
How to cite this article
J. Masaka and N. Khumbula, 2007. The Effect of Soil Compaction Levels on Germination and Biometric Characteristics of Coffee (Coffee arabica) Seedlings in the Nursery. International Journal of Agricultural Research, 2: 581-589.
DOI: 10.3923/ijar.2007.581.589
URL: https://scialert.net/abstract/?doi=ijar.2007.581.589
DOI: 10.3923/ijar.2007.581.589
URL: https://scialert.net/abstract/?doi=ijar.2007.581.589
INTRODUCTION
Coffee (Coffee arabica) is a tropical perennial crop that is not indigenous to Zimbabwe, having originated from the Highlands of Ethiopia. It is a cash crop of value competing well on the world commodity trade (in the top five) and has thus shaped land uses, economies and societies in more than eighty countries the world over and can be produced both at large scale commercial and smallholder levels (Wilson, 1991).
Coffee seeds are planted in polythene or clay pots filled with disease free soils. The process of filling the pots with soil, in most cases, casual with little consideration given to compaction levels. This results in variations in the compaction levels in nursery pots, which causes the intermittent emergence and differences in biomass characteristics (Logan and Biscoe, 1987; Wilson, 1991).
The plant organ, which is directly exposed to soil compaction, is the root whose major functions are the supply of water to the shoot, nutrients and synthesis of root hormones. The inability of the root to fulfil these functions results in an impaired shoot development. Soil compaction may restrict root growth by either mechanical resistance or insufficient aeration caused by low air-filled porosity (Grassbaugh and Bennet, 1999).
When soil is compacted, the volume of pores is reduced. However not all pores are reduced similarly. The largest pores are usually lost or reduced in size first and compaction ceases when the soil has become dense enough to withstand the applied stress without failure (Richard et al., 2001). Kooistra and Tovey (1994) observed increasing bulk density and decreasing pore volume in compacted soils. At constant water content, compaction increases the proportion of soil pores filled with water as average pore size decreases. This can lead to aeration stress, lower soil temperature and changes in biological processes. If bulk density becomes too high, it can limit plant root growth.
The specific bulk density that will adversely affect plant root growth and development depends on many factors including the parent material, soil texture, the crop grown and management practices. For silt and silt-loam soils, a bulk density of 1.55 t m-3 is often the minimum value at which root restriction may be observed (Chancellor, 1971). High bulk density values result in low pore spaces in the soil and this reduces oxygen availability at planting which in turn affects the germination percentage and emergence leading to poor stand establishment (Liacos, 1962).
Coffee plants grown on semi-compacted soil show a slightly higher rate of growth than the plants growing in fully compacted soils (Coffee Research Station Annual Report, 1984). In response to the soil compaction, plants secret abscis acid in the roots, which is transported to the stem to counter the effects of soil compaction. This results in a reduction in leaf development, expansion and stomatal conductance (Davies and Zhang, 1999).
For most plants, there is an inverse relationship between the rate of root elongation and soil strength. As a root enters a soil of high strength, its rate of elongation declines while radial expansion increases behind the root tip. Bennet (1997) found that even when root density approach high values and occupies more than 5% of the air-filled porosity, oxygen consumption rates appear to exceed the supply rate and carbon dioxide concentration increases.
Inadequate supply of oxygen reduces root growth and impairs mineral supply to the shoot. Severe soil compaction typically decreases absorption by roots of major mineral nutrients, especially N, P, and K. Mineral uptake is reduced by a combination of nutrient losses from the soil, decreased access of roots to nutrients and a lowered mineral uptake by roots (Carnell, 1977; Armstrong, 1979). In a study in Minnesota on the root-to-shoot translocation of macronutrients in maize, Engels and Marschner (1992) reported a 22% reduction in the uptake of P and K from compacted sub-soils during years when June and July precipitation was below average.
Soil compaction diverts soil conditions toward a hypoxic state that is associated with reduced aerobic microbial activity and increased denitrification. In a study of NO and N2O emissions from savannah soils through denitrification following the first simulated rains of the season, Scholes et al. (1997) observed that N2O emissions averaged 8% of total N emissions.
When roots cannot explore a large volume of soil, mineral nutrients become positionally unavailable (Kolowski, 1999). Since root growth is impeded in a compacted soil, there is a subsequent decrease in the mineral nutrient uptake by the plant. This causes nutrient deficiencies to occur which result in stunted growth of the plant and poorly developed leaves (Hussein, 2001). In addition to that, poor aeration reduces the mineralization of organic matter, which can reduce the content of nitrogen and other nutrients. Laboratory experiments indicated that the low total porosity and poor aeration at low capillary water retaining capacity of compacted soil inhibited the growth of coffee roots (Kivilia, 1998).
Coffee seed germination is induced by placing seeds in a sufficiently moist environment to absorb water. Depending on temperature and moisture, the cotyledon leaves develop after four to six weeks. Germination is first seen in the appearance of the radicle (young root) 3-4 weeks after sowing. The cotyledon appears 20-25 days later and carries the seed, which is still covered in its parchment, out of the ground. Shortly afterwards, when light covering is detached, the two cotyledons leaves open (Clowes, 2001) (Fig. 1).
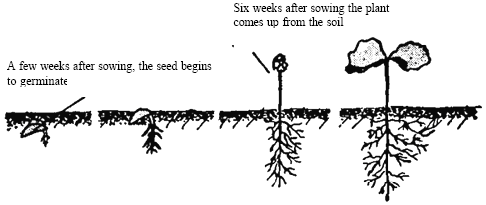 | |
| Fig. 1: | Germination of coffee seed (After Clowers, 2001) |
These cotyledons look very different from ordinary leaves. They are oval-shaped with undulating edges and 25 to 50 mm in diameter. At the same time, the terminal bud appears and produces two primary leaves. Soon after, the cotyledons die having completed their nutritional role (Clowes, 2001; Wilson, 1991).
The first lateral branch (plagiotrophic branch) appears four to six weeks after emergence, when the plant has 5 to 11 pairs of leaves. The primary branches have buds at each node that will develop either into secondary (plagiotrophic/horizontal) branches or, under certain conditions, into flowers. The young plants are ready to be transplanted when they attain a height of 20 to 30 cm (Coffee Handbook, 1987; Clowes, 2001).
The root system develops actively in the first weeks of germination when the taproot penetrates deeply into the soil and forms a great number of roots. Coffee seed germination is significantly reduced in compacted soils, as the seed requires a great deal of energy to penetrate the dense soil media (Bennett, 1997).
Soil that contains a significant portion of clay or about equal proportions of all particle sizes is more easily compacted. In addition, water acts as a lubricant, letting soil particles compress together easier. This is the reason why wet, clay soils compact more easily (Carson, 1988).
In nursery coffee seedling production, farmers often subject potted soil media to casual compaction without considering the effects of such practices on physical properties of the potted materials and their subsequent influence on the seedling emergence and biomass characteristics. Inconsistency in the casual compaction of potted soil may lead to varied and intermittent periods of seedling emergence, which pose difficulties in synchronizing transplanting events in commercialised coffee seedling nurseries. Coffee seedling biomass characteristics are fundamental determinants of coffee seedling plant sizes at pulling and yield potential in the field. It is with this background that a study was carried out for 32 weeks in the 2004-2005 summer season in order to determine the effect of different compaction levels in the nursery on the emergence and biometric characteristics of coffee. In this study, we reasoned that subjecting potted soil media to various simulated levels of compaction caused variations in the bulk densities of the soil, which, in turn, introduced different growing media conditions that were responsible for the observed unevenness in emergence and biomass characteristics.
MATERIALS AND METHODS
The Field Study Site
The study was carried out at Chipinge Coffee Research Station (20°13’ S; 32°36’ E, elev. >1500 m above sea level) near Chipinge town, 164 km south east of the city of Mutare in the Eastern Highlands of Zimbabwe. The soils at the research station are classified as Typic Paleudult and Rhodic Ferralsol in the USDA and FAO systems of soil classification respectively. The deep fine grained loamy sands are derived from quartzite and sandstones (Nyamapfene, 1991). The area lies in Natural Region II receiving rainfall ranging from 800 to 1000 mm per annum (average 900 mm per annum) with a coefficient of variation of 19%. The mean annual temperature is 21°C with significant frost occurrence in the months of June and July. The rainfall occurs during a single rainy season extending from November to April (Vincent and Thomas, 1960).
Nursery Soil Preparation and Simulation of Compaction Levels
Virgin soil was collected from the forest under tree canopy in the research station farm. The soil was dried under shed, extensively shaken to pass 2 mm sieve and mixed with fertilizer using the ratio of 50 kg of soil to 3 kg of compound J fertilizer (15%N, 5%P2O5, 20%K). Soils samples weighing 4.5 kg were collected from the fertilised soil mass and progressively placed in clay pots (25 cm top diameter, 17 cm bottom diameter, 17 cm height), which were subjected to varying soil compaction events using a 1 kg rammer. For each compaction level, three pots constituting three replications for each growing media were used. In the first set of three pots, the potted soil was not subjected to compaction (no compaction, Treatment 1 (T1)). In the second set of replicates the potted soil was rammed in five compaction events (semi compaction, Treatment 2 (T2)-) while the third set of replicated pots was subjected to fifteen ramming cycles (full compaction, Treatment 3 (T3)-). After every ramming event, the pots were replenished with fertilized soil and ram-compacted until the pots filled up. The three levels of compaction constituted three treatments for the experiment, which were blocked six times using intensity of light in the greenhouse as the blocking factor. Levels of soil compaction were quantified by measuring the bulk densities of each treatment using soil-sampling containers measuring 5 cm diameter and height. Four samples were collected from each treatment and immediately weighed on a scale. The mass and volume of the empty soil sample containers was established for computation of bulk density. The following mean bulk densities were obtained for each treatment: Treatment 1 (T1)-No compaction/control (115.56 g cm-3); Treatment 2 (T2)- Semi compaction (122.16 g cm-3); Treatment 3 (T3)- Full compaction (158.84 g cm-3).
Experiments
Effect of Soil Compaction Levels on the Emergence of Coffee Seed
Four seeds of the coffee variety SL28 were sown in each of the three replicated pots for each treatment in every block to a depth of 2.5 cm and mulched to conserve moisture. The planted pots were kept in greenhouse throughout the experiment. Seedling emergence counts commenced upon appearance of the cotyledons on the potted soil surface at weekly intervals starting on the 35th days after sowing (DAS). Recording of seedling emergence ceased on 112th day after sowing (16th week) when emergence counts leveled out.
Effect of Soil Compaction on the Seedling Biomass Characteristics
Weekly measurements of seedling stem height and girth and counts of leaves commenced a week after cessation of germination on the 17th week. Stem height and girth were measured using mm-graduated rule and callipers respectively. Measurements for stem girth were done approximately 5 cm from the soil surface in pots. Measurements and counts of these biometric characteristics ceased on the 21st week after sowing when the coffee seedlings were ready for lifting.
Root biomass was measured on the last day of recording 21 weeks after planting using the destructive sampling technique. The pots were soaked with water so as to effectively remove the whole plant from the pot. The root balls were washed clean in cold running water. The roots were cut from the stem at the point where the first primary leaves emerged and then weighed on a scale.
Data Analysis
The collected data was subjected to one-way analysis of variance (ANOVA) using GENSTAT software (Green and Margerison, 1978).
Management of the Experiment
Potted coffee seedlings were subjected to frequent irrigation events during the first 4 weeks of the experiment before thinning out to weekly irrigation up to lifting. Weeding was done by hand.
RESULTS AND DISCUSSION
Study results indicate that there was a discernible trend (F<0.001) in the effect of potted soil compaction degrees on the emergence and biomass characteristics of coffee seedlings in the nursery (Table 1-5). However, treatment separations between simulated semi and full compaction levels on the basis of seed germination counts were less convincing (Fig. 2). The first seedling emergence counts in the 5th week after sowing show that seemingly insignificant bulk density increases of 5.7% (6.6 g cm-3) and 37.5% (43.28 g cm-3) in T2 and T3 potted soils above no-compaction levels triggered uncorrespondingly massive reductions of 75 and 90% in coffee seed germination, respectively.
| Table 1: | Weekly seed emergence |
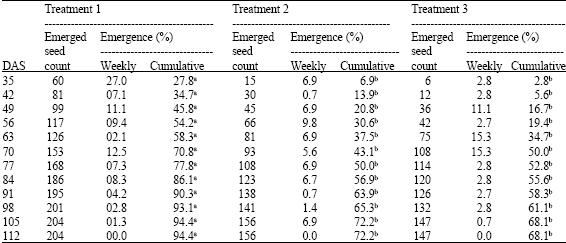 | |
| Figures superscripted a, b show significant difference at 5% level of significance | |
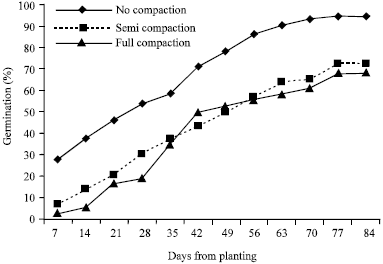 | |
| Fig. 2: | Interaction between treatments on percent germination |
Uncompacted potted soil variants (T1) comparatively exposed superior ability to propagate accelerated germination counts of 70.8% against a paltry 43.1% and 50% in T2 and T3 pots, respectively in the 7th week after sowing.
During the early stages of coffee seedling growth and development, the nutrient and energy requirements for the young plant are met by internal seed matter reserve. However, external sources of oxygen, heat energy and water for the emerging seedling energy metabolism and qualitative biomass changes should not be limiting for the successful emergence of the seedling. In the study, semi and fully compacted media soil in T2 and T3 pots generated nearly anoxic ambient environments in which the high bulk densities of 122.16 and 158.84 g cm-3 induced a reduction in air-filled porosity and amplified heat energy conductivity with associated high fluctuations of the soil climate. Such conditions in the semi and fully compacted soil entailed reduced oxygen content for energy metabolism in the germinating coffee seeds and excessive heat energy fluxes associated with nearly airless growing media, which led to subdued seedling emergence in T2 and T3 replicated pots observed in this study.
In related studies on the effect of soil compaction on growth of woody plants and grasses, Liacos (1962) and Kolowski (1999) reported reduced growth and development of plants in compacted soil due to unfavorable anaerobic high temperature fluctuation soil conditions. For similar reasons, germination counts in the no-compaction replications leveled out at a much higher count (201-204 or 94.4%) than the compacted variants (147-156 or 72.2%) after 98 days.
Study results shown in Table 2 indicate that the superior growth-supporting ability of the no-compaction coffee nursery soil media over compacted variants was perpetuated in the coffee stem height responses. It was noted, however, that while the effect of compacting potted soils on coffee seedling emergence counts significantly whittled down seedling emergence by 75-90% in the 5th week after sowing, compaction responses on seedling stem height reductions improved to a modest 15.1-24.8% in the T2 and T3 replicates in the 17th week. T1 seedlings were 30.33 and 56.27 mm taller than those in T2 and T3 pots, respectively in the 21st week after sowing.
Soil compaction inevitably causes a reduction in the air filled porosity that carries oxygen, which is required for cell energy metabolism in the cyclic ADP-ATP processes necessary for energy expending active uptake of macro and micronutrients from the Q/I equilibrium soil systems. In addition to that, an intermittent aerobic and anaerobic soil condition during and between irrigation events in compacted soils encourages elevated denitrification processes in which N is lost to the atmosphere. The subdued heights of coffee seedlings in T2 and T3 variants is, perhaps, attributable to the reduced ability of the coffee root systems to absorb vital nutrients and the amplified loss of N from the soil by denitrification.
In a study on N loss through denitrification following simulated irrigation events in the savannah Scholes et al. (1997) observed as much as 8% of N lost to the atmosphere in this process. Such significant losses of N coupled with the reduced uptake of other vital nutrients from the soil compromises biomass accumulations.
| Table 2: | Coffee seedling stem height (mm) |
 | |
| Values with superscripted letter(s) show significant difference at 5% level of significance | |
| Table 3: | Coffee seedling stem girth (mm) |
 | |
| Values with superscript letter(s) show significant difference at 5% level of significance | |
There were significant treatment signatures between non-compacted and compacted variants on stem girth measurements (F<0.001). However, treatment separation between semi and fully compacted potted soil media on the basis of stem girth was less convincing (F>0.001) (Table 3). T1 coffee seedlings recorded 0.689-0.911 mm thicker stems than those observed in T2 and T3 pots. There is considerable argument over the details of how this is achieved, but the main idea is reasonably clear. This observed trend in stem biomass characteristics was attributed to at least two growing media ambient conditions. Firstly, in the compacted variants, total voids in which the intensity factor (I) in nutrients concentration gradient necessary for their net absorption occurs is squeezed out by the action of compaction. Consequently, the nutrient supplying space of compacted media is sufficiently reduced to minimize nutrient uptake by coffee seedling roots for assimilation into stem biomass. In addition to the above, anoxic soil conditions accentuate denitrification processes in which NO3 forms of N are successively reduced to gaseous forms by facultative anaerobic bacteria and lost to the atmosphere (Scholes et al., 1997). Secondly, squeezing out total porosity entails reduction of air-filled porosity, which carries oxygen necessary for energy metabolism. Incidentally, nutrient uptake by plant roots is an energy expending process as it takes place against osmotic gradients. Compaction of growing media generates nearly anoxic (anaerobic) ambient environments, which inevitably reduces nutrient absorption by roots. In both cases, the available nutrient load for the growing coffee seedlings is severely reduced. The thin stem counts recorded in the compacted replicates were therefore not particularly surprising.
The superiority of no-compaction variants over compacted versions in biomass build-ups was perpetuated in seedling leaf emergence counts. However, the amplification of biomass fluxes in the no-compaction pots over compacted treatments was whittled down to about 12.5-25% at 17 weeks. Leaf emergence counts per seedling were higher (8 leaves) in T1 pots and lower in T2 (7 leaves) and T3 (6 leaves) at 17 weeks after sowing (Table 4). There was a comparably indiscernible trend in weekly emergence of leaves per seedling, although about 18.2-28.6% of the leaves in the treatments emerged in last two weeks of the counts. In a study on absiscic acid concentration and the associated reductions in leaf expansion and stomatal conductance in compaction-stressed plants, Davies and Zhang (1999) reported subdued leaf development and growth due to compaction.
There was a significant treatment effect (F<0.001) on seedling root biomass fluxes (Table 5). Semi compaction simulations reduced root mass per plant by a modest 1.88 g while full compaction in potted soil media dwarfed root biomass by a massive 5.34 g. Regardless of the root secretions of rhizospheral mucigels for easier penetration between abrasive soil particles, root growth impediment in the compacted media variants was excessive especially in the full compaction media treatments. Research has shown that adequate supplies of available phosphorus and nitrogen induce vigorous growth and development of lateral and fibrous roots of plants. In compacted potted soil media, the ambient dissolved nutrient holding voids in soil water porosity were effectively squeezed out by the action of compaction thereby rendering both the nutrient and oxygen supplying power of the soil voids ineffective. Oxygen is required for energy metabolism in the root systems necessary for active uptake of nutrients from reduced ambient volumes of the soil voids. Cumulatively, this had the effect of reducing assimilable quantities of soil nutrients in compacted soil media, which significantly reduced root biomass accumulations in T2 and T3 variants observed in this study.
| Table 4: | Coffee seedling leaf emergence per plant |
 | |
| Table 5: | Coffee seedling root biomass (g) per plant |
 | |
| Values with superscripted letter(s) show significant difference at 5% level of significance | |
Present study results have conclusively shown that compaction of coffee seedling growing media significantly reduces seedling emergence and biometric characteristics of coffee seedlings. Slight bulk density increases of 5.7% (6.6 g cm-3) and 37.5% (43.28 g cm-3) in T2 and T3 potted soils in excess of no-compaction levels triggered uncorrespondingly massive reductions of 75 and 90% in coffee seed germination, respectively. From these results, it can be concluded that squeezing total porosity during compaction events in the nursery comparatively created nearly oxygen-free ambient conditions in the potted media, which effectively whittled down the oxygen depleting and energy consuming active uptake of important nutrients from the soil. This effectively meant a reduction in the biomass parameter sizes of coffee seedling stems, leaves and roots.
ACKNOWLEDGMENTS
This manuscript is based on one of the many studies carried out at Chipinge Coffee Research Station under the shrewd management by Mr. Kutywayo (Head), Mr. Chingwara (Agronomist) and Mr. Masasi (Laboratory Technician). Our sincere gratitude goes to these amazing individuals for their support throughout the experiment. We also acknowledge the contributions of Mrs B. Kachigunda (Biometrician) and Miss C. Tsiko (Lecturer) at Midlands State University.
REFERENCES
- Kozlowski, T.T., 1999. Soil compaction and growth of woody plants. Scand. J. For. Res., 14: 596-619.
CrossRefDirect Link - Scholes, M.C., R. Martin, R.J. Scholes, D. Parson and E. Winstead, 1997. NO and N2O emissions from savannah soils following the first simulated rains of the season. Natl. Cycles Agro-Ecosyst., 48: 115-122.
CrossRefDirect Link