
S. Eshghi
Department of Horticultural Science, College of Agriculture,
Shiraz University, Shiraz, Iran
E. Tafazoli
Department of Horticultural Science, College of Agriculture,
Shiraz University, Shiraz, Iran
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 180-184







ABSTRACT
Flowering is an important step in crop production. Under flower-inducing conditions, biochemical or physiological changes can be recognized. Changes in mineral nutrient contents during flower bud induction and differentiation in strawberry have not been thoroughly investigated. In this study, runner plants were potted in 18 cm diameter pots and grown under non-inducing conditions (high temperature; long day). After the plants were established, half of them were kept under flower-inducing conditions (low temperature; short day) for 3 weeks, then brought back to non-inductive conditions. To determine N, P, K, Ca, Fe, Zn and Mn contents, leaves of induced and non-induced plants were collected every 3 d during 3 weeks and thereafter weekly for the period of 7 weeks. The results indicated that in induced plants total nitrogen increased from 3 to 18 days after the start of the short day treatment (DASST) and after that decreased. The amount of Ca in leaves of induced plants increased at 3, 9 and 28 DASST. The amount of Mn in leaves of induced plants was significantly higher than those of non-induced plants at 3, 9, 12 and 18 DASST. Since N, Ca and Mn were significantly higher in induced plants it seems that these elements have a decisive role during floral transition in strawberry.
PDF Abstract XML References
How to cite this article
S. Eshghi and E. Tafazoli, 2007. Changes in Mineral Nutrition Levels during Floral Transition in Strawberry (Fragaria x ananassa Duch.). International Journal of Agricultural Research, 2: 180-184.
DOI: 10.3923/ijar.2007.180.184
URL: https://scialert.net/abstract/?doi=ijar.2007.180.184
DOI: 10.3923/ijar.2007.180.184
URL: https://scialert.net/abstract/?doi=ijar.2007.180.184
INTRODUCTION
Strawberry (Fragaria x ananassa Duch.) belongs to Rosaceae family. The expansion of strawberry culture in the world for its considerable income and nutrient characteristics has drawn the attention of most growers (Hancock, 1999). Since flowering is an important stage in crop production, understanding of factors related to flowering might be a key role to improve production. During flower differentiation biochemical changes such as carbohydrate, hormones and mineral nutrient changes occur in plants (Darnell et al., 2003). Therefore, mineral nutrient changes may have a possible role in flower induction, differentiation and development in strawberry.
The finding that nitrogen (N) content of induced strawberry plants was higher than non-induced plants in shoot apices, young leaves and roots showed a complicated relationship of internal N level and flower induction (Archbold and Mackown, 1997; Yamasaki et al., 2000). However, a low N content is usually a desirable factor for flower induction in strawberry (Guttrridge, 1985). Yamasaki et al. (2000) suggested that N metabolism in shoot apex and crown of strawberry plants had a stimulatory role for flowering. The allocation pattern of 15N in their study indicated shoot apices, crowns and roots need recently absorbed N for flower initiation. Archbold and Mackown (1997) demonstrated that recently absorbed N accumulated in leaflets, while the N already present in those leaflets was remobilized and relocated to other tissues; for example, to roots. It is considered that some flower-inductive treatments can change the pattern of N remobilization. On the other hand, high level of N decreases reproductive growth (Tworkoski et al., 2001; Preusch et al., 2004).
Ca2+ has been proposed as a possible second messenger for cytokinins in their effects on the cell division process and chitinase activity (Saunders, 1992; Barwe et al., 2001). The level of Ca2+ in the root exudates increased early and transiently in response to induction of Sinapis alba plants by one LD or one displaced SD (Havelange and Bernier, 1993). A pulse of Ca2+ reached the apical bud at 30 to 40 h after the start of induction. Ca2+ was supplied to the bud via the apoplast, not the phloem and at a time when cell division is activated in the apical meristem and leaf primordia. Thus, the increased supply of Ca2+ to the bud appeared as a late and secondary induction effect. Interestingly, unlike Ca2+, Mg2+ and K+ were not supplied in greater amounts to the bud of induced plants (Havelange and Bernier, 1993).
Decreasing the K: Ca ratio from high (24:1) to low (1:5) delayed time to flowering of pot roses by 5 days. The total number of flower buds of roses was enhanced by increasing humidity and was highest at the medium (1:1) K: Ca ratio (Mortensen, 2001). Ulger et al. (2004) indicated that the N, Ca, Fe, Mn, Zn and Cu concentrations were highest during the differentiation period in flower bud of olive while K and P were higher in induction and initiation periods, respectively. Except for K and Zn, macro and micro element levels were approximately the same during both the induction and initiation periods. However, they suggested that mineral nutrients may not have a direct effect to induce flower initiation in olive.
After adding EDDHA to the nutrient solution, before floral induction and during flowering, Zn, Mn and Cu contents were significantly increased in Lemna minor. Results of above experiment showed that EDDHA promoted floral induction in L. minor under LD conditions, especially through chelating Zn, Mn and Cu and, in addition, through eliminating the antagonism between Mn and Zn chelates EDDHA (Krajncic and Nemec, 2003). He and Loh (2000) reported that cerium and lanthanum, at low concentrations promoted floral initiation, early flowering and increased the number of flowers produced. Thus, cerium and lanthanum (or other rare earth elements) may have the potential to be developed as non-hormonal flowering promoting agents for certain crop species.
It this study, investigation on the amount of mineral nutrients such as N, P, K, Ca, Fe, Zn and Mn in flower-induced strawberries were made and compared with non-induced plants.
Materials and Methods
Experimental Plant
Runner plants of cv. ‘Kordestan’ were potted in 18 cm diameter pots and grown under non-inductive conditions (28±3/22±3°C day/night; 16 h daylength) using supplemental lighting from a combination of fluorescent and incandescent lamps. After the establishment of plants, half of the plants were transferred to flower-inducing conditions (22±3°/13±3°C day/night; 8 h daylength). After a 3 weeks period of flower induction, plants were returned to non-inductive conditions again.
Mineral Nutrients Analysis
Leaf samples of induced and non-induced plants were obtained every 3 days during 3 weeks of induction period and thereafter weekly for the period of seven weeks. All samples were kept in plastic bags and transferred into the laboratory. Leaves were then washed in a weak detergent solution, rinsed in tap water several times, followed by a final rinse in distilled water. Leaves were dried at 70°C for 48 h in a forced air oven and ground using a Willey Mill to pass a 40-mesh screen for nutrient analysis. Samples were analyzed for N, P, K, Ca, Fe, Zn and Mn. Total N was determined by kjeldhal procedure (Kjeltec Auto 1030 Analyzer). For other elements, the stored samples (0.5 g) were ashed at 550°C for 5 h, dissolved in 10 mL 2 N HCl and diluted to 100 mL with distilled water. Potassium was determined using flame photometer (Corning 405). Phosphorus was determined spectrophotometrically. Calcium, Fe, Zn and Mn were determined by atomic absorption spectrophotometer (Shimadzu AA-670G, Japan) (Almaliotis et al., 2002; Niskanen and Dris, 2002; Ozden and Ayanoglu, 2002).
Statistical Analysis
The experiment was arranged in a complete randomized design with four replications. Statistical analysis was performed using MSTATC software.
RESULTS
The results in Table 1 show that the amount of Total Nitrogen (TN) in leaves of induced plants was higher than in non-induced plants in most sampling dates. In induced plants TN increased from 3 to 18 days after the start of the short-day treatment (DASST) and decreased afterwards. The amount of TN in leaves of induced plants at 3, 15, 18, 42, 56 and 63 DASST were higher than in those of non-induced plants, at corresponding dates. Total mean of nitrogen of induced plants was significantly more than that of non-induced plants (Table 1).
The amount of phosphorus in leaves of induced plants was higher than in those of non-induced plants in most sampling dates too, but these differences were not significant (data not presented). The amount of K in leaves of induced plants increased at 6, 9 and 12 DASST and then decreased to the same level as in those of non-induced plants (Table 2). In induced plants total mean of K was significantly greater than in non-induced plants. There were no noticeable changes in the amount of calcium in leaves, between induced and non-induced plants in most sampling dates (Table 3). However, the amount of calcium in leaves of induced plants increased at 3, 9 and 28 DASST.
There were no significant differences in the amount of Fe and Zn contents in leaves of induced and non-induced plants (data not presented), however the amount of Mn (mg kg-1 DW) in leaves of induced plants was significantly higher than in those of non-induced plants at 3, 9, 12, 15 and 18 DASST (Table 4). In induced plants, the highest Mn contents were at 3, 9, 15 and 18 DASST. Total mean of Mn content in leaves of induced plants (195.21 mg kg-1) was significantly higher than in non-induced plants (173.37 mg kg-1) (Table 4).
| Table 1: | Changes in total nitrogen percent (TN% DW) in leaves of induced and non-induced strawberries during flower induction and differentiation |
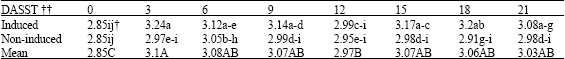 | |
| Table 1: | Continued |
 | |
| H Means in each row or column followed by the same letter(s) are not significantly different at 5% level of probability using LSD test. †† Days after the start of the short-day treatment | |
| Table 2: | Changes in potassium percent (K% DW) in leaves of induced and non-induced strawberries during flower induction and differentiation |
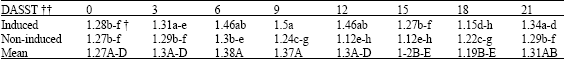 | |
| Table 2: | Continued |
 | |
| H Means in each row or column followed by the same letter(s) are not significantly different at 5% level of probability using LSD test. †† Days after the start of the short-day treatment | |
| Table 3: | Changes in calcium percent (Ca% DW) in leaves of induced and non-induced strawberries during flower induction and differentiation |
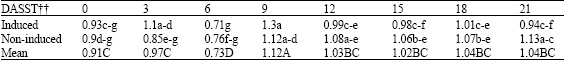 | |
| Table 3: | Continued |
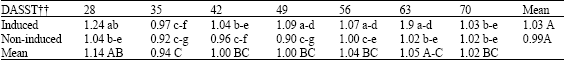 | |
| † Means in each row or column followed by the same letter(s) are not significantly different at 5% level of probability using LSD test. †† Days after the start of the short-day treatment | |
| Table 4: | Changes in Mn content (mg kg-1 DW) in leaves of induced and non-induced strawberries during flower induction and differentiation |
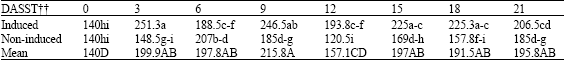 | |
| Table 4: | Continued |
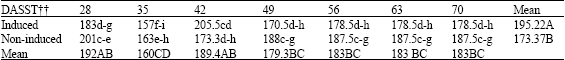 | |
| H Means in each row or column followed by the same letter(s) are not significantly different at 5% level of probability using LSD test. †† Days after start of the short-day treatment | |
DISCUSSION
Minerals analyzed of N, K, Ca and Mn showed an increase in induced plants during inductive period. Increased N concentration in induced plants may be due to increase in protein and RNA synthesis during flower induction period. This shows a complicated relationship of internal N levels and flower induction, since a low N content is usually a desirable factor for flower induction in strawberry (Guttridge, 1985). Moreover, Yamasaki et al. (2000) observed that in induced strawberry plants more N was allocated to the shoot apex, crown and root than in non-induced plants. Interestingly, although a low internal N level was reported to promote flower initiation in strawberry (Guttridge, 1985), the allocation pattern of 15N suggested that shoot apices, crowns and roots needed recently absorbed N for flower initiation (Yamasaki et al., 2000).
Archbold and Mackown (1997) demonstrated recently that absorbed N were accumulated in leaflets of strawberry, while the N already present in those leaflets was remobilized and relocated to other tissues, such as roots. It is believed that some flower inductive treatments can change the pattern of N remobilization. Tanaka et al. (1990) reported that the early process of flower initiation in Lemna paucicostata induced by nitrogen deficiency involves protein hydrolysis prior to the release of amino acids Ca2+ has been proposed as a possible secondary messenger for cytokinins in their effects on the cell division process (Saunders, 1992). The levels of Ca2+ in the root exudates increased early and transiently in response to induction of Sinapis alba plants by one LD or one displaced SD (Havelange and Bernier, 1993). Despite this enhanced supply to the shoot, the levels of this cation in mature leaves and leaf exudates were not altered. However, a pulse of Ca2+ reached the apical bud at 30 to 40 h after the start of induction. In experiment reported here, flower induction might have influenced Ca2+ absorption from the soil and hence increasing Ca2+ level in leaves.
The use of Mn-EDDHA and Zn-EDDHA resulted in the highest percentage of flowering plants (74%) in Lemna minor (Krajncic and Nemec, 2003). Krajncic and Nemec (2003) proposed that one possible pathway through which Mn-EDDHA promoted flowering was via auxin oxidase, whereas Zn-EDDHA promoted it through the enhancement of the photosynthesis and synthesis of sucrose. Since N, K, Ca and Mn were higher in induced plants it seems that these elements have an important role in flowering of strawberry.
REFERENCES
- Archbold, D.D. and C.T. Mackown, 1997. Nitrogen availability and fruiting influence nitrogen cycling in strawberry. J. Am. Soc. Hortic. Sci., 122: 134-139.
Direct Link - Barwe, S.P., M. Sathiyabama and C. Jayabaskaran, 2001. Induction of chitinase activity by exogenous cytokinins in excised dark-grown cucumber cotyledons: Involvement of Ca2+ and staurosporine-sensitive protein kinase(s) in cytokinin signaling. J. Plant Physiol., 158: 1-7.
Direct Link - Darnell, R.L., D.J. Cantliffe, D.S. Kirschbaum and C.K. Chandler, 2003. The physiology of flowering in strawberry. Hortic. Rev., 28: 352-359.
Direct Link - He, Y. and C. Loh, 2000. Cerium and lanthanum promote floral initiation and reproductive growth of Arabidopsis thaliana. Plant Sci., 159: 117-124.
Direct Link - Krajncic, B. and J. Nemec, 2003. Mechanisms of EDDHA effects on the promotion of floral induction in the long-day plant Lemna minor (L.). J. Plant Physiol., 160: 143-151.
PubMedDirect Link - Mortensen, L.M., C. Ottosen and H.R. Gislerod, 2001. Effects of air humidity and K: Ca ratio on growth, morphology, flowering and keeping quality of pot roses. Sci. Hortic., 90: 131-141.
CrossRefDirect Link - Preusch, P.L., F. Takeda and T.J. Tworkoski, 2004. N and P uptake by strawberry plants grown with composted poultry litter. Sci. Hortic., 102: 91-103.
CrossRefDirect Link - Tworkoski, T.J., T.E. Benassi and F. Takeda, 2001. The effect of nitrogen on stolon and ramet growth in four genotypes of Fragaria chiloensis L. Sci. Hortic., 88: 97-106.
Direct Link - Salih, U., S. Sahriye, K. Mustafa, E. Nisa, A. Ozgur and A. Mehmet, 2004. Determination of endogenous, sugars and mineral nutrition levels during the induction, induction and differentiation stage and their effects on flower formation in olive. Plant Growth Regulator, 42: 89-95.
Direct Link