
J. Satisha
Indian Institute of Horticultural Research, Bangalore, Karnataka, India
G. S. Prakash
Indian Institute of Horticultural Research, Bangalore, Karnataka, India
R. M. Bhatt
Indian Institute of Horticultural Research, Bangalore, Karnataka, India
P. Sampath Kumar
Indian Institute of Horticultural Research, Bangalore, Karnataka, India
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 159-164







ABSTRACT
Five grape rootstocks namely Dogridge, Salt Creek, 1613 C, St. George and VC clone were subjected to different levels of moisture stress to study their physiological responses. Control, 50% stress and 100% stress were imposed for 14 days by withholding the irrigation. Observations on relative water content, water potential, osmotic potential, specific leaf weight and gas exchange parameters like photosynthesis rate, transpiration rate, stomatal conductance and water use efficiency were recorded. All the five rootstocks wilted before completion of stress cycle at 100% stress. Dogridge and Salt Creek performed better at 50% moisture stress through maintenance of leaf turgidity as indicated by higher RWC and water potential attributing to better osmotic adjustment. The marginal reduction in photosynthesis and greater reduction in transpiration rate in these two rootstocks might have resulted in higher WUE in these two rootstocks. The higher photosynthetic rate, lower transpiration rate, higher water relation parameters and high WUE of Dogridge and Salt Creek indicates their better tolerance to drought.
PDF Abstract XML References
How to cite this article
J. Satisha, G. S. Prakash, R. M. Bhatt and P. Sampath Kumar, 2007. Physiological Mechanisms of Water Use Efficiency in Grape Rootstocks under Drought Conditions. International Journal of Agricultural Research, 2: 159-164.
DOI: 10.3923/ijar.2007.159.164
URL: https://scialert.net/abstract/?doi=ijar.2007.159.164
DOI: 10.3923/ijar.2007.159.164
URL: https://scialert.net/abstract/?doi=ijar.2007.159.164
INTRODUCTION
Indian viticulture in the past was growing grapes on its own roots of the commercial varieties. With the increasing levels of soluble salts in the soil and ground water table and also due to over exploitation of aquifers, drought and salinity are the major constraints in the semi arid regions. Due to these two major problems, it has become inevitable to grow grapes on rootstocks as they provide different root system to the vine and capable of exploiting maximum water from deeper soil layers through their extensive root system. With the use of rootstocks in Indian viticulture, many of the rootstocks are being introduced but the physiological behavior of different rootstocks overcome adverse effects of water scarcity has not been studied systematically under Indian conditions.
Water Use Efficiency (WUE) is the biomass produced over a period of time to the water transpired during the same period of time. At single leaf level, WUE is the ratio of the mole of carbon assimilated to the moles of the water transpired (Passioura, 1986). Rootstocks have a profound effect on the vigor of the scions and the size and shape of its canopy. The rooting behavior of the rootstocks has obvious effect on the water relations of the scion leaves. Basic research work has revealed that photosynthesis and stomatal conductance, which are main physiological parameters in drought tolerance, are affected by rootstocks (Sobhana, 1988). An important feature of the rootstocks in this response is, the ability of some roots to continue elongating at the water potential lower than those that inhibit shoot growth (Westgate and Boyer, 1985).
The physiological mechanisms related to drought tolerance vary from genotype to genotype. It is necessary to screen the genotypes for drought tolerance taking into consideration all aspects like photosynthesis rate, transpiration rate, WUE, stomatal conductance, relative water content etc. at different levels of water stress. Even though plants experience water stress, they have developed several adoptive mechanisms to overcome drought related harmful effects. There are lots of mechanisms operating in the plants for drought tolerance. Many of the physiological aspects involved in drought tolerance are sometimes confusing for their interpretation. Increasing Water Use Efficiency (WUE) is the common adaptation by perennial crops under drought conditions and when plants are provided with copious water the question of minimizing the water use does not arise at all. So, water use efficiency is understood under water-limited conditions (Bindu Madhava, 2000). Various physiological parameters involved in overcoming water stress are interrelated and some are not independent.
Hence to know the relationship among various physiological parameters involved in improving the water use efficiency of grape rootstocks this experiment was conducted.
MATERIALS AND METHODS
This experiment was conducted in the experimental plots of Indian Institute of Horticultural Research, Bangalore under open conditions during 2003-04. Rooted cuttings of the rootstock genotypes namely Dog ridge, Salt Creek, 1613C, St. George and Vitis champinii clone (VC clone) {Source: Germplasm collection of National Research Centre for Grapes, Pune, India} from the nursery bed were transplanted to pots of 14 diameter containing standard potting mixture of Farm Yard Manure (FYM), red earth and sand (1: 2:1). The potting mixture was porous with water holding capacity of 30%. The plants were maintained under uniform cultural practices like irrigation, fertilizer application, weeding and plant protection measures for six months. After six months plants were irrigated to field capacity before imposing different levels of soil moisture stress. To calculate the field capacity, the pot filled with known volume of potting mixture was placed in the large plastic bucket and irrigated with known quantity of water and kept for 6 h to attain the field capacity. The drained water was collected in the bucket. After six hours the amount of the drained water in the plastic bucket was measured and was subtracted from total amount of water applied. The obtained value was treated as the volume of the irrigation water that has to be applied to attain 100% field capacity (100% irrigation). Half the amount of this was considered as 50% irrigation. One set of plants were maintained without any irrigation (0% irrigation).
The above treatments were given for 14 days and periodic observations were recorded for various physiological parameters on 4th, 9th and 14th day of the stress cycle. The irrigation was done manually.
Relative water content was determined as per the procedures of Barrs and Weatherly (1962), leaf water potential was measured using water potential system CR-7, Campbell Scientific Inc, USA and leaf osmotic potential was measured using vapor pressure osmometer model 5100 C, wescor. The gas exchange parameters namely photosynthetic rate (Pn), transpiration rate (E) and stomatal conductance (gs) were measured using portable open photosynthesis system (Model LCA-3, ADC, UK). Water use efficiency at the single leaf level (A/E) was calculated using the photosynthesis and transpiration rate values. Specific Leaf Weight (SLW) was calculated using the formula leaf dry weight/leaf area and expressed in mg cm-2. The data were computed for statistical analysis and three replications were taken for each measurement.
RESULTS
No significant difference was observed for Relative Water Content (RWC) at the beginning of the experiment (Table 1). Reduction in RWC was recorded with the increased moisture stress as stress cycle progressed. On 14th day of the stress cycle at 100% irrigation VC clone had highest RWC (86.5%) followed by Dogridge. But at 50% moisture stress Salt Creek had the highest RWC followed by VC clone. None of the rootstocks could survive for 14 days without irrigation (100% stress). Water potential did not significantly differ among the rootstocks at the beginning of the experiment. But as the stress progressed there was reduction in water potential (more negative) in all the rootstocks. Among the rootstocks Salt Creek recorded highest water potential (less negative) at 50% stress followed by Dogridge, while 1613 C and St. George recorded least water potential (more negative). Similar to water potential, there was reduction in osmotic potential too, with increased soil moisture stress. On the 4th day of the stress cycle osmotic potential at no stress ranged from -1.06 to-1.34 MPa while at 50% stress it ranged from -1.20 to-1.42 Mpa. On both 9th and 14th day of the stress cycle Dogridge and salt Creek recorded maximum osmotic potential at 50% moisture stress (Table 2).
| Table 1: | Influence of moisture stress on Relative Water Content (RWC%) in grape rootstocks |
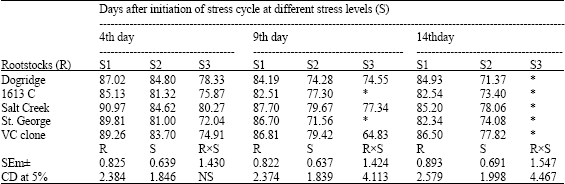 | |
| S1: control (100% irrigation); S2: 50% stress (50% irrigation); S3: 100% stress (no irrigation) *: Plants died and the observations were not recorded | |
| Table 2: | Influence of moisture stress on water potential (-Mpa) and osmotic potential (-Mpa) in grape rootstocks |
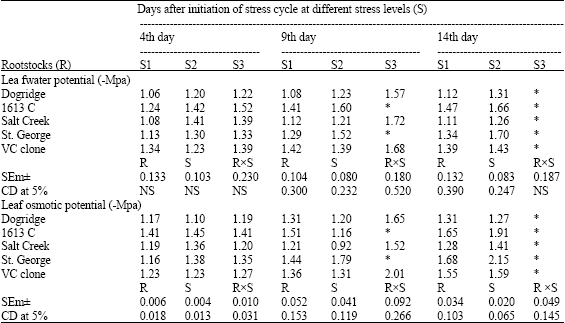 | |
| S1: Control (100% irrigation); S2: 50% stress (50% irrigation); S3: 100% stress (no irrigation) *: Plants died and the observations were not recorded | |
| Table 3a: | Photosynthetic rate (μmol m-2 sec-1) and transpiration rate (mmol m-2 sec-1) of grape rootstocks under different levelsof moisture stress |
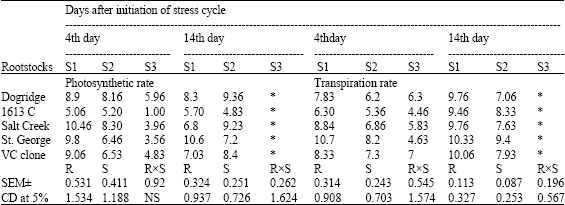 | |
| S1: control (100% irrigation); S2: 50% stress (50% irrigation); S3: 100% stress (no irrigation) *: Plants died and the observations were not recorded | |
| Table 3b: | Stomatal conductance (μmol m-2 sec-1) and instantaneous water use efficiency of grape rootstocks under different levels of moisture stress |
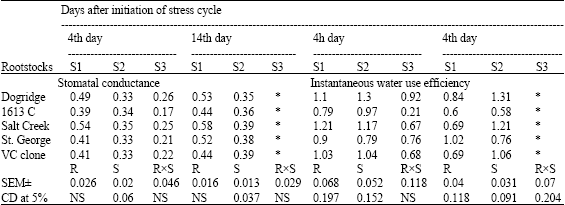 | |
| S1: Control (100% irrigation); S2: 50% stress (50% irrigation); S3: 100% stress (no irrigation) *: Plants died and the observations were not recorded | |
Gas exchange parameters like photosynthesis, transpiration rate and stomatal conductance recorded significant difference among rootstocks and stress levels on all the days of the stress cycle (Table 3a and b). Similarly instantaneous WUE significantly differed among rootstocks and stress levels. Rate of photosynthesis decreased with increased soil moisture stress. At 100% moisture stress 1613 C recorded least photosynthesis while Dogridge recorded highest photosynthetic rate. On 14th day of the stress cycle at 50% stress Dogridge recorded highest photosynthesis followed by Salt Creek while least was in 1613 C. Similarly transpiration rate reduced drastically with increased soil moisture stress in all the rootstocks. The rate of reduction in transpiration rate was maximum in Dogridge and Salt Creek while it was least in St. George and 1613 C. On 14th day of stress cycle Dogridge had least transpiration rate while St. George had highest transpiration rate at 50% stress. Drastic reduction in stomatal conductance was noticed among the rootstocks with soil moisture stress. Stress levels had significant effect on stomatal conductance at 14th day of the stress cycle. At 50% moisture stress on 14th day of the stress cycle, Salt Creek and VC clone had highest stomatal conductance while Dogridge had the least stomatal conductance.
Instantaneous WUE increased with increased soil moisture stress on 14th of the stress cycle in all the rootstocks. But on the 4th day of the stress cycle WUE was lesser under 100% stress than under 50% stress among all the rootstocks. Dogridge recorded highest WUE on 14th day at 50% stress followed by Salt Creek, while 1613 C had the least WUE.
There was reduction in specific leaf weight (SLW) with increased soil moisture stress in all the rootstocks. At 50% stress on 14th day of the stress cycle, rootstock Dogridge had highest SLW, while St. George had the least SLW.
DISCUSSION
Relative water content, leaf water potential and leaf osmotic potential in the beginning of the experiment under normal conditions were similar suggesting the similar behavior of all the rootstocks. Their ability to cope up with induced moisture stress is evident under 50% stress. The maintenance of high RWC in Salt Creek and Dogridge indicates their capacity to maintain turgidity even under soil moisture stress. This is evident from the higher water potential in these two rootstocks. The lowering of leaf osmotic potential indicates the osmotic adjustment and it varies from rootstock to rootstock. During and Dry (1995) reported that lowering of the osmotic potential in response to soil moisture stress may help in maintaining required water relations. Schultz (2000) also obtained a striking differences in the relationship between osmotic potential, turgor potential, leaf water potential and RWC of grape vines, which he related to the differences in apoplastic water content. The Dogridge and Salt Creek maintained better osmotic adjustment as indicated by leaf water potential. The increased osmotic adjustment in these rootstocks has been attributed to increased sugar and other compatible solutes (Rodrigues et al., 1993). Similarly, water stress reduced both water and turgor potential in grape vines but stressed plants tried to maintain turgor potential by osmotic adjustment. The reduction in leaf water potential induces stomatal closure and increases Abscissic acid levels in leaves (Nagarajah, 1989). In studies on contribution of various mechanisms involved in diurnal changes of osmotic potential in leaves of grape cultivar Victoria under stressed condition, dehydration accounted for 36% of the diurnal changes in osmotic potential in stressed plants. Net accumulation of solutes accounted for 73% of the diurnal changes indicating the occurrence of an active osmotic adjustment during the day in stressed plants (Patakas and Noitsakis, 1999). The present investigation also reveled increased osmotic adjustment in Dogridge and Salt Creek, which may be due to high K content in these rootstocks (data not shown) as K is an effective inorganic osmolyte. The turgid leaves with high moisture content must have helped in normal functioning of the Dogridge and Salt Creek under moisture stress conditions.
The marginal reduction in photosynthesis and greater reduction in transpiration rate under moisture stress may be due to reduced stomatal conductance similar to the findings of Reynolds and Naylor (1994), where transpiration rate and stomatal conductance reduced progressively by increasing duration of water stress in grapevines. Lakso (1985), also reported the marginal reduction in photosynthesis and greater reduction in transpiration rate in grape vines. Rootstocks can strongly influence plant response to low soil moisture stress in terms of photosynthesis, stomatal conductance and carboxylation efficiency of the grafted plants (Iacono et al., 1998). The marginal reduction in photosynthesis and greater reduction in transpiration rate in the rootstocks Dogridge and Salt Creek must have resulted in their increased WUE. Similar increase in WUE at decreased water potential was reported by Behaboudian et al. (1986) in pistachio varieties. The reduced WUE at 100% stress in all the rootstocks than at 50% stress on 4th day of stress indicates mere survival of plant will be more important than economy of water use. The reduced WUE in rootstocks in rootstocks 1613 C and St. George at 50% stress may be due to greater reduction in photosynthesis and lesser reduction in transpiration rate resulting in more water loss than the carbon assimilation. Reduction in specific leaf weight with increased moisture stress supports the findings of Rodrigues et al. (1993) where they observed reduced SLW in Rosaki grape cultivar under moisture stress.
Finally, it is summarized that, the marginal reduction in photosynthesis, lower transpiration rate, better water relation, higher specific leaf weight and increased WUE under mild water stress conditions in the rootstocks Dogridge and Salt Creek suggests their distinctness and differential sensitivity to soil moisture stress than the other three rootstocks. Thus, these rootstocks can be employed for raising vineyards under limited water availability situations to increase water use efficiency without reducing the yield.
ACKNOWLEDGMENT
We are thankful to Mr. Nageshwara Rao for his assistance in taking observation on gas exchange parameters and in laboratory analysis.
REFERENCES
- Barrs, H.D. and P.E. Weatherley, 1962. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci., 15: 413-428.
Direct Link - Behboudian, M.H., R.R. Walker and E. Torokfalvy, 1986. Effect of water stress and salinity on photosynthesis of pistachio. Scientia Hortic., 29: 251-261.
Direct Link - Iacono, F.A., Bucella and Peterlunger, 1998. Water stress and rootstock influence on leaf gas exchange of grafted and ungrafted grapevines. Scientia Hortic., 75: 27-39.
Direct Link - Patakas, A. and B. Noitsakis, 1999. Mechanisms involved in grapevine under drought conditions. J. Plant Physiol., 154: 767-774.
Direct Link - Schultz, H.R., 2000. Physiological mechanisms of water use efficiency in grapevines under drought conditions. Acta Hortic., 526: 115-136.
Direct Link - Westgate, M.E. and J.S. Boyer, 1985. Osmotic adjustment and the inhibition of leaf, root, stem and silk growth at low water potentials in maize. Planta, 104: 540-549.
CrossRef