
Nsikan-Abasi B. Udoh
Department of Biological Sciences, Faculty of Natural and Applied Sciences, Akwa Ibom State University, PMB 1169, Uyo, Nigeria
Theresa B. Ekanem
Department of Anatomical Science, Faculty of Basic Medical Sciences, University of Calabar, PMB 1115, Calabar, Nigeria
Moses B. Ekong
Department of Anatomy, Faculty of Basic Medical Sciences, University of Uyo, PMB 1017, Uyo, Nigeria
LiveDNA: 234.29958
Current Research in Neuroscience
Year: 2020 | Volume: 10 | Issue: 1 | Page No.: 1-10







ABSTRACT
Background and Objective: Artequin (ATQ) is an artemisinin combination therapy antimalarial drug, effective against all stages of Plasmodium falciparum. The aim of this study was to investigate its effects on the hippocampal microstructure and immunoreactivity, as well as the neurobehaviour of adult Wistar rats. Materials and Methods: Forty-two inbred adult male Wistar rats of average weight 200 g were divided into groups 1-6 (n = 7) of control (5 mL kg–1 of distilled water) and test groups receiving oral doses of 0.86/1.07 mg kg–1 (ATQ1), 1.71/2.14 mg kg–1 (ATQ2), 3.42/4.28 mg kg–1 (ATQ3), 6.84/8.56 mg kg–1 (ATQ4) and 13.68/17.12 mg kg–1 (ATQ5) body weight of ATQ for 3 days. Morris water neurobehavioural test was carried out for 2 days each, prior and after the administrations. The animals were sacrificed after ketamine hydrochloride anaesthesia and perfusion-fixed using 10% buffered formalin. They were routinely processed using haematoxylin and eosin and silver impregnation methods. Representative sections were immunolabeled for glial fibrillary acidic protein (GFAP). Results: Except for ATQ1 group, other ATQ groups had significantly (p<0.05) higher escape latency compared with the control. The hippocampus of the ATQ groups showed histopathological features including hypertrophy, pyknosis, karyorrhexis and less (p<0.05) cell population, with increased expressions of glial fibrillary acidic protein compared with the control. Conclusion: ATQ administration induced dose-dependent adverse effects on the hippocampal microstructure and GFAP immunolabeling, as well as on learning behaviour. This may suggests neuronal and glial pathology which may lead to neuronal and glial degeneration, ultimately resulting in altered functional processes of this brain area.
PDF Abstract XML References Citation
Received: November 06, 2019;
Accepted: December 07, 2019;
Published: December 15, 2019
How to cite this article
Nsikan-Abasi B. Udoh, Theresa B. Ekanem and Moses B. Ekong, 2020. Behavioural and Microstructural Evaluation of the Hippocampus of Adult Wistar Rats Following Artequin Administration. Current Research in Neuroscience, 10: 1-10.
DOI: 10.3923/crn.2020.1.10
URL: https://scialert.net/abstract/?doi=crn.2020.1.10
DOI: 10.3923/crn.2020.1.10
URL: https://scialert.net/abstract/?doi=crn.2020.1.10
INTRODUCTION
Artemisinin combination therapy (ACT) came to be because of the failure of previous mono- and other combination therapies and this therapy has shown greater potentials against falciparum malaria with less failure risk and resistance1. The ACT concept is based on the synergistic potential of two or more drugs with different mechanism of actions, thereby complementing each other2. One such ACT drug of interest is artequin.
Artequin is effective against all stages of Plasmodium falciparum. It is made up of two highly potent drugs, artesunate and mefloquine; which are the active ingredients of the artequin co-blister, as well as the fixed dose combination of the artequin paediatric stick pack3. Mefloquine is a 4-quinoline methanol blood schizonticide with a long acting half-life, while artesunate is a sesquiterpene lactone with a unique endoperoxide bond, usually water soluble, semi synthetic and short acting half-life and is a derivative of the Chinese medicinal herb, Qinghua2. Artesunate and mefloquine are both blood schizonticides that act by forming toxic heam complexes that damage the parasitic food vacuoles. The activities of these two drugs make their combination very effective against the Plasmodium malaria with no parasite resistance4. In this instance, artesunate reduces the parasitic load almost immediately, while mefloquine sustains the elimination, thus, preventing re-infection5.
However, mefloquine and artequin administrations have been reported to cause an increased density of reactive astrocytic processes, pyknotic nuclei formation, astrocytic glioma apoptosis, as well as reduction in pyramidal neuron density in the pyramidal cell layer of the hippocampus of rats6-8.
The hippocampus is an integral part of the limbic system whose functions include learning and memory, with roles in emotion associated with the sense of smell9,10. This brain area is reportedly affected by mefloquine and artequin treatments6,7,11,12. Even at low concentrations, mefloquine inhibits electrical coupling of neurons in the limbic region and affects their dopaminergic signals13,14. Therefore, as mefloquine is reported with adverse effects, its presence in artequin may also indicate a possibility of danger to the hippocampus. It is therefore pertinent to study the effects of exposure of this important brain area to artequin misuse. Thus, this study aimed to investigate the effects of artequin on the hippocampal microstructure and immunoreactivity, as well as the neurobehaviour of adult Wistar rats.
MATERIALS AND METHODS
Animal handling: Forty-two inbred adult male Wistar rats of average weight 200 g were obtained and housed in the Animal House of the Faculty of Basic Medical Sciences, University of Uyo, Nigeria. The animals were housed in 14 standard home cages (40 cm×35 cm) with wire gauze roof and wood shavings beddings. The study was carried out in the month of June, with room temperature between 27-30°C. The animals were exposed to 12:12 h light/dark cycles and fed with normal commercial pelletized growers mash (Grand Cereal Ltd., Nigeria) and clean water ad libitum. The animals were allowed to acclimatize for 14 days before commencement of the experiment. Ethical approval was obtained from the Ethics Committee of the University of Uyo and the animals were handled according to the guidelines of the National Institute of Health (NIH) of the United States of America for the regulation of laboratory animals15.
Artequin (J0004912, Acino Pharmaceutical Limited, Switzerland) was obtained from the University of Uyo Pharmacy and each co-blister tablet was dissolved in 100 mL of distilled water. The therapeutic dosages for the rats were determined against the therapeutic doses for humans, which is 600/750 mg kg–1 of artequin. The drug suspensions were administered to the animals based on their body weight orally with the aid of orogastric tubes.
Animal grouping and administration of artequin: The Wistar rats were divided into 6 groups of seven animals each, modified from Udoh et al.7. Groups 1 was the control, while groups 2-6 served as the treatment groups. Each co-blister tablet of the drugs, artequin (ATQ, artesunate, 600/mefloquine, 750 mg kg–1) dissolved in 100 mL of distilled water was administered to the animals orally with the aid of orogastric tubes for 3 days according to their body weights (Table 1).
Morris water test: Morris water maze was used to test for learning prior and after the drug administrations modified from Vorhees and Williams16.
| Table 1: | Weights and dosages of drugs administered to the rats for 3 days |
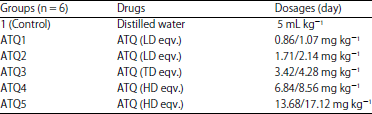 | |
ATQ: Artequin, LD eqv: low dose equivalent, TD eqv: Therapeutic dose equivalent, HD eqv: High dose equivalent | |
Briefly, the water maze was constructed out of a circular rubber basin that measured 150 cm in diameter and 30 cm in depth. The pool was filled to a depth of 15 cm and left to sit overnight in order to achieve room temperature. The water was made opaque with the addition of liquid milk to ensure camouflage of the white escape platform which was submerged 1 cm under the water. A square solid block (10 cm×28 cm) covered with white gloves was used as the escape platform in the maze. The pool was divided into four quadrants of Northwest, Northeast, Southwest and Southeast and their boundaries marked on the edges of the pool with masking tapes and labelled North, South, East and West. The pool was located in a room measuring 36 cm×34 cm. The room was dimly lit with 60 Watts electric lamp, with several posters on the walls and furniture that acted as visual cues. Three scorers who remained in the same position throughout the experiment days observed the performance of the animals in the water maze.
Before drug administration (BDA), the animals were exposed to acquisition training for 2 days. The water was adjusted appropriately such that the platform was covered by 1 cm of water (invisible platform). The platform was placed in the centre of the northwest quadrant. Each animal received four trials of 60 sec/day. The starting positions of the animals were predetermined which prevented any sequence of two trials to be repeated by the same animal. Start positions were at the boundaries of the quadrants (West, North, East and South). Each rat was removed from its holding cage using a small clean 500 mL plastic container to minimize handling stress. The animal was then placed into the water at the appropriate start position, facing the centre of the pool. The rat was then permitted to explore the pool and to search for the hidden escape platform for 60 sec. When the animal located the platform, the timer was stopped and the animal was allowed to explore the platform for 5 sec before being removed using the plastic container and placed back in the holding cage.
If the animal did not find the platform during the allotted time, the animal was guided onto the platform. Once on the platform, the rats were permitted to visually explore their surrounding for 20 sec at which point they were picked up in the plastic container and returned to the appropriate holding cage. The next rat was then placed in the pool and the same procedure repeated. Each animal completed four trials/day over 2 days.
After drug administration (ADA), there was acquisition training again carried out 24 h later (on day 1), while the probe trial was introduced on the second day. At this time, there was no escape platform at all in the maze. Each animal completed the 4 trials of 60 sec. Each rat was placed in the maze from one of the 4 possible start positions and allowed to explore the pool. The time to platform (latency) was noted and used to determine learning in the animals.
Tissue processing: Immediately after the Morris water test, the animals were sacrificed after they were anaesthetized with 60 mg kg–1 of ketamine-hydrochloride (#50155, Rotex Medica, Trittau, Germany). Intracardial perfusion with phosphate-buffered saline (PBS, 2M, pH 6.4), was carried out on the animals by means of a cannula and then perfusion-fixed using 10% buffered formalin. On complete perfusion, the skull was opened and the brain of the animal removed and post-fixed in 10% buffered formalin for 48 h. The whole brain was further routinely processed for histological studies by means of haematoxylin and eosin17 and silver impregnation methods18. Representative sections were also used for glial fibrillary acidic protein immunolabelling19. Sections were then viewed under the light microscope and photomicrographs were obtained using the microscope camera linked to a computer. Cellular population estimation was determined by the ImageJ™ software.
Statistical analysis: One-way analysis of variance was used to analyze all the data, followed by a post hoc Tukey’s test. All analysis was done using Graphpad Prism for Windows (version 5.01, San Diego California, USA). Data at probability level p<0.05 were regarded as significant and are presented as Mean±Standard error of mean.
RESULTS
Morris water test: Before drug administration (BDA), the high doses ATQ4 and ATQ5 groups had a significantly (p<0.05) higher escape latency in trial day 1 compared to the control group. No significant difference in escape latency was observed for the other ATQ groups. On trial day 2, there was no significant difference in escape latency between the ATQ groups and the control group. After drug administration (ADA), ATQ2, ATQ3, ATQ4 and ATQ5 groups had significantly (p<0.05) higher escape latencies in trials days 1 and 2 compared with the control and ATQ1 group. However, there was no significant difference in the escape latency between ATQ1 group and the control group on both trial days (Fig. 1).
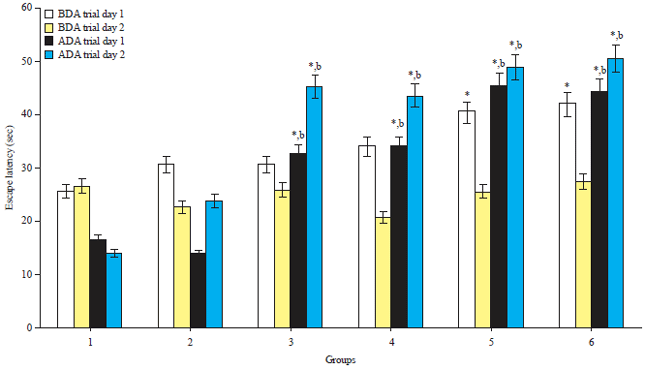 | |
| Fig. 1: | Escape latency before and after the drug administrations |
Mean±standard error of mean, *Significantly different from the control group at p<0.05, bSignificantly different from the ATQ1 group at p<0.05, all others are not significantly different from the control group at p<0.05, BDA: Before drug administration, ADA: After drug administration | |
Histomorphology/histomorphometry observation: Haematoxylin and Eosin (H and E): Sections of the hippocampus of control rat showed normal histological appearance with pyramidal neurons having triangular shaped soma (Fig. 2a). The sections of the hippocampus of the ATQ2 and ATQ3 groups showed hypertrophied and pyknotic neurons in the pyramidal cell layer with the polymorphic and molecular layers appearing unaffected compared with the control group (Fig. 2b, c). The section of the hippocampus of ATQ3 group showed hypertrophied neurons with some having pyknotic and karyorrhectic nuclei throughout the three cortical layers (Fig. 2d). The sections of the hippocampus of ATQ4 and ATQ5 groups showed atrophied neurons with some having pyknotic and karyorrhectic nuclei in the pyramidal and molecular layers compared with the control group, while the polymorphic layer appears unaffected (Fig. 2e, f).
The population of cells of the hippocampus was significantly (p<0.05) lower in ATQ2 group compared with the control group. No significant difference was observed between the other ATQ groups and the control group. However, ATQ3 group ATQ4 cellular population was significantly (p<0.05) higher than the other ATQ2 group (Fig. 3).
Silver impregnation: Sections of the hippocampus of control group animals showed pyramidal cell neurons with pyramidal shaped soma in the pyramidal cell layer with non-expressions of silver grains in the molecular and polymorphic layers (Fig. 4a). Sections of the hippocampus of ATQ1 and ATQ2 groups showed hypertrophied pyramidal cells with argyrophilic nuclei. The cells in the other 2 cortical layers appear unaffected (Fig. 4b, c). The section of the hippocampus of ATQ3 group showed degenerating nuclei with silver grains in the molecular and pyramidal cell layers, the polymorphic layer appear unaffected (Fig. 4d). The section of the hippocampus of ATQ4 group showed mostly hypertrophied and pyknotic nuclei in the molecular and pyramidal cell layers, the polymorphic layer appear unaffected (Fig. 4e). Also the sections of the hippocampus of ATQ5 group showed degenerating pyramidal and polymorphic layer nuclei, the molecular layer appear unaffected (Fig. 4f) compared with the control group.
Immunolabeling: The section of hippocampus of the control animals showed expression of glial fibrillary acidic protein throughout the cortical layers (Fig. 5a). The sections of the hippocampus of the ATQ1 and ATQ2 groups showed increased expressions of glial fibrillary acidic protein in the astrocyte’s soma and processes in the molecular and pyramidal cell layers compared to the control (Fig. 5b, c).
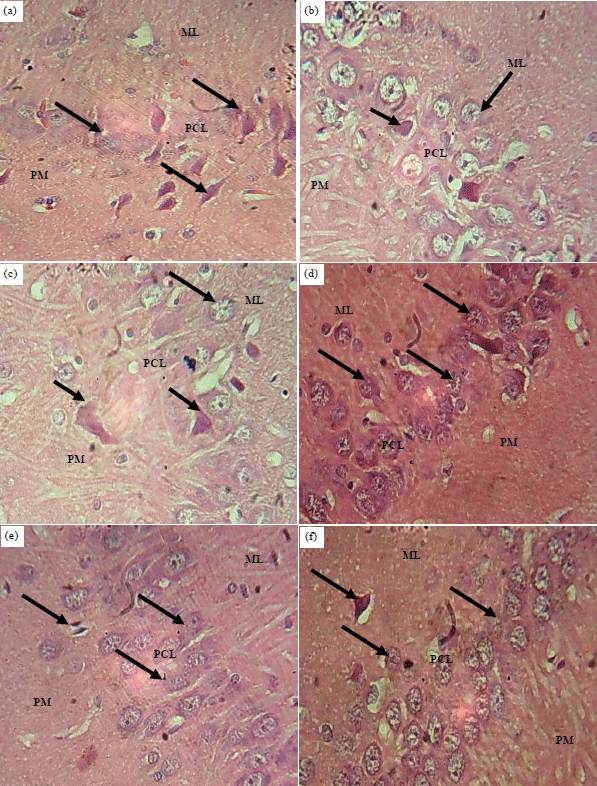 | |
| Fig. 2(a-f): | Sections of the hippocampus of the control and test groups: (H and E), ×400, (a) Control, (b) ATQ1, (c) ATQ2, (d) ATQ3, (e) ATQ4 and (f) ATQ5 |
Control group showing normal histological appearance of pyramidal neurons (arrows), ATQ1-ATQ2 groups showing hypertrophied (short arrows) and pyknotic neurons (arrows), ATQ3-ATQ5 groups showing hypertrophied neurons (short arrows) with some having karyorrhectic nuclei (arrows), ML: Molecular layer, PCL: Pyramidal cell layer, PM: Polymorphic layer | |
Also, the sections of hippocampus of the ATQ3, ATQ4 and ATQ5 groups, showed increased expressions of glial fibrillary acidic protein astrocyte’s soma and processes throughout the cortical layers compared to the control (Fig. 5d-f).
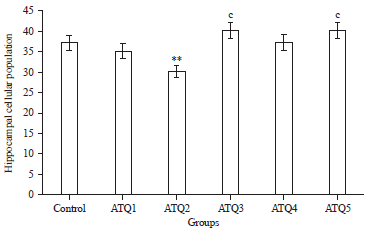 | |
| Fig. 3: | Hippocampal cellular population in the control and test groups |
Mean±standard error of mean, **Significantly different from the control group at p<0.01, cSignificantly different from ATQ3 at p<0.05 | |
DISCUSSION
This study investigated the hippocampal microstructure and neurobehaviour of adult Wistar rats following artequin administration. The results showed that apart from the ATQ1 group, all the other ATQ groups had significantly (p<0.05) higher escape latency. Their hippocampi showed histopathological features including hypertrophy, pyknosis, karyorrhexis and significantly (p<0.05) less cell population, as well as increased expressions of glial fibrillary acidic protein compared with the control.
In the present study, Morris water maze test results showed that prior to administration of the drug on trial day 2, there was no significant difference in the time spent to locate the platform between the ATQ and control groups. This result indicates that the animals were of the same baseline. However, after administration of the drug, the ATQ2, ATQ3, ATQ4 and ATQ5 groups had significantly (p<0.05) higher escape latencies in both ADA trials days 1 and 2 compared with the control and ATQ1 groups. However, there was no significant difference in the escape latency between ATQ1 and the control groups on both trial days. These results show that the administration of increasing dosages of ATQ to the animals in these groups may have resulted in hippocampus trauma in these rats; hence their ability to locate the hidden platform was impaired.
The Morris water maze is used to test spatial learning and memory and rely on cues to navigate and locate a hidden escape platform20,21. This behavioural test is particular sensitivity to the effects of hippocampal lesions in rats. Hippocampal place cells are the primary substrate of the spatial memory abilities underlying spatial navigation processes involved in the hidden-platform Morris water maze learning. As such is necessary for acquisition and retrieval of spatial information, as well as for consolidation and storage22,23.
The spatial learning impairment of these rats’ hippocampi in the present study after ATQ administration is related to the volume of damaged hippocampal tissue in these ATQ groups. The result in this present studies relates to earlier reports that hidden-platform acquisition and probe trial performances are impaired in hippocampal-lesion patients and animals22,23.
Histological findings of the hippocampus of the animals in the ATQ groups showed alterations including cell atrophy, hypertrophy, karyorrhexis and pyknosis in the artequin groups compared to the control group. The presence of pyknotic nuclei in the pyramidal cells of the hippocampus, as well as vacuolation of the surrounding glia indicates neuronal death24,25. Neuronal changes and damage in the hippocampus are manifested in the form of discontinuation of the nuclear membrane, pyknosis of nuclei and vacuolation of neuropil around the damaged neurons26. These degenerative features, especially the pyramidal neurons of the ATQ groups in the present study may hinder the participatory role of the pyramidal cell as the primary excitation unit in hippocampal function.
The cellular population of the hippocampus was significantly lower (p<0.05) in ATQ2 group compared with the control. However, ATQ3 and ATQ4 groups cellular population was significantly (p<0.05) higher than the ATQ2 group. These imply that ATQ administration may have lead to neuronal death resulting in a decrease cellular population in the ATQ2 group. Global cerebral ischemia is reported to induce selective acute neuronal injury of the CA1 and CA3 regions of the hippocampus. The type of cell death that ensues may include different programmed cell death mechanisms including necroptosis. The present study is in agreement with an earlier report by Vieira et al.27, who observed that oxygen-glucose deprivation induced a death receptor-dependent component of necroptotic cell death in primary cultures of hippocampal neurons, which upregulated the receptor-interacting protein kinase 3 (RIP3) mRNA and protein levels, with a concomitant increase of the RIP1 protein. Thus, damage to the hippocampus may impair its functions, as well as its inter-relational functions with other brain structures.
Silver impregnation sections of the hippocampus of the animals in the ATQ groups in this present study, showed alterations in their cytoarchitecture with strong affinity for the silver ions, thus indicating degenerating neurons and axonal terminals. The silver impregnation method is designed for the detection of degenerating neurons in fixed tissue sections of the central nervous system.
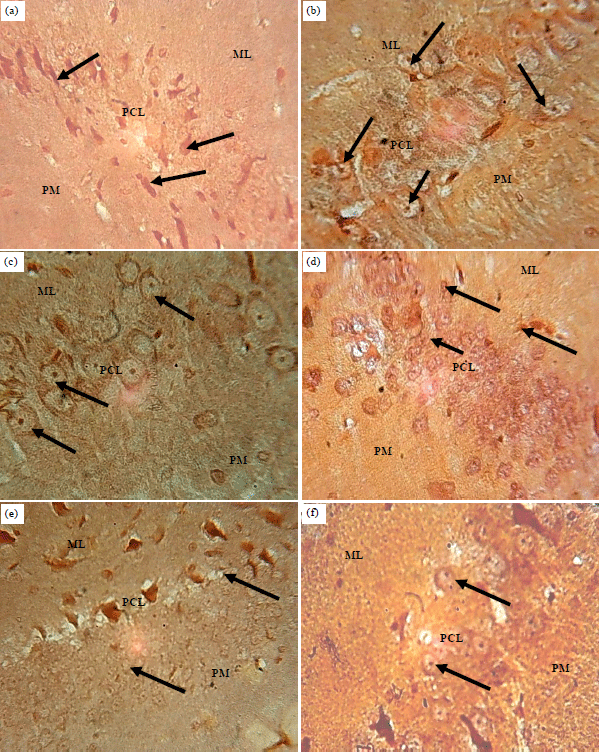 | |
| Fig. 4(a-f): | Sections of the hippocampus of control and test group: Silver Impregnation, ×400, (a) Control, (b) ATQ1, (c) ATQ2, (d) ATQ3, (e) ATQ4 and (f) ATQ5 |
Control group showing normal pyramidal cell neurons, astrocytes and oligodendrocytes (arrows), ATQ1 group showing degenerating nuclei (arrows), ATQ2 and ATQ3 group showing hypertrophied pyramidal cells which are also argyrophilic (arrows), ATQ4 and ATQ5 showing hypertrophied pyramidal cells with argyrophilic nuclei (arrows), ML: Molecular layer, PCL: Pyramidal cell layer, PM: Polymorphic layer | |
The principle of this technique is based on the findings that certain components of neurons undergoing degeneration become particularly argyrophilic28. These may be due to the formation of argyrophilic metallic grains that are visible under the light microscope upon reduction, as well as depositions similar to perivascular amyloid plaques seen in Alzheimer’s diseases in rats models17. These argyrophilic manifestations may bring about cognitive decline, behavioural abnormalities, as well as episodic memory loss in the animals17,29.
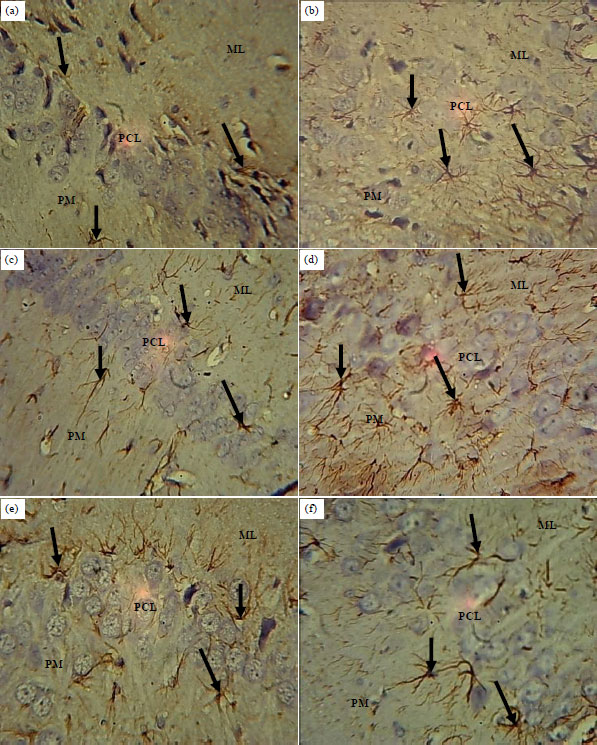 | |
| Fig. 5(a-f): | Sections of the hippocampus of control and test groups: GFAP; ×400, (a) Control, (b) ATQ1, (c) ATQ2, (d) ATQ3, (e) ATQ4 and (f) ATQ5 |
Control rats showing normal expression of GFAP (arrow) throughout the cortical layers, ATQ1-ATQ5 groups showing increased GFAP expressions in the astrocytes soma and processes, ML: Molecular layer, PCL: Pyramidal cell layer, PM: Polymorphic layer | |
Immunohistochemical reactivity of the hippocampal cortical layers of rats treated with increasing dosages of ATQ in this present study, revealed GFAP expressing reactive astrocytes indicating increased reactive GFAP astrogliosis proliferations. Trauma to the brain, as in the ATQ groups result in hypertrophy of astroglial processes and a significant increase in the astrocyte cytoskeleton and the upregulation and synthesis of intermediate filament proteins, especially glial fibrillary acidic protein (GFAP)30. Brain damage very rapidly turns most of the astroglial cells into GFAP-expressing ‘reactive’ astrocytes, which is critical for the development of reactive astrogliosis.
Reactive GFAP astrogliosis is a defensive brain reaction aimed at isolating damaged areas from the rest of the central nervous system tissues, reconstruction of the blood-brain barrier and facilitation of the remodelling of brain circuits in areas surrounding the lesion31. Hence, the reactive GFAP expressing astrocyte may have acted to cushion this damage done to this brain region. Astrocytes located immediately around the damaged zone undergo a robust hypertrophy and proliferation, which ultimately ends up in complete substitution of previously existing tissue architecture with a permanent glial scar. Reactive astrocytes in these areas produce chondroitin and keratin, which inhibit axonal regeneration and thus prevent nerve processes from entering the damaged zone. They also release mucopolysaccharides, which eventually cement the areas of damage and produce the astrocytic scar32. The expression of GFAP in the hippocampus in the present study suggests neuronal and glial pathology which may lead to neuronal and glial degeneration, ultimately resulting in altered functional processes of this brain area.
CONCLUSION
This study showed that oral administration of ATQ had significantly higher escape latency, which is associated with adverse learning behaviour. The hippocampal microstructure showed hypertrophy, pyknosis, karyorrhexis and significantly less cell population, which are associated with tissue trauma and injury. There were also increased expressions of glial fibrillary acidic protein known to occur in brain tissue injury. These effects were dose dependent.
SIGNIFICANCE STATEMENT
This study observed that artequin overdose may lead to adverse consequences including learning deficit and hippocampal damage. This study will help in clinical practice, as well as the society at large to understand the implication of artequin abuse. However, more thoroughput techniques still needs to be applied.
ACKNOWLEDGMENTS
We appreciate the assistance of the technical staff of the Histology Laboratory of the Department of Anatomy, University of Uyo, Uyo, Nigeria. We are also thankful to the management of the University of Uyo, for providing the needed research facilities and that of the Akwa Ibom State University for supporting this study.
REFERENCES
- Dow, G., R. Bauman, D. Caridha, M. Cabezas and F. Du et al., 2006. Mefloquine induces dose-related neurological effects in a rat model. Antimicrobial Agents Chemoth., 50: 1045-1053.
Direct Link - Gofton, T.E., A. Al-Khotani, B. O'Farrell, L.C. Ang and R.S. McLachlan, 2011. Mefloquine in the treatment of progressive multifocal leukoencephalopathy. J. Neurol. Neurosurg. Psychiat., Vol. 82.
CrossRefDirect Link - Ekanem, T., E. Salami, M. Ekong, M. Eluwa and A. Akpantah, 2009. Combination therapy anti-malaria drugs, mefloquine and artequin induce reactive astrocyte formation in the hippocampus of rats. Int. J. Health, Vol. 9.
Direct Link - Udoh, N.A.B., T.B. Ekanem, M.B. Ekong, A.I. Peter and A.O. Akpantah, 2014. Hippocampal glial degenerative potentials of mefloquine and artequin in adult wistar rats. Int. J. Brain Sci., Vol. 2014.
CrossRefDirect Link - Liu, B. and J. Yu, 2017. Effect of apoptosis inducing factors on artesunate-induced astrocytic glioma apoptosis. Int. J. Clin. Exp. Med., 10: 4450-4456.
Direct Link - Voss, J.L., D.J. Bridge, N.J. Cohen and J.A. Walker, 2017. A closer look at the hippocampus and memory. Trends Cognit. Sci., 21: 577-588.
CrossRefDirect Link - Heshmati, M., S.A. Golden, M.L. Pfau, D.J. Christoffel and E.L. Seeley et al., 2016. Mefloquine in the nucleus accumbens promotes social avoidance and anxiety-like behavior in mice. Neuropharmacology, 101: 351-357.
CrossRefDirect Link - Nevin, R.L., 2018. Measurement of mefloquine exposure in studies of veterans' sleep disorders. J. Clin. Sleep Med., 14: 1273-1274.
CrossRefDirect Link - Allison, D.W., R.S. Wilcox, K.L. Ellefsen, C.E. Askew and D.M. Hansen et al., 2011. Mefloquine effects on ventral tegmental area dopamine and GABA neuron inhibition: A physiologic role for connexin‐36 GAP junctions. Synapse, 65: 804-813.
CrossRefDirect Link - Steffensen, S.C., K.D. Bradley, D.M. Hansen, J.D. Wilcox and R.S. Wilcox et al., 2011. The role of connexin‐36 gap junctions in alcohol intoxication and consumption. Synapse, 65: 695-707.
CrossRefDirect Link - NRC, DELS and ILAR, 2011. Guide for the Care and Use of Laboratory Animals. 8th Edn., National Academies Press, Washington, DC., ISBN: 9780309154000, Pages: 246.
Direct Link - Vorhees, C.V. and M.T. Williams, 2006. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nature Protocols, 1: 848-858.
CrossRefDirect Link - D'Hooge, R. and P.P. De Deyn, 2001. Applications of the morris water maze in the study of learning and memory. Brain Res. Rev., 36: 60-90.
CrossRefPubMedDirect Link - Higaki, A., M. Mogi, J. Iwanami, L.J. Min and H.Y. Bai et al., 2018. Predicting outcome of Morris water maze test in vascular dementia mouse model with deep learning. Plos One, Vol. 13.
CrossRefDirect Link - Cho, Y.H., E. Friedman and A.J. Silva, 1998. Ibotenate lesions of the hippocampus impair spatial learning but not contextual fear conditioning in mice. Behav. Brain Res., 98: 77-87.
CrossRefDirect Link - Moser, E.I., M.B. Moser and B.L. McNaughton, 2017. Spatial representation in the hippocampal formation: A history. Nature Neurosci., 20: 1448-1464.
CrossRefDirect Link - De Souza Pagnussat, A., M.C. Faccioni‐Heuser, C.A. Netto and M. Achaval, 2007. An ultrastructural study of cell death in the CA1 pyramidal field of the hippocapmus in rats submitted to transient global ischemia followed by reperfusion. J. Anat., 211: 589-599.
CrossRefDirect Link - Faheem, N.M. and A. El Askary, 2017. Neuroprotective role of curcumin on the hippocampus against the structural and serological alterations of streptozotocin-induced diabetes in Sprague Dawely rats. Iran. J. Basic Med. Sci., 20: 690-699.
Direct Link - Mitra, N.K., V.D. Nadarajah and H.H. Siong, 2009. Effect of concurrent application of heat, swim stress and repeated dermal application of chlorpyrifos on the hippocampal neurons in mice. Folia Neuropathol., 47: 60-68.
Direct Link - Vieira, M., J. Fernandes, L. Carreto, B. Anuncibay-Soto and M. Santos et al., 2014. Ischemic insults induce necroptotic cell death in hippocampal neurons through the up-regulation of endogenous RIP3. Neurobiol. Dis., 68: 26-36.
CrossRefDirect Link - Nobakht, M., S.M. Hoseini, P. Mortazavi, I. Sohrabi, B. Esmailzade, N.R. Roosh and S. Omidzahir, 2011. Neuropathological changes in brain cortex and hippocampus in a rat model of Alzheimer’s disease. Iran. Biomed. J., 15: 51-58.
Direct Link - Ferrer, I., G. Santpere and F.W. van Leeuwen, 2008. Argyrophilic grain disease. Brain, 131: 1416-1432.
PubMedDirect Link - Pekny, M. and M. Nilsson, 2005. Astrocyte activation and reactive gliosis. Glia, 50: 427-434.
CrossRefDirect Link - Auld, D.S. and R. Robitaille, 2003. Glial cells and neurotransmission: An inclusive view of synaptic function. Neuron, 40: 389-400.
CrossRefDirect Link - Seifert, G., K. Schilling and C. Steinhäuser, 2006. Astrocyte dysfunction in neurological disorders: A molecular perspective. Nature Rev. Neurosci., 7: 194-206.
CrossRefDirect Link