
Ibrahim A.A. Abou Ayana
Department of Dairy Research, Food Technology Research Institute (FTRI), Agriculture Research Centre, Ministry of Agriculture, Giza, Egypt
H.M. El-Shabrawy
Animal Production Research Institute (APRI), Agricultural Research Center (ARC), Dokki, Giza, Egypt
Current Research in Dairy Sciences
Year: 2015 | Volume: 7 | Issue: 1 | Page No.: 1-17







ABSTRACT
Investigating dairy goat feeding is a good way to increase milk yield and improve milk composition and dairy products. Thirty five lactating Zaraibi goats were divided into five groups; seven in each, besides control, two levels of canola seeds 5 and 10% and two levels of sunflower seeds 7 and 14% were used instead of 5, 10, 7 and 14% of concentrated feed for group 2, 3, 4 and 5, respectively. Ruminal individual Volatile Fatty Acids (VFA's), some blood constituents (Urea-N, Total protein, Albumin and Globulin) of lactating goats moreover, milk production, major components of milk and Fatty Acid (FA) profile were determined. Milks resulting from these treatments were manufactured Labneh. Significantly (p<0.05), VFA's, acetic acid and the ratio of acetate to propionate increased, but NH3-N, propionic acid and butyric acid decreased for goats fed canola or sunflower diets than the control. A significant difference was noticed among treatments in blood total protein. All treatments improved the milk yield, 4% fat corrected milk (4% FCM). The most of saturated fatty acids reduced while unsaturated and Conjugated C18:2 increased. High Sunflower Seeds (HSFS) treatment was the greatest sensory evaluation with redacting of "goaty odour" in other treatments except the control.
PDF Abstract XML References Citation
Received: May 15, 2015;
Accepted: July 08, 2015;
Published: May 03, 2019
How to cite this article
Ibrahim A.A. Abou Ayana and H.M. El-Shabrawy, 2015. Improvement of Milk Composition and Properties of Labneh Using Oil Seeds as Supplement for Lactating Zaraibi Goat Rations. Current Research in Dairy Sciences, 7: 1-17.
DOI: 10.3923/crds.2015.1.17
URL: https://scialert.net/abstract/?doi=crds.2015.1.17
DOI: 10.3923/crds.2015.1.17
URL: https://scialert.net/abstract/?doi=crds.2015.1.17
INTRODUCTION
Goat milk and its products have three-fold significance in human nutrition: (1) feeding more starving and malnourished people in the developing world than from cow milk. (2) treating people afflicted with cow milk allergies and gastro-intestinal disorders, which is a significant segment in many populations of developed countries and (3) filling the gastronomic needs of connoisseur consumers, which is a growing market share in many developed countries (Haenlein, 2004). Goat milk is a healthy, functional food, has higher digestibility, certain therapeutic value in medicine and human nutrition, reduction of chronic diseases risks and modification of physiologic it is not surprising that goat milk has a higher price than cow milk (Park and Haenlein, 2007). However, the national U.S. dairy products judging procedures list "goaty" as one of the four odor characteristics of bad versus good milk and dairy products.
Goats are important domestic animals in many parts of the world, because of their adaptability to different environmental conditions and utilizing poor quality feed stuffs. Egyptian Zaraibi goats famous for higher yield of milk and lambs thus they are considered economical dairy animals.
Oil seeds are used as supplements in concentrate feed mixtures for lactating and growing animals. Among those, canola and sunflower seeds are characterized by high levels of lipids and proteins. Whole canola seed contains approximately 55% lipid, of which over 85% is 18-carbon fatty acids, 18:1 is the predominant fatty acid (>60% of total fatty acids, Ackman, 1990). Whole canola and sunflower seeds contain approximately 34 and 22% lipids, respectively as reported by El-Kholy et al. (2005).
Lactating animals fed oil seeds supplemented diets have increased milk yield (El-Shafie and El-Ashmawy, 2010). Feeding oil seeds to lactating dairy cows changed the proportion of unsaturated fatty acids in milk fat, with increase as high as 40% (Jacobs et al., 2011). Sunflower seeds increase the proportion of unsaturated fatty acids in milk compared to cows fed no supplemental fat (Petit, 2003).
Canola oil is high (61%) in the monounsaturated fatty acids, oleic acid (C18:1). However, canola oil has a much lower level of the saturated fatty acid, palmitic acid (7%). Canola oil contains a moderate level (22%) of the polyunsaturated omega 6 fatty acid, linoleic acid (C18:2) and appreciable amount (11%) of the polyunsaturated omega 3 fatty acid, alpha-linolenic acid (C18:3). Sunflower oil is rich in polyunsaturated omega 6 fatty acid, linolenic acid (C18:2, 66% of total fatty acids). However, data are not available to determine the optimal or maximal amount of oil seeds to incorporate in diets fed to lactating goats producing large amount of milk.
Labneh is a concentrated yogurt, a semisolid fermented dairy product, produced by removing part of yogurt whey, this product is likable, popular, known as Labneh in the Middle East, Suzme yogurt in Turkey, strained yogurt in Greece, and other countries of Europe. Labneh usually contains moisture 76-78%, TS 22-25%, fat 7-11%, protein 8.5-10%, Lactose 3.3-3.45% and Ash 1.3-1.4%.
Therefore, this study was designed to investigate the effects of canola or sunflower seeds in the diets of lactating goats on nutrient digestibility, some rumen fermentation characteristics, blood constituents and their reflection on milk yield, milk composition, milk fatty acids profile, properties of the Labneh made from those samples of milk and economic efficiency.
MATERIALS AND METHODS
This work was carried out at EL-Serw Experimental Station, Animal Production Research Institute, Agricultural Research Center, Ministry of Agriculture, Egypt.
Thirty-five lactating Zaraibi goats (after weaning period) were balanced for body weight, milk yield, days after calving and parity. The animals were randomly divided into five groups (7 does each) and received one of five experimental diets (Table 1). Diets were formulated to contain (% of DM) 40% berseem hay and 60% Concentrate Feed Mixture (CFM) (control, diet 1), Low Canola Seeds (LCS, 5% of CFM as DM, diet 2), High Canola Seeds (HCS, 10% of CFM as DM, diet 3), Low Sunflower Seeds (LSFS, 7% of CFM as DM, diet 4) and High Sunflower Seeds (HSFS, 14% of CFM as DM, diet 5). The CFM consisted of 22% yellow maize, 26% wheat bran, 32% undecorticated cotton seed cake, 5% linseed meal, 9% rice bran, 3% molasses, 2% limestone and 1% sodium, chloride.
| Table 1: | Formulation of the experimental diets for lactating goats (DM%) |
 | |
LCS: low canola seeds, 5%, HCS: High canola seeds, 10%, LSFS: Low sunflower seeds, 7% and HSFS: High sunflower seeds, 14%, BH: Berseem hay, CFM: Concentrated feed mixture, CS: Canola seeds, SFS: Sunflower seeds | |
In addition to 2 g chamomile/head/day of all animals, Canola seeds (Brassica napus) variety Serw 4 used in this study are characterized by free Erucic acid in oil, low glucosinolate in the seeds. They are produced in EL-Serw, Agriculture Research Station, oil crops section, Agricultural Research Center, Ministry of Agriculture, Egypt; Sunflower seeds were also obtained from there. ABT-culture (Lactobacillus acidophilus; Bifidobacterium bifidium and Streptococcus salivariussub sp. thermophilus) was obtained from Chr. Hansen-(Denmark). Sodium chloride was obtained from El- Naser Company for salt, Egypt. Chamomile and plastic containers were purchased from local market.
The experiment started after weaning and began after 10 days of adaptation to the tested diets and lasted for 14 weeks. The experimental diets were formulated and offered to the experimental animals in quantities to cover their requirements according to NRC (1989) recommendations. Daily amounts of feed were readjusted every two weeks based on the average milk production and body weight changes. The berseem hay and concentrate were offered two times daily.
Animals were hand milked twice daily at 5.00 a.m. and 5.00 p.m. and milk yield of individual animals was recorded at each milking. Milk was sampled biweekly from two consecutive milkings and composited according to milk yield. Major milk components of samples were analyzed.
At the end of digestibility trial, rumen fluid samples were taken from three animals in each group just before offering morning meal, 2, 4, 6 and 8 h post-feeding using stomach tube. Rumen-fluid pH was immediately measured on a fresh aliquot then samples were filtered through two layers of surgical gauze. Total Volatile Fatty Acids (VFA’s) concentrations were estimated using steam distillation method (Warner, 1964), Molar proportions of VFA’s were determined according to Erwin et al. (1961).
During the last week of the experimental period, blood samples were collected in heparinized test tubes from the jugular vein from three animals of each group before morning feeding, 2 and 4 h post-feeding. Blood samples were centrifuged immediately at 3500 revolution per min (rpm) for 15 min. to separate blood plasma and stored at -20°C until further analysis. Blood plasma was analyzed for urea-N (Patton and Crouch, 1977), total protein (Peters, 1968) and albumin (Webster, 1974). Globulin concentration was calculated by difference (total protein-albumin).
Separately, 20 kg milk of each experimental group, was heated at 95°C/15 min, immediately cooled to 40°C then inoculated with 4% active ABT- culture and incubated at 40°C until completely coagulation, fermented milks from all treatments were cooled to 10°C overnight, fermented milk each treatment mixed and put into sterilized cloth bags, was hung in refrigerator room at 6-8°C to remove apart of whey for 12 h, 0.5% NaCl was added to the contents of bags, the resultant Labneh was filled into plastic containers and stored at 6-8°C for 30 days. Samples were analyzed chemically, microbiological and organoleptically at fresh and after 1, 2, 3 and 4 weeks.
Analysis of milk and Labneh: All chemical analysis of milk, chemical and microbial analysis of Labneh were conducted according to AOAC (2003) include Total Solids (TS), Fat, Solids Not Fat (SNF), Lactose, Protein, Ash, pH, Titratable Acidity (TA) Dry Matter (DM) salt, Total Nitrogen (TN), Soluble Nitrogen (SN) None Protein Nitrogen (NPN). However, methyl esters of fatty acids of milk lipids were analyzed according to the method described by Chouinard et al. (1997). Microbial testes include total viable count, lactic acid bacteria, spore-forming bacteria, coliform, psychrotrophic bacteria, moulds and yeasts, Staph. aureus and Salmonella spp. Total carbonyl compounds was determined according to Basset and Harper (1958).
Statistical analysis: Data were subjected to statistical analysis by the computer program of SAS (1996) using the General Linear Model (GLM).
RESULTS AND DISCUSSION
Beginning, volatile fatty acids such as acetic, propionic, butyric, isobutyric, valeric, isovaleric, 2-methylbutyric, hexanoic and heptanoic acid, are produced in several parts of the gastrointestinal tract by microbial fermentation of dietary fiber. In addition to their involvement as the major source of energy, volatile fatty acids also serve as building blocks for milk synthesis; acetate is a necessary component in the formation of milk fat, while propionate is used for glucose production, which is needed for synthesis of lactose. In ruminants, propionate is also the major substrate of hepatic gluconeogenesis (Herdt, 1988). Therefore it was necessary determine VFA’s, the effects of oil seeds (CS and SFS) on ruminal individual VFA’s are presented in Table 2. Molar proportions of acetic acid were higher (p<0.05) being on average 55.37 vs. 51.62 mol/100 mol) for goats fed the oil seeds diets than for those fed control diet, respectively. In contrast, molar proportions of propionic acid and butyric acid were lower (p<0.05), being on average 24.64 and 16.33 vs. 27.54 and 17.27 mol/100 mol) for goats fed the oil seeds diets than those fed control diet, respectively. Ruminal proportions of valeric acid were not different among tested diets. Proportions of iso-butyrate were higher (p<0.05) for goats fed LCS diet than those fed other diets. Proportions of iso-valerate were higher in ruminal fluid of goats fed on the HSFS diet, but were lower for goats fed on the HCS diet. Control, LCS and LSFS diets recorded intermediate values between HSFS and HCS diets. The acetate to propionate ratios were greater (p<0.05) for goats fed oil seeds diets than those fed the control diet (average 2.25 vs. 1.87 mol/100 mol, respectively). It is clear that the highest values for acetic acid and acetate to propionate ratios were recorded for HCS diet (56.37 mol/100 mol and 2.41, respectively). These results are similar to those of Aldrich et al. (1997) who found that ruminal proportions of acetic acid and acetate to propionate ratios were higher (p<0.01) and those of propionic acid were lower (p<0.01) for cows fed canola seeds diets than for cows fed the control diet. They also, found that valerate molar proportions were not different among treatment groups (diets supplemented with fat vs. control diet). Schauff and Clark (1992) found that molar percentages of ruminal propionate tended to be lower in cows fed high fat rations. Onetti et al. (2002) reported that molar proportions of butyrate and valerate were increased when supplement fat diet was fed for lactating cows.
Concerning the effect of sampling time, proportions of acetate, iso-butyrate, valerate, iso-valerate and A: P in rumen liquor of goats were significantly (p<0.05) increased after four h post feeding compared to those recorded before feeding. While proportions of propionate and butyrate were significantly (p<0.05) decreased after 4 h post feeding.
Generally, the fermentation parameters indicated quite clearly that the pattern of NH3-N (not shown) and VFA’s values followed the reverse trend to the obtained pH values (not shown)and reflect the pattern of fermentation in the rumen. It appears that the relatively high ether extract in diets did not adversely affect the fermentation pattern in the rumen.
| Table 2: | Impacts of feeding the experimental diets on the means of ruminal individual volatile fatty acids (mol/100 mol) for Zaraibi goats |
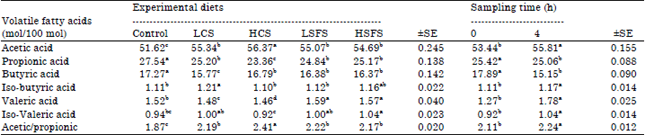 | |
Means within the same row having different superscripts are significantly different (p<0.05), HCS: High canola seeds, LSFS: Low sunflower seeds, LCS: Low canola seeds, HSFS: High sunflower seeds, ±SE: Standard error | |
| Table 3: | Mean values of the concentration of some blood constituents of lactating goats fed the different experimental diets |
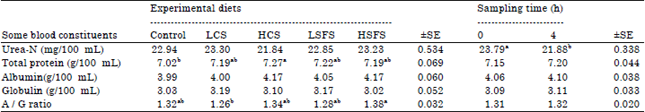 | |
Means within the same row having different superscripts are significantly different (p<0.05), LCS: Low canola seeds, HCS: High canola seeds, LSFS: Low sunflower seeds, HSFS: High sunflower seeds, A/G ratio: Albumin/Globulin ratio | |
Some blood constituents: The results in Table 3 indicated that the blood urea-N concentration values did not differ among the tested diets. In contrast, higher (p<0.05) values of total protein concentration were recorded for goats given diets containing oil seeds than those given control diet. The obtained results in this study were close to the previous findings reported by Petit (2003) who found that blood urea-N concentrations were similar for cows fed diet containing flax seed compared with those fed diet containing sunflower seeds. On the other hand, (Chichlowski et al., 2005) found lower blood urea-N (p<0.08) for cows fed canola seed (14% of diet DM) than those fed diet no containing canola seeds. Johnson et al. (2002) found that feeding cows on diet containing oil seeds increased blood urea-N compared to those fed diet no containing oil seeds. Bernard et al. (2009) noticed increase in blood urea-N concentrations (p<0.05) for lactating Alpine goats fed maize silage based diets containing sunflower seeds oil (6.1% of diet DM) or linseed oil (6.2% of diet DM) compared with the control (no additional oil).
The concentrations of albumin and globulin in blood were slightly increased for goats fed diets containing CS and SFS compared to those fed control diet but the differences were not significant. Moreover, sampling time as well as its interaction with tested diets did not show significant effect on all measured blood parameters, expect the sampling time for urea-N concentration which was significant (p<0.05). Generally, all values were within the normal ranges for healthy goats.
Milk yield and composition: The results in Table 4 indicated that milk and 4% Fat Corrected Milk (FCM) yields significantly (p<0.05) increased by 17.9 and 17.0% for goats fed oil seeds than for goats fed control diet, respectively. Johnson et al. (2002) showed that milk and 3.5% FCM yields were improved by 22 and 17% for lactating cows fed diets containing oil seeds than those fed diet no additional oil seeds, respectively. Diet ewes with flaxseed increased (p<0.05) milk yield by 8% (Zhang et al., 2006). Similarly, Bernard et al. (2009) and EL-Shafie and EL-Ashmawy (2010) noticed an increase in milk production by lactating goats fed diets containing sunflower seeds oil or full sunflower seeds as compared to those fed diet containing no additional oil or sunflower seeds. Linseeds, Sunflower seeds, lupine seeds and soybean increased the goats’ milk production (Chilliard et al., 2003). In contrary, on average over the 12 week period, supplementation with Whole Cottonseed (WCS) decreased the yield of milk (10%) (Grainger et al., 2010). In dairy cows, dry matter intake can be decreased by gut peptides released in response to extra fat, this effect has not been observed in ewes and goats. Milk yield increases in dairy cows, ewes and goats in a curvilinear manner with increasing amounts of dietary fat (Marin et al., 2013). When fat supply is low, the increase in milk yield is probably caused by a higher energy intake. However, milk yield decreases when fat supply is high, which may be related to diminished energy availability for milk production due to negative effects on rumen digestion and/or dry matter intake.
| Table 4: | Effects of feeding the experimental diets on the means of milk yield and milk components |
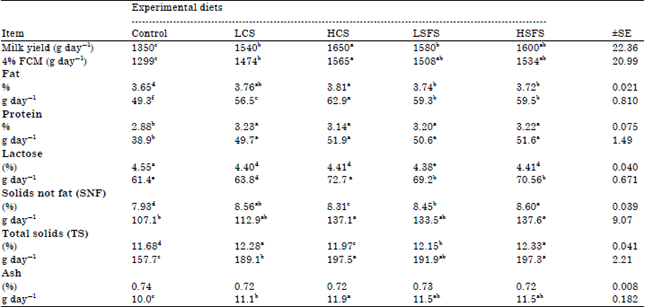 | |
Means within the same row for each effect having different superscripts are significantly different at p<0.05 | |
On the other hand, Toral et al. (2010) reported that milk yield was not significantly affected by lipid addition to diets lactating ewes. Similar, results confirmed by Luna et al., (2008) when they supplement ewes’ diets with flax seeds and sunflower oil. Diet ewes with sunflower seed had no effect on milk yield (Zhang et al., 2006). Cow’s milk yield was not affected by oilseed (Crushed sunflower, flax or canola seeds) supplement (Beauchemin et al., 2009).
Concerning effect of levels of CS and SFS on milk yield, it could be noticed that adding and increasing seeds level in rations tended to significantly (p<0.05) higher milk yield. Moreover, the increasing in milk yield with levels of SFS was not significant. The 4% FCM yield followed the same trend as milk yield. EL-Shafie and EL-Ashmawy (2010) indicated higher response in milk yield by lactating goats when they were fed a diet containing 10% sunflower seeds as compared to those fed a diet containing 5% sunflower seeds. On the other hand, milk yield and 3.5% FCM yields were similar for cows fed high and low levels oil seeds (Johnson et al., 2002). Sunflower oil levels did not effect on milk yield of lactating ewes (Toral et al., 2010).
Milk components were significantly (p<0.05) increased for goats fed diets containing CS and SFS than those fed the control diet, except milk lactose percentage, which showed an increment (p<0.05) for control diet compared with CS and SFS diets. These results were due to the change of rumen fermentation and the change in volatile fatty acids. Acetate is a necessary component in the formation of milk fat as a result of which the increment of Acetic acid increased the level of milk fat, as well as the shortage of lactose due to lower Propionic acid (Table 2), while the increase of protein might due to the increase of blood urea, albumin and globulin ( Table 3). These results are in agreement with the previous results of (Chilliard et al., 2003) who found that feeding goats on linseeds, sunflower seeds, lupine seeds and soybean increased milk components. However, Johnson et al. (2002) and Chichlowski et al. (2005) found that feeding lactating dairy cows diets supplemented with oil seeds decreased milk fat percentages. Recently, Grainger et al. (2010) confirmed that supplementation with whole cottonseeds decreased the milk fat (11%). However, unprotected lipids negatively affected milk fat content in dairy cows, but not in dairy ewes and goats (Marin et al., 2013). On the other hand, Aldrich et al. (1997) reported that feeding lactating dairy cow diets supplemented with canola seeds (11.2% of DM) maintained or increased milk fat percentages. The percentage of milk fat tended to decrease as the level of CS was increased. Supplementing rations of goats with sunflower seeds increased milk protein but soybeans decreased it, cited from (Chilliard and Ferlay, 2004).
Fatty Acid (FA) profile of the milk
Saturated Fatty Acid (SFA): Firstly, Milk fatty acids have a dual origin: they are either taken up from plasma lipoproteins (60% of the fatty acids secreted in milk (Chilliard et al., 2000) or they are synthesized de novo in the mammary gland from acetate and 3-hydroxybutyrate. The main metabolic pathway involves two key enzymes: Acetyl CoA Carboxylase (ACC) and Fatty Acid Synthetase (FAS). FAS synthesize FAs that feature up to 16 carbon atoms. The cellular and molecular factors that regulate the chain length of the fatty acids synthesized, within the same species or between the various ruminant species, have yet to be identified. As an example, the factors that determine the specificities of goat milk, which contains more 8- and especially 10-carbon atom fatty acids (Glass et al., 1967), are very little known. Pre-formed fatty acids are transported in plasma as Non-Esterified Fatty Acids (NEFA) or as triglyceride-rich lipoproteins. Lipoprotein Lipase (LPL) permits triglyceride hydrolysis and FA uptake by the mammary gland. As shown in Table 5, percentages (g/100 g of total FA) of all short (C2-6) and medium-chain (C8-12) saturates were significantly (p<0.05) reduced in supplemented diets with oilseeds (CS or SFS) compared to control diet, except Butyric C4:0 (lightly increased). Contrary to these results, sunflower, Lupine seeds and soybean increased short (C2-6) but they and linseeds increased medium-chain (C8-12) in goat’s milk fat (Chilliard et al., 2003). Tridecanoic (C13), Myristic (C14), Pentadecamic (C15), Palmitic (C16), Arachidic (C20) took the same trend (declining) but Margaric (C17) increased in LCS and HSFS treatments (0.74a and 0.61b resp.) against 0.32 and 0.47 for HCS and LSFS, respectively.
| Table 5: | Fatty acid (FA) profile of the milk from goats fed the experimental diets |
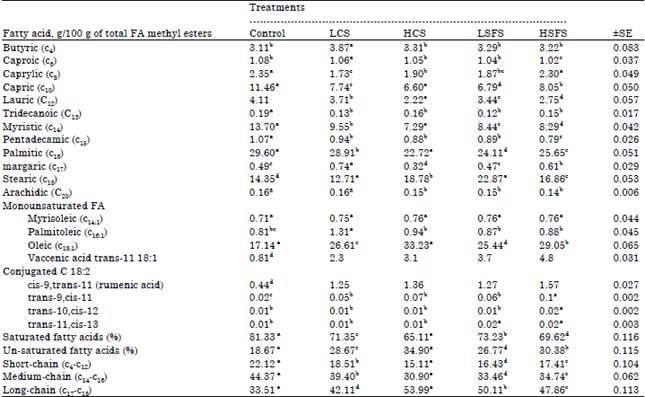 | |
Means within the same row within each category having different superscripts are significantly different at (p<0.05) | |
Stearic (C18) increased in HCS, LSFS and HSFS too. Feeding LCS significantly (p<0.05) decreased Stearic (C18) concentration, but when goats fed HCS, LSFS and HSFS, Stearic (C18) significantly reached 18.78b, 22.87a and 16.86c, respectively. These findings agree with Toral et al. (2010). Goat’s milk fat normally has 35% of medium chain fatty acids (C6-C14) compared to cow milk fat 17% include three strong smelling acids Caproic (C6), Caprylic (C8), Capric (C10), which significantly (p<0.05) decreased in our study. The results that related "fatty acids, C4-C16" were similar findings (Chilliard et al., 2003) when supplementing of goats diets with sunflower seeds, soybean, linseeds and lupine seeds. Clearly significantly (p<0.05) saturated fatty acids (SFA)% decreased, CS treatments reduced SFA more than SFS, LCS, HCS, LSFS and HSFS reduced SFA with 9.9, 16.22, 8.1 and 11.7%, respectively. It is now well established that increases in the supply of long-chain FA to the mammary gland alter the synthesis of short- and medium-chain saturates (Kadegowda et al., 2009). The decrease of short and medium chain FA and saturated FA indicated lower de novo FA synthesis within the mammary gland (Bernard et al., 2009). Depressed proportion of short and medium chain FA and saturated FA because of the decrease in de novo FA synthesis has been attributed to a direct inhibition of mammary acetyl-coenzyme-A carboxylase activity (Palmquist and Jenkins, 1980). This inhibition may be as the result of formulation of trans isomers, which is a result from biohydrogenation of long chain and unsaturated FA in the rumen (Chichlowski et al., 2005) and increased mammary uptake of long chain and unsaturated FA from plasma triacylglycerol (Storry, 1988).
Reduction of fatty acids (C6-C14) improved the characteristics of goats’ milk, "goaty" flavor reduced; this is a perfect goal in this work, whereas acceptability of goats’ milk and its products increased. This is very beneficial for public health, heart diseases and arteries, because high intakes of saturated FA could be related to an increased risk of cardiovascular diseases and development of the metabolic syndrome (Shingfield et al., 2008).
There were also important changes in milk content of almost all monounsaturated FA, all determined acids, Oleic acid (cis-9 C18:1), Myrisoleic (C14:1), Palmitoleic (C16:1) and Vaccenic acid trans-11 18:1 levels were significantly (p<0.05) increased with addition of CS or SFS to diets of goats, canola seeds increased Palmitoleic (C16:1) and Oleic acid (cis-9 C18:1) than Myrisoleic (C14:1) and Vaccenic acid trans-11 18:1 reverse SFS treatments, Vaccenic acid trans-11 18:1 increased by 3.3 and 5.8 times in CS and SFS, respectively compared to the control. This may be related to the higher level of unsaturated C18:1 fatty acid in the CS diets (Chichlowski et al., 2005). Furthermore, feeding canola seeds (11.2% of diet DM; 55% lipid) increased C18:1 67% in milk fat (Aldrich et al., 1997). Also, Chichlowski et al. (2005) reported that cows fed canola seeds (14% of diet DM; 34% lipid) increased C18:1 in milk fat by nearly 25%. The differences in the proportion of C18:1 in milk fat between control diet and CS diets in the current study could be attributed to rapid availability of oil in the rumen and its potential to reduce fiber digestibility (not shown). Piperova et al. (2000) reported that in cows fed a high-grains, low forage diet that contained soybean oil (5% DM), the increases in trans-10 C18:1 and trans-10, cis-12 C18:2 in milk fat were associated with significantly lower levels of de novo synthesized fatty acids. Monounsaturated fat consumption has been associated with decreased Low-Density Lipoprotein (LDL) cholesterol and possibly increased High-Density Lipoprotein (HDL) cholesterol. e.g., Oleic acid may hinder the progression of adrenoleukodystrophy (ALD), a fatal disease that affects the brain and adrenal glands and Oleic acid may be responsible for the hypotensive (blood pressure reducing) effects of olive oil Teres et al. (2008). Vaccenic acid is also found in human orbitofrontal cortex of patients with bipolar disorder and schizophrenia (McNamara et al., 2007) as well as lowered total cholesterol, LDL cholesterol and triglyceride levels Conjugated C18:2 that include cis-9 trans-11, trans-9 cis-11, trans-10, cis-12 and trans-11, cis-13 fatty acids influenced by the addition of oilseeds to diets of goats, cis-9 trans-11 (rumenic acid), trans-9 cis-11 FA significantly (p<0.05) increased compared to the control whereas no significant (p<0.05) difference appeared in levels of trans-10 cis-12 and trans-11 cis-13 FA except at HSFS treatment. Because of the transient nature of RA formed in the rumen, endogenous synthesis from VA through mammary Δ9-desaturase is considered the principal origin of this CLA isomer in ruminant milk (Shingfield et al., 2008). Probably because the usually high ruminal BH of linoleic acid can be further increased with marine lipid supplements (up to 90% according to Sinclair et al., 2010). The notable rise in milk trans-C18:1 would also support this hypothesis. Conjugated linoleic acid (CLA) has been shown to have beneficial health effects; it may protect against cancer, improve blood cholesterol and protect against coronary heart disease (Huth et al., 2006).
Regardless the level of CS and SFS, the percentage of saturated FA was lower in goats fed CS or SFS (68.2 and 71.4%, respectively) than those fed control diet (81.3%). While, the percentage of unsaturated FA was higher in goats fed CS or SFS (31.8 and 28.6%, respectively) than those fed control diet (18.7%). In the present study, concentration of saturated FA was decreased in goat fed LCS, HCS, LSFS and HSFS by 12.3, 19.9, 10.0 and 14.4%, respectively. Whereas, concentrations of unsaturated FA increased by 53.6, 86.9, 43.4 and 62.7% respectively.
Concerning the effect of levels of CS and SFS on percentages of saturated, short-chain and medium-chain fatty acids, results indicated that they were decreased (p<0.05) when goats were given HCS and HSFS diets than when they were given LCS and LSFS diets. On the other hand, the percentages of unsaturated and long chain fatty acids increased. Johnson et al. (2002) reported that the concentrations of unsaturated FA in milk tended to increase as the level of oil seeds was increased in the diet.
These changes in milk FA are similar to those of reported by Johnson et al. (2002), Abo-Donia et al. (2003) and Petit (2003) who fed canola seeds, cotton seeds, heated soybeans, calcium salts of palm oil and sunflower seeds to dairy cows, feeding oil seeds reduced (p<0.05) the proportion of both short-chain and medium-chain fatty acids and increased (p<0.05) the proportion of long-chain fatty acids in milk fat. Soybean oil and linseed oil in diets of dairy cows increased long-chain FA in milk fat Jacobs et al. (2011). In the present study, the secretion of long-chain FA increased in response to CS and SFS by 46.18 and 43.33%, respectively. Bernard et al. (2009) reported increases in the secretion of long-chain FA which ranged between 109 and 111% in goats fed maize silage based diets supplemented with plants oil.
Yield of Labneh: Addition of oil seeds to diet’s goats significantly (p<0.05) affected the yield of Labneh along storage period (Table 6).
| Table 6: | Mean effect of feeding the experimental diets on yield of Labneh stored at refrigerator (p<0.05) |
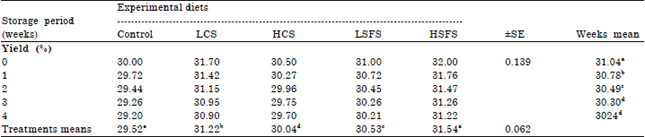 | |
Means within the same row or column within each category having different superscripts are significantly different at (p<0.05) | |
Generally, the yield mean of HSFS treatment was the greatest followed by LCS, LSFS then HCS, they were 31.54, 31.22, 30.53 and 30.04 % against 29.52 for the control. Surely, this increase due to the high increases of total solids, fat and protein of milk; Table 4. HSFS and LCS treatments increased the yield 6.84 and 5.76%, respectively. On the other hand, there was a significant difference between the periods of storage until the third week; there was not a significant (p<0.05) difference after that.
Some chemicals properties of fresh and stored Labneh were summarized in Table 7, TA of all treatments and control, gradually increased along the storage period to reach 2.21, 2.08, 2.21 and 1.82% for LCS, HCS, LSFS and HSFS, respectively against 2.08% for control at the end of storage period. The low levels of oil seeds (LCS and LSFS) recorded higher TA than HCS and HSFS that was the lowest TA. The low levels of oilseeds probably activated lactic acid bacteria. Statistical analysis (p<0.05) of acidity means showed a different significant among the control and all treatments, HSFS was 1.73%, the lowest unlike LSFS was the highest (2.10%). On the other hand, the pH-values took a contrast trend for TA, whereas LSFS was the lowest pH 3.52 but HSFS was the highest pH 3.64.
Naturally, the chemical composition of Labneh was reflection for the milk composition. Data listed in Table 7 explain that there were significant (p<0.05) differences between the control and treatments in TS or F/DM. HSFS treatment was the greatest in TS (30.74) average (30) antithesis the control that recorded the lowest TS (28.80) average (28.02).
| Table 7: | Some chemical composition (pH-values, acidity%, TS%, Fat/DM, Salt/DM) of Labneh |
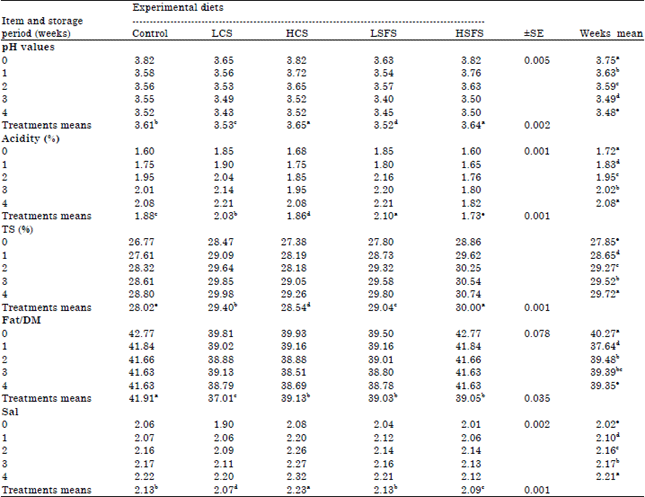 | |
Means within the same row or column within each category having different superscripts are significantly different at (p<0.05) | |
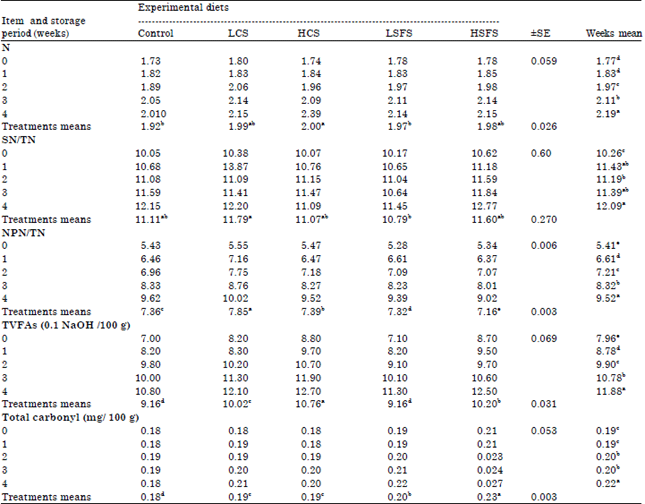
| Table 8: | Nitrogen analysis (TN, SN/TN, and NPN/TN), TVFAs and Total carbonyl TC mg/ 100 g of Labneh during cold storage |
 | |
Means within the same row or column within each category having different superscripts are significantly different at (p<0.05) | |
Normally, the concentration of TS increase with increasing duration of storage period, so a significant difference appeared between TS of samples during the storage period, the highest concentrations were at the end of storage. The mean of TS at the end of fourth week was 29.72 against 27.85 at fresh samples of Labneh. The milk fat reduced in all treatments compared the standard ration; consequently, Fat/DM decreased in all Labneh samples whether fresh or stored except HSFS that was similar the control. Regard to average of Fat/DM, no significant difference among HCS, LSFS and HSFS but it was detected between the control and other treatments.
Data of Table 8 show that significantly (p<0.05), TN, SN (not shown) and SN/TN nearly took the same trend in analyzed Labneh samples. HCS recorded the greatest TN (2.0a), no significant difference was appeared among all other treatments include the control. Furthermore, TN significantly (p<0.05) increased gradually with increment the storage period to reach maximum level (2.19) after month. Nevertheless, there were not significant (p<0.05) differences among all treatments and control in the ratio SN/TN.
Generally, NPN/TN significantly (p<0.05) increased along storage, HSFS was the lowest (9.02) average (7.16) followed by HSFS, LSFS, Control, HCS then LCS, which had the highest NPN/TN (10.02) average (7.85a) whilst the weeks’ means heightened along storage to reach maximum concentration after four weeks (9.52a).
TVFFA content: TVAs contents depend on lipolytic and proteolytic analysis during storage period, chemical composition of dairy products, some enzymes and microorganisms act these decompositions. Data presented in Table 8 confirmed that supplementing goat’s diet with oilseeds influenced on total volatile free fatty acids (TVFFAs) contents of Labneh. In general, TVFFAs significantly (p<0.05) increased with prolongation of storage. The increases in TVFAs could be explained on the basis that the increase in the cheese proteolysis as amino acids can serve as precursors for the development of certain volatile fatty acids (Salem et al., 2007). Control and LSFS were the lowest followed by LCS, HSFS then HCS that had the greatest TVFFAs. Another noticing, concentration of TVFFAs incremented and recorded significant variation among the means of analysis periods whilst reached the greatest level after month (11.88). Clearly, the disparity in TVFAs may be due to the difference in the milk composition particularly fatty acids, as well as the impact of microorganisms that play an important role in the formation of TVFAs.
Total Carbonyl (TC) contents: From the same table, Total carbonyl contents significantly (p<0.05) increased gradually during storage period, HSFS was the best followed by LSFS, LCS and HCS then the control. There was no significant difference between the treatments of canola seeds but there was significant difference between CS and SFS. This may due to the effect of milk composition and fatty acids on organisms affecting the synthesis of TC compounds. TC differed based on activity of microorganisms and storage period.
Microbiological analysis: (Data not shown) pathogenic bacteria include Salmonella spp., Staph. aureus and Coliform group were not detected in fresh or stored Labneh, this might be due to the efficient heat treatment of milk (95°C for 15 min) and high sanitation conditions during manufacture and storage samples. Counts of Aerobic Spore Forming Bacteria (ASFB) were close to each other in all treatments and control, these results indicated that all treatments were not effective on their growth, the source of ASFB is milk.
As samples of Labneh are kept in refrigerator, it is necessary evaluate Psychrotrophic Bacteria (PB) counts, it may release heat-resistant proteases and lipases, these enzymes will not be totally inactivated and may give rise to off-flavors. PB counts usually significantly decreased after 60 days at cold storage. Generally, PB in all treatments (CS and SFS) were lower than control, but there was not significantly (p<0.05) differences among all treatments, it was around 15×102 and could growth on medium after 4 weeks but was nearly 22×102 in control after 21 days. SFS recorded Total Viable Count (TVC) lower than CS whilst control samples were higher TVC than all treatments. These findings agreed with Abou Ayana and Gamal El Deen (2011). Concerning Lactic Acid Bacteria (LAB), the high levels of CS or SHS achieved LAB counts higher than low levels of CS or SHS, these results may due to increment of essential fatty acids or providing some important compounds for LAB. On the other hand, low levels of oilseeds recorded mold and yeasts counts lower than high levels of oilseeds. In general, mold and yeasts counts were quite limited; it may be that the privacy of milk goats, this property increased with feeding goats on rations containing oilseeds.
Organoleptic properties: Firstly, it has to be ascertained that addition of oil seeds or any additives to goat’s diets would not be detrimental to the sensory quality of dairy products. An important parameter to determine the quality and shelf life of Labneh is sensoric properties (Table 9). Significantly (p<0.05), HSFS (14% sun flower seeds) treatment gained the best organoleptic properties which gained higher score for appearance, body and texture, flavor and total score (the only shown) at different stages of storing followed by LSFS, HCS, LCS treatments then the control (without oil seeds).
| Table 9: | Total score organoleptic properties for Labneh made from treated goats milk |
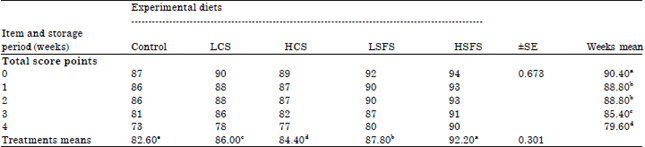 | |
Means within the same row or column within each category having different superscripts are significantly different at (p<0.05). Data of appearance, body and texture and flavour not shown | |
The change of milk fatty acid profile, affected Labneh texture, unsaturated fatty acid-rich milk produces less firm the samples. Supplementing maize silage-based diets with 3% sunflower oil led to moderate but noticeable sensory differences. Addition of 5% linseed oil to a maize silage-based diet was easily identified. Adding 2.5% fish oil induced sensory defects (Hurtaud et al., 2002). The most important observation was reduction of "goaty odour" and increment susceptibility of Labneh during the storage period contrary to the stored control sample. Generally, SFS treatments were better organoleptic properties than CS treatments. Certainly these results due to the improvement of milk composition particularly Fatty Acid (FA) profile of the milk which played an important role in improving the odour of Labneh and ameliorate its sensory properties. Some experiments have also shown the effects of forage and lipid supplements and their interactions on goat cheese flavor (Gaborit et al., 2004). Linseed oil or sunflower oil supplementation (5-6% of the ration) reduces the "goaty" taste in milk or fresh cheese, linked to the lower secretion of lipase and reduced post-milking lipolysis. However, Chilliard et al. (2002) reported that more bitter, piquant, oxidized or fishy flavours might occur, especially with the hay + linseed oil combination that maximizes milk C18: 3 concentration. Addition of some aromatic oils such as chamomile or thyme clearly improved the Labneh properties (Abou Ayana and Gamal El Deen, 2011).
Feed efficiency and economic evaluation: Data in Table 10 revealed that feed efficiency as kg 4% FCM per one kg DM, kg TDN and Kg DCP were higher with those fed oil seeds (CS and SFS) than those fed the control diet. The highest value was recorded with HCS diet followed by HSFS diet and then LCS and LSFS diets.
Regarding the economic evaluation, data indicated that feed cost to produced one kg 4% FCM was lower for goats received HCS, HSFS, LCS and LSFS diets than those fed the control diet, being 1.37, 1.38, 1.44, 1.42 and 1.55 L.E., respectively. Consequently, the economic evaluation was improved for goats fed HCS, HSFS, LCS and LSFS diets by 13.27, 11.95, 9.73 and 9.29%, respectively, than those fed control diet.
In this connection, El-Kholy et al. (2005) reported that replacing 10 and 20% of CFM by 5 and 10% of canola or sunflower seeds in growing buffalo diets improved the economical efficiency by 6, 13.7, 16.0 and 21.4%, respectively than those fed control diet. Awadalla et al. (2003) recommended that replacing 20% of the complete ration with 10% soybean seed or sunflower seeds in growing lambs diet’s resulted in superior economical efficiency compared with control diet. Aboul-Fotouh et al. (1999) found that the profit above feeding cost was higher with oil seeds diets than the control by 14.5% with cottonseeds diet and 29.8% with sunflower seeds diet for lactating buffaloes.
| Table 10: | Feed efficiency and economical evaluation of the experimental diets used in lactating Zaraibi goats |
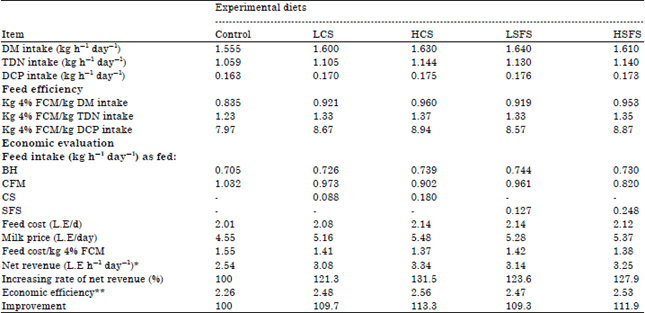 | |
The price list of one ton BH, CFM, CS and SFS were 650, 1500, 1700 and 1600 L.E, respectively and price of kg for row milk was 3.5 L.E. (Based on year 2012 prices), *Revenue (LE/h/d Money output - money input , **Efficiency: Money output/ money input | |
CONCLUSION
This study confirmed that milk and 4% Fat Corrected Milk (FCM) yields significantly (p<0.05) increased by 17.9 and 17.0% for Zaraibi goats fed oil seeds (canola, 5 and 10% or sunflower seeds, 7 and 14%) than for goats fed control diet, respectively. Milk fat, milk protein, unsaturated fatty acids and Conjugated linoleic acid (CLA) significantly (p<0.05) increased unlike lactose and saturated fatty acid significantly (p<0.05) decreased. Labneh made by milk with HSFS (14%) has the greatest organoleptic scores followed by LSFS, HCS, LCS treatments then the control,"goaty" odour in all treatments clearly decreased. VFA’s, acetic acid and the ratio of acetate to propionate increased, but NH3-N, propionic acid and butyric acid were decreased for goats fed canola or sunflower diets than the control. Worth noting, feed cost to produced one kg 4% FCM was lower for goats received HCS, HSFS, LCS and LSFS diets than those fed the control diet, being 1.37, 1.38, 1.44, 1.42 and 1.55 L.E., respectively. Consequently, the economic evaluation was improved for goats fed HCS, HSFS, LCS and LSFS diets by 13.27, 11.95, 9.73 and 9.29%, respectively, than those fed control diet. We recommend using canola or sunflower seeds as supplement for lactating goat rations at previous ratios.
REFERENCES
- Abou Ayana, I.A.A. and A.A. Gamal El Deen, 2011. Improvement of the properties of goat's milk labneh using some aromatic and vegetable oils. Int. J. Dairy Sci., 6: 112-123.
CrossRefDirect Link - Aboul-Fotouh, G.E., S.M. Allam and G.M. El-Gharhy, 1999. Effect of feeding whole cotton and sunflower oilseeds on lactation performance of buffaloes. Egypt. J. Anim. Prod., 36: 93-102.
Direct Link - Aldrich, C.G., N.R. Merchen, J.K. Drackley, G.C. Jr. Fahey and L.L. Berger, 1997. The effects of chemical treatment of whole canola seed on intake, nutrient digestibilities, milk production, and milk fatty acids of Holstein cows. J. Anim. Sci., 75: 512-521.
PubMedDirect Link - Basset, E.W. and W.J. Harper, 1958. Isolation and identification of acidic and neutral carbonyl compounds in different varieties of cheese. J. Dairy Sci., 41: 1206-1217.
CrossRefDirect Link - Beauchemin, K.A., S.M. McGinn, C. Benchaar and L. Holtshausen, 2009. Crushed sunflower, flax, or canola seeds in lactating dairy cow diets: Effects on methane production, rumen fermentation and milk production. J. Dairy Sci., 92: 2118-2127.
CrossRefDirect Link - Bernard, L., M. Bonnet, C. Leroux, K.J. Shingfield and Y. Chilliard, 2009. Effect of sunflower-seed oil and linseed oil on tissue lipid metabolism, gene expression and milk fatty acid secretion in Alpine goats fed maize silage-based diets. J. Dairy Sci., 92: 6083-6094.
CrossRefDirect Link - Chichlowski, M.W., J.W. Schroeder, C.S. Park, W.L. Keller D.E. Schimek, 2005. Altering the fatty acids in milk fat by including canola seed in dairy cattle diets. J. Dairy Sci., 88: 3084-3094.
CrossRefDirect Link - Chilliard, Y., A. Ferlay, J. Loor, J. Rouel and B. Martin, 2002. Trans and conjugated fatty acids in milk from cows and goats consuming pasture or receiving vegetable oils or seeds. Italian J. Anim. Sci., 1: 243-254.
CrossRefDirect Link - Chilliard, Y., A. Ferlay, R.M. Mansbridge and M. Doreau, 2000. Ruminant milk fat plasticity: Nutritional control of saturated, polyunsaturated, trans and conjugated fatty acids. Ann. Zootech., 49: 181-205.
CrossRefDirect Link - Chilliard, Y., A. Ferlay, J. Rouel and G. Lamberet, 2003. A review of nutritional and physiological factors affecting goat milk lipid synthesis and lipolysis. J. Dairy Sci., 86: 1751-1770.
CrossRefDirect Link - Chilliard, Y. and A. Ferlay, 2004. Dietary lipids and forages interactions on cow and goat milk fatty acid composition and sensory properties. Reprod. Nutr. Dev., 44: 467-492.
CrossRefPubMedDirect Link - Chouinard, P.Y., V. Girard and G.J. Brisson, 1997. Lactational response of cows to different concentrations of calcium salts of canola oil fatty acids with or without bicarbonates. J. Dairy Sci., 80: 1185-1193.
CrossRefDirect Link - El-Shafie, M.H. and T. El-Ashmawy, 2010. Performance of goats fed rations containing whole sunflower seeds. Egypt. J. Sheep Goat Sci., 5: 45-52.
Direct Link - Erwin, E.S., G.J. Marco and E.M. Emery, 1961. Volatile fatty acid analyses of blood and rumen fluid by gas chromatography. J. Dairy Sci., 44: 1768-1771.
CrossRefDirect Link - Glass, R.L., H.A. Troolin and R. Jenness, 1967. Comparative biochemical studies of milks-IV. Constituent fatty acids of milk fats. Comp. Biochem. Physiol., 22: 415-425.
CrossRefDirect Link - Grainger, C., R. Williams, T. Clarke, A.D.G. Wright and R.J. Eckard, 2010. Supplementation with whole cottonseed causes long-term reduction of methane emissions from lactating dairy cows offered a forage and cereal grain diet. J. Dairy Sci., 93: 2612-2619.
CrossRefDirect Link - Haenlein, G.F.W., 2004. Goat milk in human nutrition. Small Rumin. Res., 51: 155-163.
CrossRefDirect Link - Herdt, T.H., 1988. Fuel homeostasis in the ruminant. Vet. Clin. North Am. Food Anim. Pract., 4: 213-231.
PubMedDirect Link - Huth, P.J., D.B. Di Rienzo and G.D. Miller, 2006. Major scientific advances with dairy foods in nutrition and health. J. Dairy Sci., 89: 1207-1221.
CrossRefDirect Link - Jacobs, A.A.A., J. van Baal, M.A. Smits, H.Z.H. Taweel, A.M. van Vuuren and J. Dijkstra, 2011. Effects of feeding rapeseed oil, soybean oil, or linseed oil on stearoyl-CoA desaturase expression in the mammary gland of dairy cows. J. Dairy Sci., 94: 874-887.
CrossRefDirect Link - Johnson, K.A., R.L. Kincaid, H.H. Westberg, C.T. Gaskins, B.K. Lamb and J.D. Cronrath, 2002. The effect of oilseeds in diets of lactating cows on milk production and methane emissions. J. Dairy Sci., 85: 1509-1515.
CrossRefDirect Link - Kadegowda, A.K.G., M. Bionaz, L.S. Piperova, R.A. Erdman and J.J. Loor, 2009. Peroxisome proliferator-activated receptor-γ activation and long-chain fatty acids alter lipogenic gene networks in bovine mammary epithelial cells to various extents. J. Dairy Sci., 92: 4276-4289.
CrossRefDirect Link - Marin, A.L.M., M.P. Hernandez, L.M.P. Alba, D.C. Pardo, A.I.G. Sigler and G.G. Castro, 2013. Fat addition in the diet of dairy ruminants and its effects on productive parameters. Revista Colombiana Ciencias Pecuarias, 26: 69-78.
Direct Link - McNamara, R.K., R. Jandacek, T. Rider, P. Tso, C.G. Hahn, N.M. Richtand and K.E. Stanford, 2007. Abnormalities in the fatty acid composition of the postmortem orbitofrontal cortex of schizophrenic patients: Gender differences and partial normalization with antipsychotic medications. Schizophr. Res., 91: 37-50.
CrossRefDirect Link - Onetti, S.G., R.D. Shaver, M.A. McGuire, D.L. Palmquist and R.R. Grummer, 2002. Effect of supplemental tallow on performance of dairy cows fed diets with different corn silage: Alfalfa silage ratios. J. Dairy Sci., 85: 632-641.
CrossRefDirect Link - Palmquist, D.L. and T.C. Jenkins, 1980. Fat in lactation rations: Review. J. Dairy Sci., 63: 1-14.
CrossRefPubMedDirect Link - Patton, C.J. and S.R. Crouch, 1977. Spectrophotometric and kinetics investigation of the Berthelot reaction for the determination of ammonia. Anal. Chem., 49: 464-469.
CrossRefDirect Link - Petit, H.V., 2003. Digestion, milk production, milk composition and blood composition of dairy cows fed formaldehyde treated flaxseed or sunflower seed. J. Dairy Sci., 86: 2637-2646.
CrossRefDirect Link - Luna, P., A. Bach, M. Juarez and M.A. de la Fuente, 2008. Influence of diets rich in flax seed and sunflower oil on the fatty acid composition of ewes' milk fat especially on the level of conjugated linoleic acid, n-3 and n-6 fatty acids. Int. Dairy J., 18: 99-107.
CrossRefDirect Link - Piperova, L.S., B.B. Teter, I.J. Bruckental, S.E. Sampugna and M.P. Mills et al., 2000. Mammary lipogenic enzyme activity, trans fatty acids and conjugated linoleic acids are altered in lactating dairy cows fed a milk fat-depressing diet. J. Nutr., 130: 2568-2574.
PubMedDirect Link - Salem, M.M.E., Mona A.M. Abd El-Gawad, Fatma A.M. Hassan and B.A. Effat, 2007. The use of synbiotics for production of functional low fat Labneh. Pol. J. Food Nutr. Sci., 16: 151-159.
Direct Link - Schauff, D.J. and J.H. Clark, 1992. Effects of feeding diets containing calcium salts of long-chain fatty acids to lactating dairy cows. J. Dairy Sci., 75: 2990-3002.
CrossRefDirect Link - Sinclair, L.A., W.M.P.B. Weerasinghe, R.C. Wilkinson, M.J. de Veth and D.E. Bauman, 2010. A supplement containing trans-10, cis-12 conjugated linoleic acid reduces milk fat yield but does not alter organ weight or body fat deposition in lactating ewes. J. Nutr., 140: 1949-1955.
PubMedDirect Link - Teres, S., G. Barcelo-Coblijn, M. Benet, R. Alvarez, R. Bressani, J.E. Halver and P.V. Escriba, 2008. Oleic acid content is responsible for the reduction in blood pressure induced by olive oil. Proc. Natl. Acad. Sci. United States Am., 105: 13811-13816.
CrossRefDirect Link - Toral, P.G., G. Hervas, P. Gomez-Cortes, P. Frutos, M. Juarez and M.A. de la Fuente, 2010. Milk fatty acid profile and dairy sheep performance in response to diet supplementation with sunflower oil plus incremental levels of marine algae. J. Dairy Sci., 93: 1655-1667.
CrossRefDirect Link - Warner, A.C., 1964. Production of volatile fatty acids in the rumen: Methods of measurement. Nutr. Abstr. Rev., 34: 339-352.
PubMed - Zhang, R.H., A.F. Mustafa and X. Zhao, 2006. Effects of feeding oilseeds rich in linoleic and linolenic fatty acids to lactating ewes on cheese yield and on fatty acid composition of milk and cheese. Anim. Feed Sci. Technol., 127: 220-223.
CrossRefDirect Link