
Noura El-Ahmady El-Naggar
Department of Bioprocess Development, Genetic Engineering and Biotechnology Research Institute, City of Scientific Research and Technological Applications, Alexandria, New Borg El-Arab City, Egypt
Sahar Deraz
Department of Protein Research, Genetic Engineering and Biotechnology Research Institute, City of Scientific Research and Technological Applications, Alexandria, New Borg El-Arab City, Egypt
Ashraf Khalil
Department of Protein Research, Genetic Engineering and Biotechnology Research Institute, City of Scientific Research and Technological Applications, Alexandria, New Borg El-Arab City, Egypt







ABSTRACT
All over the world, research on bioethanol production has grown with increasing of energy needs and it has become a research area of great interest to many governments, academic groups and companies. Ethanol which is presently the most common renewable fuel, can be produced biologically from a variety of feedstocks and wastes. Due to advances in agriculture and biotechnology one can envision inexpensive production of ethanol based on lignocellulosic biomass. This review summarizes various processes involved in lignocellulosic-derived biofuel bioconversion. Several methods of pretreatment of lignocelluloses are discussed. Characteristics of enzymes and important factors in enzymatic hydrolysis of the cellulose and hemicellulose are reviewed. Different strategies are then described and illustrated in a simpler form for enzymatic hydrolysis and fermentation, including separate enzymatic hydrolysis and fermentation, simultaneous saccharification and fermentation, simultaneous saccharification and co-fermentation and consolidated bioprocessing. Furthermore, recent trends, major challenges and perspective of future development are highlighted.
PDF Abstract XML References Citation
Received: December 20, 2013;
Accepted: January 10, 2014;
Published: May 06, 2019
How to cite this article
Noura El-Ahmady El-Naggar, Sahar Deraz and Ashraf Khalil, 2014. Bioethanol Production from Lignocellulosic Feedstocks Based on Enzymatic Hydrolysis:
Current Status and Recent Developments. Biotechnology, 13: 1-21.
DOI: 10.3923/biotech.2014.1.21
URL: https://scialert.net/abstract/?doi=biotech.2014.1.21
DOI: 10.3923/biotech.2014.1.21
URL: https://scialert.net/abstract/?doi=biotech.2014.1.21
INTRODUCTION
One of the biggest problems facing the world is the energy crisis. The price of oil dramatically escalated in 1973, creating an oil crisis. Energy consumption has increased steadily as the world population has grown and more countries have become industrialized. The fossil fuels, including crude oil, coal and natural gas are the major resources to meet the increased energy demands. As widely believed, the fossil energies will be exhausted in the near future (Wyman, 2001). The world currently consumes 30 billion barrels of oil per year; Colin (2003) estimates that oil reserves will become scarce by the 2050s. Because petroleum is a nonrenewable resource, there is an urgent need to seek alternative energy sources that are inexhaustible.
In recent decades, an international awareness of the increasing CO2 concentration levels in the atmosphere and concern for global warming led to the formulation of the Kyoto Protocol in 1997 which has led many countries to make the commitment to decrease the emission of CO2. One way of decreasing CO2 emissions could be substitution of fossil fuels with renewable energy sources.
The development of alternative fuel and energy sources has thus become a worldwide research priority in recent years. Biofuels are fuels produced from biomass. These fuels are usually in the form of bioalcohols, biodiesel, biogas and other chemicals produced from biomass. The two main biofuels are biodiesel and bioethanol. Among those, bioethanol produced by bioconversion of lignocellosic biomass is being seen as one of the most promising alternative biofuels. The choice of lignocellulose to bioethanol conversion should be decided on the basis of overall economics (lowest cost), environment (lowest pollutants) and energy (higher efficiency), i.e., comprehensive process development and optimization are still required to make the process economically viable. The increasing petroleum price and negative impact of fossil fuels on the environment are encouraging the use of lignocellulosic materials to help meet energy needs (Di Nasso et al., 2011).
There are many advantages of biofuels over fossil fuels that make the alternative fuel source an attractive option now and in the future. The main advantage of biofuels is that they are considered ‘carbon neutral’ by some people. This is because the carbon dioxide released during the combustion of biofuels is equal to the amount that assimilated during photosynthesis (Kheshgi et al., 2000) resulting in no net increase to CO2 levels. Therefore, they don’t contribute to global warming. Consumption of biofuels releases no sulfur and has much lower particulate and toxic emissions, particularly when compared with other liquid transportation fuels (Scott and Wyman, 2004). Bioethanol production can provide an attractive route to dispose of problematic lignocellosic wastes such as stalks, stovers and leaves of agricultural crops. Biofuels are renewable energy source (made from organic materials and even organic waste, there is practically an infinite amount of biofuels available), inexpensive to produce and reduce dependence on foreign oils.
Ethanol is one of the most promising biofuels that can be used to replace gasoline for tomorrow’s transportation vehicles. Fuel ethanol is mainly used as an oxygenated fuel additive. The higher octane number of the fuel mixture, when it contains ethanol, reduces the need for toxic, octane-enhancing additives such as methyl tertiary butyl ether. Due to the oxygen in ethanol molecules, there is also a reduction of carbon monoxide emission and non-combusted hydrocarbons (Hu et al., 2008).
Feedstocks for bioethanol production
Generations of bioethanol: There are three “generations” of bioethanol: First-generation or conventional bioethanol is bioethanol produced from sugar substances, such as sugarcane juice, sugar beets and molasses, as well as starch-based materials such as wheat, corn, potatoes or cassava. Sucrose can be fermented directly to ethanol, but starch must be hydrolyzed to glucose before it can be fermented by yeast, generally by Saccharomyces cerevisiae (Sameera et al., 2011). Even though the production of ethanol from starch biomass materials represents the most convenient and technically advanced option for bioenergy, such practices would result in severe competition between energy and food supplies. The raw materials account for 40-70% of the total ethanol production costs based on current sugar- or starch-containing feedstocks. Reduction of the production cost would increase the competitiveness of ethanol with fossil fuels. Lignocellulosic biomass is believed to be less expensive and more plentiful than either starch- or sucrose-containing feedstocks (Hu et al., 2008).
Second generation bioethanol is bioethanol produced from sustainable feedstock that cannot be used directly for food production. Examples of wastes that could potentially be converted into bioethanol include perennial grasses, agricultural wastes and forest residues. The conversion of these potential sources of feedstock for bioethanol offers the double benefit of a reduction in global waste and the generation of valuable transportation fuels (Kaylen et al., 2000).
Third generation bioethanol is produced via., direct cellulose fermentation (cellulase production, substrate hydrolysis and fermentation are accomplished in a single process step by cellulolytic microorganisms).
Lignocelluloses: Approximately 2x1011 tons of lignocellulosic biomass is produced every year and organic agricultural and forestry by-products are annually renewable, available in abundance and of limited value (Chen et al., 2007). By-products produced from the cultivation of corn (corn stover), wheat (wheat straw), rice (rice straw), sorghum (sorghum stalks), barley (barley straw) and sugarcane (sugarcane bagasse) are the major sources of agro-based lignocellulosics which contain high contents of cellulose and hemicellulose, as described in Table 1 (Sun and Cheng, 2002). However, the excess lignocellulosic waste is often disposed of by biomass burning, causing an environmental pollution problem through smoke and development of CO2 gas which is not restricted to developing countries, but is considered a global phenomenon (Levine, 1996). Burning also causes almost complete losses of soil N, P, K and S (Dobermann and Fairhurst, 2002). At the same time, the cellulose and hemicellulose could provide sugar for bioconversion to biofuels (Malherbe and Cloete, 2003).
It is an important issue to deal with the agricultural waste both for the comprehensive utilization of lignocellulosic resources and for the prevention of environmental pollution. One of the different approaches to overcome this problem is the biological conversion of agricultural waste to biofuels.
| Table 1: | Composition of some common agricultural and forestry residues |
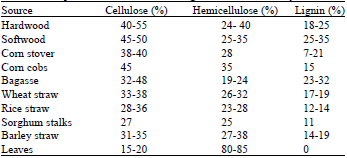 | |
| Adapted from Kumar et al. (2009) | |
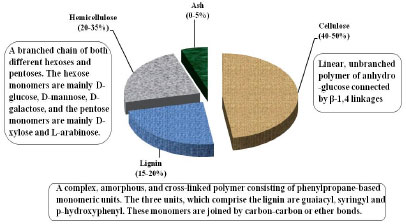 | |
| Fig. 1: | Constituents of lignocellulosic biomass |
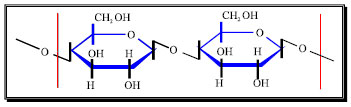 | |
| Fig. 2: | Cellulose molecular structure |
Production of ethanol from renewable lignocellulosic sources is a promising means to decrease the accumulation of greenhouse gas and alleviate pressure on fossil fuel shortage (Galbe and Zacchi, 2002).
The main constituents of lignocellulosic biomass are cellulose, hemicellulose and lignin. Minor components include ash and extractives such as waxes. On a dry weight basis, lignocelluloses contain 40-50% cellulose, 20-35% hemicellulose and 15-20% lignin (Fig. 1). Together, these components represent approximately 90% of the dry weight of most plant material (Brown, 2003).
The cellulose chain is a linear, unbranched polymer of glucose monomers linked by β-1,4-glucosidic bonds (Fig. 2). The conformation of cellulose, as well as its close association with lignin and hemicellulose makes cellulose very resistant to hydrolysis.
Lignocelluloses-to-bioethanol processes: There are two basic approaches to degrading biomass to sugars: Enzymatic hydrolysis and dilute acid hydrolysis. Compared to dilute acid hydrolysis, enzymatic approach is promising because it can achieve high sugar yields and eliminate the need for large quantities of chemicals and the formation of inhibitory by-products during dilute acid hydrolysis (Tran and Chambers, 1986).
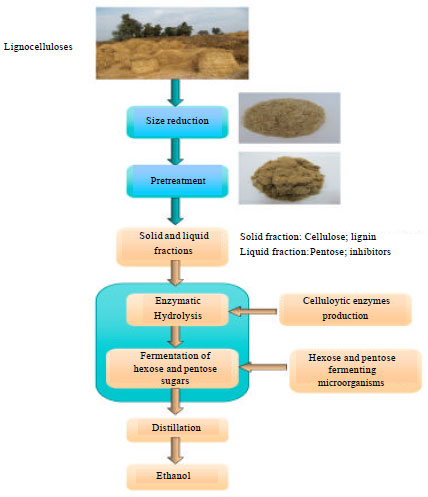
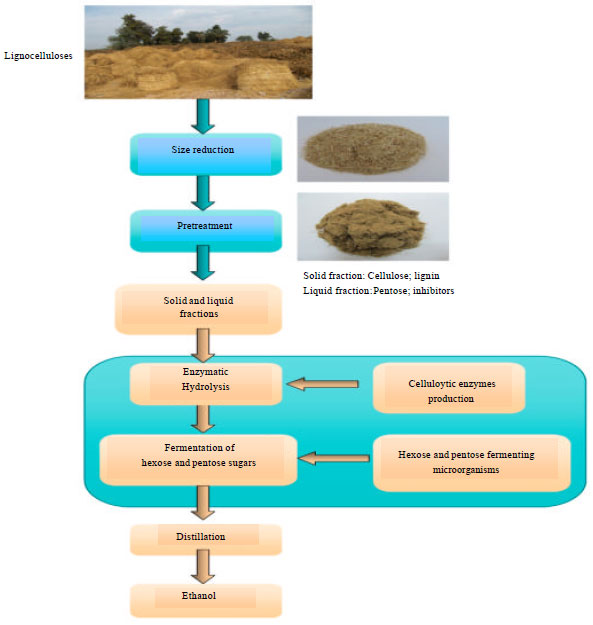
The production of ethanol from lignocellulosic material requires the release of monomeric sugar units constituting cellulose and hemicellulose. Based on the specific lignocellulosic structure, the bioconversions of lignocellulosic materials to bioethanol normally require multi-step processes. There are five stages to produce ethanol using a biological approach (Zhu et al., 2009) (1) A "pretreatment" phase, to make the lignocellulosic material such as wood or straw amenable to hydrolysis, (2) Cellulose hydrolysis, to break down the molecules into sugars, (3) Separation of the sugar solution from the residual materials, notably lignin, (4) Microbial fermentation of the sugar solution and (5) Distillation to produce roughly 95% pure alcohol. An optional further step would be dehydration by molecular sieves to bring the ethanol concentration to over 99.5%.
Pretreatment of lignocellulose for enzymatic hydrolysis: The combination of hemicellulose and lignin provide a protective sheath around cellulose which must be modified or removed before efficient hydrolysis of cellulose can occur. Additionally, the higher order crystalline structure of cellulose is an obstacle to hydrolysis (Eriksson et al., 1993). To make cellulose and hemicellulose accessible for enzymatic digestibility, a number of pretreatments should be carried out. These pretreatments are summarized in Table 2. Pretreatment is the first step required to fractionate lignocellulosic materials into the major plant components of lignin, cellulose and hemicellulose and/or modify the pores in the material to allow the enzymes to penetrate into the fibers to render them amenable to enzymatic hydrolysis (Galbe and Zacchi, 2002; Mosier et al., 2005). The rice straw before and after pretreatment is depicted in Fig. 3.
The mechanisms by which pretreatments improve the digestibility of lignocellulose are however, not well understood (Brown, 2003).
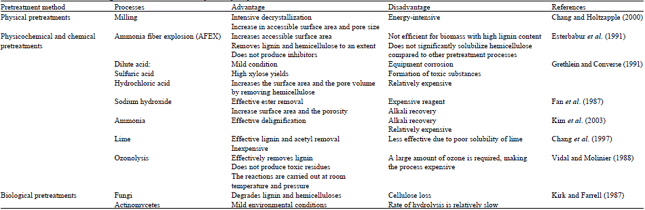
| Table 2: | Pretreatment methods of lignocellulosic materials for enzymatic hydrolysis |
 | |
 | |
| Fig. 3(a-b): | Rice straw before and after pretreatment (a) Rice straw and (b) Pretreated rice straw |
A successful pretreatment must meet the following requirements: (1) Improve formation of sugars or the ability to subsequently form sugars by hydrolysis, (2) Avoid the degradation or loss of the pentose (hemicellulose) fractions to completely use the carbohydrates, (3) Avoid the formation of byproducts inhibitory to the subsequent hydrolysis and fermentation processes and (4) Be cost effective (Sun and Cheng, 2005).
Several methods have been introduced for pretreatment of lignocellulosic materials prior to enzymatic hydrolysis. The pretreatment methods can be classified into “Physical pretreatment” such as milling and irradiation (Wyman, 1996), “Physico-chemical pretreatment” such as steam explosion or autohydrolysis, ammonia fiber explosion (AFEX) (Ohgren et al., 2005), “Chemical pretreatment” including ozonolysis, dilute-acid pretreatment and alkaline pretreatment (Berlin et al., 2006) and “Biological pretreatment” (Fan et al., 1982; Wyman, 1996).
Physical pretreatment: Physical pretreatment can increase the accessible surface area and size of pores and decrease the crystallinity and degrees of polymerization of cellulose. Different types of physical processes such as milling and irradiation can be used to improve the enzymatic hydrolysis of lignocellulosic waste materials.
Milling: Milling (cutting the lignocellulosic biomass into smaller pieces) can be employed to alter the inherent ultrastructure of lignocelluloses and degree of crystallinity and consequently make it more amenable to cellulase. Milling and size reduction have been applied prior to enzymatic hydrolysis, or even other pretreatment processes with dilute acid, steam or ammonia, on several lignocellulosic waste materials (Mais et al., 2002). Milling can improve enzymatic degradation of lignocellulosic materials toward ethanol by reducing the size of the materials (Chang and Holtzapple, 2000; Zeng et al., 2007), the reduction in particle size leads to an increase of available specific surface and a reduction of the degree of polymerization (Palmowski and Muller, 1999). Although, decrystallization has been reported to be less important than lignin removal on sugar yields, decreasing crystallinity significantly increased the initial hydrolysis rate and to some extent, the ultimate biomass conversion (Chang and Holtzapple, 2000).
Irradiation: Irradiation by e.g., gamma rays and microwaves can improve enzymatic hydrolysis of lignocelluloses. The combination of the irradiation and other methods such as acid treatment can further accelerate enzymatic hydrolysis (Mamar and Hadjadj, 1990). Irradiation has enhanced enzymatic degradation of cellulose into glucose. Kumakura and Kaetsu (1983) studied the effect of irradiation for pretreatment of bagasse prior to its enzymatic hydrolysis. The pretreated bagasse resulted in doubling the yield of glucose by the hydrolysis compared to the untreated one. The cellulose component of the lignocellulose materials can be degraded by irradiation to fragile fibers and low molecular weight oligosaccharides and even cellobiose (Kumakura and Kaetsu, 1983).
Chemical pretreatment
Dilute acid pretreatment: Treatment of lignocellulosic materials with acid at a high temperature can efficiently improve the enzymatic hydrolysis. Sulfuric acid is the most applied acid while other acids such as HCl and nitric acid also have been reported (Taherzadeh and Karimi, 2007a). The acid pretreatment can operate either under a high temperature (e.g., 180°C for 5 min) and low acid concentration (dilute-acid pretreatment) or under a low temperature (e.g., 120°C for 30-90 min) and high acid concentration (concentrated-acid pretreatment). Dilute-acid hydrolysis is probably the most commonly applied method among the chemical pretreatment methods. It can be used either as a pretreatment of lignocellulose for enzymatic hydrolysis, or as the actual method of hydrolyzing to fermentable sugars. Dilute acid pretreatment increases the surface area and the pore volume by removing hemicellulose. Actually, chemical treatments have a dual effect on biomass crystallinity: (1) They remove amorphous lignin and hemicellulose components to increase biomass crystallinity and (2) They loosen the highly packed crystalline structure through swelling to decrease crystallinity (Gharpuray et al., 1983).
The main advantage of dilute-acid pretreatment is the possibility to recover a high portion (e.g., 90%) of the hemicellulose sugars. The hemicellulose, mainly xylan or mannan, accounts for up to a third of the total carbohydrate in many lignocellulosic materials. Thus, hemicellulose recovery (e.g., achieving high xylan to xylose conversion yields) can have a highly positive effect on the overall process economics of ethanol production from lignocellulosic material (Sun and Cheng, 2002). On the other hand, a main disadvantage of this pretreatment method is the necessity of neutralization of pH for the downstream enzymatic hydrolysis. Furthermore, different chemical inhibitors might be produced during the acid pretreatment which reduces cellulase activity and therefore, water wash is necessary for the pretreated biomass before enzymatic hydrolysis (Sun and Cheng, 2002). Dilute-acid pretreatment is not effective in dissolving lignin, but it can disrupt lignin and increases the cellulose’s susceptibility to enzymatic hydrolysis (Yang and Wyman, 2004).
Alkaline pretreatment: Alkaline pretreatment refers to the application of alkaline solutions such as NaOH, Ca(OH)2 (lime) or ammonia to remove lignin and a part of the hemicellulose and efficiently increase the accessibility of enzyme to the cellulose. Pretreatment can be performed at low temperatures but with a relatively long time and high concentration of the base. The mechanism of alkali pretreatment is believed to be saponification (breaking) of intermolecular ester bonds between lignin, hemicellulose and cellulose and avoiding fragmentation of the hemicellulose polymers (Gaspar et al., 2007). After alkaline pretreatment, the porosity of the material is increased due to the extensive swelling facilitated by removal of the crosslinks (Tarkow and Feist, 1969). Dilute NaOH pretreatment causes swelling, leading to an increase in internal surface area and the porosity of the material is increased, a decrease in the degree of polymerization and crystallinity and disruption of lignin structure (Fan et al., 1987). In comparison with dilute acid pretreatment, the high cost of caustic soda is the main obstacle to its large scale implementation. Therefore, aqueous ammonia and lime (calcium hydroxide) pretreatments have been developed to reduce the cost of chemicals. These two kinds of alkali are effective in removing lignin and acetyl groups in biomass. Lime is a promising chemical because it is inexpensive, safe and can be recovered by carbonating wash water (Chang et al., 1997).
Ozonolysis pretreatment: Pretreatment of lignocellulosic materials can be performed by treatment with ozone, referred to as “ozonolysis” pretreatment. This method can effectively degrade lignin and part of hemicellulose. The pretreatment is usually carried out at room temperature and does not lead to inhibitory compounds (Vidal and Molinier, 1988). However, ozonolysis might be expensive since a large amount of ozone is required (Sun and Cheng, 2002). The main parameters in ozonolysis pretreatment are moisture content of the sample, particle size and ozone concentration in the gas flow. Among these parameters, an essential factor is the percentage of water in the feed and it has the most significant effect on the solubilization. The optimum water content was found to be around 30%, corresponding to the saturation point of the fibers. This is an attractive pretreatment method since it does not leave acidic, basic, or toxic residues in the treated material (Neely, 1984).
Physico-chemical pretreatment: Pretreatments that combine both chemical and physical processes are referred to as physicochemical processes (Chandra et al., 2007).
Ammonia fiber explosion (AFEX): AFEX is one of the alkaline physico-chemical pretreatment processes. Here the biomass is exposed to liquid ammonia at relatively high temperature (e.g., 90-100°C) for a period of e.g., 30 min, followed by immediate reduction of pressure. The effective parameters in the AFEX process are ammonia loading, temperature, water loading, blowdown pressure and time (Holtzapple et al., 1991). The AFEX process can either modify or effectively reduce the lignin fraction of the lignocellulosic materials while the hemicellulose and cellulose fractions may remain intact. One of the major advantages of AFEX pretreatment is no formation of some types of inhibitory by-products which are produced during the other pretreatment methods, such as furans in dilute-acid and steam explosion pretreatment. However, part of phenolic fragments of lignin and other cell wall extractives may remain on the cellulosic surface. Therefore, washing with water might be necessary to remove part of these inhibitory components, although increasing the amount of wastewater from the process (Chundawat et al., 2007). However, there are some disadvantages in using the AFEX process compared to some other processes. AFEX is more effective on the biomass that contains less lignin and the AFEX pretreatment does not significantly solubilize hemicellulose compared to other pretreatment processes such as dilute-acid pretreatment.
Biological pretreatment: Microorganisms can also be used to treat the lignocelluloses and enhance enzymatic hydrolysis. The applied microorganisms usually degrade lignin and hemicellulose but very little part of cellulose, since cellulose is more resistance than the other parts of lignocelluloses to the biological attack. White-rot fungi are among the most effective microorganisms for biological pretreatment of lignocelluloses (Sun and Cheng, 2002). Some bacteria such as Bacillus circulans (Kurakake et al., 2007) can also be used for biological pretreatment of lignocellulosic materials. Low energy requirement and mild environmental conditions are the main advantages of biological pretreatment. However, this method is relatively slow (Hahn-Hagerdal et al., 2006).
Detoxification of lignocellulose-derived inhibitor compounds: The major drawback of some pretreatment methods is the formation of different types of potential microbial inhibitors (Chandel et al., 2009, 2010). The amounts and types of inhibitory compounds vary strongly between different raw materials and also depend on the pretreatment method. These inhibitors can be divided into three major groups, (1) Organic acids (acetic, formic and levulinic acids), (2) Furan derivatives [furfural and 5-hydroxymethylfurfural (5-HMF)] and (3) Phenolic compounds (Chandel et al., 2010). These inhibitors may not affect the enzymatic hydrolyses, but they usually inhibit the microbial growth and fermentation which results in less yield and productivity of ethanol (Taherzadeh and Karimi, 2007a). The ethanologenic microorganisms have ability to degrade some of the inhibitors. Progress has been made to achieve higher levels of sugars by diminishing the overall impact of fermentative inhibitors which in-turn improves the fermentability of lignocellulosic hydrolysates (Alriksson et al., 2011; Parawira and Tekere, 2011). The ion exchange resins, active charcoal, enzymatic detoxification using laccase and overliming with calcium hydroxide are among selective detoxification (i.e., removal of inhibitory compounds) strategies which have been investigated in the past (Jurado et al., 2009). Treatment with the soft-rot fungus Trichoderma reesei and other microorganisms to degrade inhibitors in a hemicellulase hydrolysate has also been proposed (Fonseca et al., 2011; Yu et al., 2011). Biodetoxification methods provides high yields, uses limited waste water and has low energy costs, but its slow rate prohibits its practical use.
Enzymatic hydrolysis: Enzymatic hydrolysis has attracted increasing attention as an alternative to concentrated acid hydrolysis because the process is highly specific, can be performed under mild reaction conditions (pH around 5 and temperature less than 50°C) with lower energy consumption and lower environmental impact. In addition, it does not present corrosion problems and gives high yield of pure glucose with low formation of by-products that are favorable for the subsequent hydrolysate use in fermentation processes. The cellulolytic enzymes are either secreted into the substrate or attached to the cell wall of the microorganism. The former system is called noncomplexed and the latter complexed cellulase system. Noncomplexed cellulase systems are mostly found in filamentous fungi and actinomycete bacteria, because they can penetrate the lignocellulosic material with their hyphal extensions. The enzymes of noncomplexed cellulase systems are then just released into the substrate and the free enzymes start hydrolyzing the cellulose. The glucose and cellodextrins of a length of maximal four glucose molecules are taken up by the microorganism and either used directly or cleaved further via intracellular hydrolases. Organisms that produce noncomplexed cellulase systems are most often used in the industrial production of cellulolytic enzymes, because the secreted enzymes can easily be harvested (Lynd et al., 2002). Complexed cellulase systems are mainly found in anaerobic bacteria and anaerobic fungi that cannot penetrate lignocellulosic material. The different cellulolytic enzymes of these bacteria form a stable “complex”, called the cellulosome.
Enzymatic degradation of cellulose to glucose is generally accomplished by synergistic action of at least three major classes of enzymes: Endo-glucanases, exoglucanases and β-glucosidases. These enzymes are usually called together cellulase or cellulolytic enzymes (Wyman, 1996).
Cellulases: Cellulases responsible for the hydrolysis of cellulose are composed of three major enzymes (Rabinovich et al., 2002): Endoglucanases or carboxymethyl cellulases (endo-1,4-β-glucanase; EC 3.2.1.4), are proposed to initiate attack randomly at multiple internal sites in the amorphous regions of cellulose fiber opening-up sites and create free chain-ends for subsequent attack by the cellobiohydrolases (Wood, 1991). Exoglucanases or cellobiohydrolases (Exoglucanase; EC 3.2.1.91), is the major component of cellulase system accounting for 40 to 70% of the total cellulase proteins and can hydrolyze highly crystalline cellulose (Esterbabur et al., 1991). Exoglucanases act in a processive manner on the reducing or nonreducing ends of cellulose polysaccharide chains, liberating either glucose or cellobiose as major products. Exoglucanases can also act on microcrystalline cellulose, presumably peeling cellulose chains from the microcrystalline structure. β-glucosidases (EC 3.2.1.21), splits cellobiose into two glucose units (Lynd et al., 2002). This enzyme is not a cellulase, but its action is very important to complete depolymerization of cellulose to glucose.
Synergism in cellulose hydrolysis: The application of cellulolytic enzymes in degrading lignocellulosic materials needs synergistic actions of by endoglucanase, exoglucanase, or cellobiohydrolase and β-glucosidase (Rabinovich et al., 2002; Zhang et al., 2006). In enzymatic reactions, synergism occurs when the combined activity of two enzymes exceeds the sum of the individual enzyme activities. In the hydrolysis of filter study, a one to one mixture of exoglucanase and endoglucanase from T. reesei resulted in 23% (w/w) conversion, whereas the individual conversion was 6 and 8% using exoglucanase and endoglucanase, respectively (Gama et al., 1998).
Hemicellulases: Hemicelluloses are degraded more easily than cellulose because of a lower degree of polymerization and their amorphous nature. Since, hemicellulose contains different sugar units, the hemicellulolytic enzymes are more complex and include endo-1,4-β-D-xylanases, exo-1,4-β-D-xylosidases, endo-1,4-β-D-mannanases, β-mannosidases, galactanases, α-glucuronidases, endo-arabinases, α-L-arabinofuranosidases and α-galactosidases (Jorgensen et al., 2003). Hemicellulases also include the accessory enzymes, such as ferulic acid esterases, coumaric acid esterases and acetylxylan esterase which are necessary for hydrolyzing various substituted xylans (Saha, 2003). Xylanases are one of the major hemicellulases which hydrolyse the β-1,4 bond in the xylan backbone yielding short xylooligomers which are further hydrolysed into single xylose units by β-xylosidase. β-mannanases hydrolyse mannan-based hemicellulose and liberate short β-1, 4-manno-oligomers which can further be hydrolysed to mannose by β-mannosidases. Hemicellulolytic esterases include acetyl esterases which hydrolyse the acetyl substitutions on xylose moieties and feruloyl esterase which hydrolyse the ester bond between the arabinose substitutions and ferulic acid. Feruloyl esterase aids the release of hemicellulose from lignin and renders the free polysaccharide product more amenable to degradation by the other hemicellulases (Prates et al., 2001).
Ligninases: Lignin is very hard to be degraded compared with other celluloses and hemicelluloses in most agriculture wastes. The breakdown of lignin is not well understood; however, most literatures proved correlation between the ability to degrade lignin and the production of extracellular phenolases, such as laccase, peroxidase and tyrosinase which oxidize phenolic compounds, e.g., gallic and tannic acids. Fungi breakdown lignin aerobically through the use of family of extracellular enzymes collectively termed lignases. Phenol oxidase (laccase), lignin peroxidase and manganese peroxidase are widely considered to play a key role in lignin degradation (Krause et al., 2003; Malherbe and Cloete, 2003).
Enzyme cost: Cellulase production was the most expensive step during ethanol production from cellulosic biomass, in that it accounted for approximately 50% of the total hydrolysis process cost. Significant cost reduction is required in order to enhance the commercial viability of cellulase production technology (Howard et al., 2004). However the cost of enzymes has decreased over the last twenty years but is still considered to be very high. Much research efforts have been focused on lowering the cost of enzymes.
The following strategies can be applied for reducing the cellulase production cost: (1) Strain improvement of existing industrial organisms (by mutagenesis, by protoplast fusion technology, by genetic engineering and recombinant DNA technology), (2) Screening for overproducing strains with novel enzymes, (3) Using mixed cultures or co cultures of organisms. In order to enhance cellulase activity, mixed cultures are generally used. Application of mixed cultures is an alternative tool to overcome feedback inhibition and catabolite repression, (4) Using cheap, renewable and easily available cellulosic wastes. Due to the high production cost of pure cellulose such as Avicel, the various lignocellulosic raw materials is proved to be an best alternative for cellulase production such as wheat straw, rice straw, corn stover etc., (5) By using fed batch system and (6) Applying recent trends in solid state fermentation, solid state fermentation gives a concentrated product which is advantageous in cellulase use for biomass hydrolysis.
Microorganisms and their cellulolytic enzymes: Several species of bacteria such as Cellumonas, Bacillus, Thermomonospora, Erwinia, Microbispora and Streptomyces (El-Naggar et al., 2011; El-Naggar and Abdelwahed, 2012) and fungi such as Tricoderma, Penicillium, Fusarium, Humicola spp. and Aspergillus fumigates (Sherief et al., 2010) are able to produce cellulases and hemicellulases. Among the cellulases produced by different microorganisms, cellulases of Trichoderma reesei or T. viride and its mutants have been the most broadly studied and best characterized (Esterbabur et al., 1991). A full complement production of cellulase, stability under the enzymatic hydrolysis conditions and resistance of the enzyme to chemical inhibitors are the advantages of the cellulase produced by Trichoderma. The main disadvantage of Trichoderma cellulase is low activity of β-glucosidases. On the other hand, Aspergilli are very efficient β-glucosidase producers. In several studies, Trichoderma cellulase was supplemented with extra β-glucosidases and showed good improvement (Krishna et al., 2001; Itoh et al., 2003).
Factors affecting the enzymatic hydrolysis of lignocellulosic materials: The enzymatic hydrolysis of lignocellulose is affected by many factors. The limiting factors have been traditionally divided into two groups: Those related to lignocellulose structural features and those related to the mechanism and interactions of the cellulolytic enzymes. Generally, structural features can be categorized into two groups: Physical and chemical. Physical structural features include cellulose crystallinity, degree of cellulose polymerization, pore volume, accessible surface area and particle size. Chemical structural features include the contents of lignin, hemicellulose and acetyl groups. Table 3 summarizes the relationship between structural features and biomass digestibility.
Substrate-related factors
Lignin content: Lignin plays a significant role in the rate and extent of lignocellulose hydrolysis. Literature results have all shown that cellulose digestibility enhances with increasing lignin removal (Draude et al., 2001).
| Table 3: | Summary of relationship between structural features and digestibility (Zhu et al., 2008) |
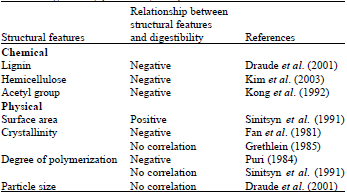 | |
The major inhibitory role of lignin has been attributed to nonspecific adsorption of enzyme to lignin (Sewalt et al., 1997) thus preventing their reaction with substrates. The removal of lignin leaves the cellulose more accessible and more open to swelling on contact with cellulase (Mooney et al., 1998). The degree that lignin adsorption of enzymes is decreased depends on the severity of pretreatment and the resulting decrease in lignin content (Ooshima et al., 1991). Addition of surfactants during hydrolysis can modify the cellulose surface properties. An important effect of surfactant addition in a process for lignocellulose conversion is the possibility to lower the enzyme loading. One reason for this effect might be adsorption of surfactants to lignin which prevents unproductive binding of enzymes to lignin and results in higher productivity of the enzymes (Eriksson et al., 2002).
Hemicellulose content: Because hemicellulose and lignin are covalently linked, acid hydrolysis of hemicellulose can open biomass structure as well. Moderate hemicellulose removal (>50%) is required to significantly increase cellulose digestibility (Mosier et al., 2005).
Acetyl content: In hemicellulose, xylan backbones are extensively acetylated (CH3COO-) (Holtzapple, 1993). Several studies showed that the removal of acetyl groups from hemicellulose reduced the steric hindrance of enzymes and greatly enhanced cellulose and xylan digestibility (Kong et al., 1992). The acetate groups interfere with enzyme recognition thereby slowing the hydrolysis rate.
Accessible surface area: Particle size associated with accessible surface area has a significant impact on the saccharification of lignocellulose by cellulolytic enzymes and is thought to be a controlling factor for conversion rates and yield (Zeng et al., 2007). Since, enzyme adsorption is a prerequisite step in the hydrolytic process, it seems that specific surface area would have an effect on hydrolysis rates since a higher surface area-to-weight ratio should mean more available adsorption sites per mass of substrate (Mansfield et al., 1999). It was hypothesized that initial rate of hydrolysis is a function of cellulose’s accessible surface area. Small particle sizes of untreated cellulosic substrate are more readily hydrolyzed than large ones because of higher specific surface area (Laivins and Scallan, 1996). Pretreatment increases accessible and susceptible surface area leading to enhanced enzymatic cellulose hydrolysis.
Cellulose crystallinity: Cellulose crystallinity presents a unified front that resists enzymatic attack. The rate of hydrolysis of amorphous cellulose is 3-30 times faster than that of high crystalline cellulose (Lynd et al., 2002). Several studies have shown that crystallinity prevents the rapid access of enzymes (Zhu et al., 2008); however, there are conflicting views about this (Fan et al., 1980). The conflict likely results from not distinguishing between the crystallinity of cellulose and biomass. For example, a pretreatment that selectively removes lignin enhances enzymatic digestibility, but would actually increase biomass crystallinity because lignin is amorphous. To properly assess the role of crystallinity, it would be necessary to know how the cellulose crystallinity, not the biomass crystallinity, is affected by the pretreatment. This is difficult to determine because measuring the crystallinity of cellulose that is blended with other biomass components is difficult.
Degree of polymerization: The degree of polymerization (DP, number of glucosyl residues per cellulose chain) of cellulosic substrates varies greatly, depending on substrate origin and preparation. It is still unclear if the DP of cellulose is a contributing limiting factor that influences the efficiency of enzymatic hydrolysis, because different conclusions have been drawn. Some of the results show that the DP of wood-derived cellulose fragments decreased with increasing enzyme hydrolysis time (Ramos et al., 1993) while others showed that the molecular weight of residual material remains unchanged after hydrolysis which indicates that the DP is relatively unimportant (Sinitsyn et al., 1991).
Enzyme-related factors
End-product inhibition: End-product inhibition is a major enzymatic factor that limits cellulase hydrolysis (Xiao et al., 2004). β-glucosidase, hydrolyzes the glucose dimer cellobiose to glucose. With a shortage of β-glucosidase, cellobiose accumulates, thereby inhibiting the action of endoglucanases and cellobiohydrolases in hydrolyzing cellulose. Adding extra β-glucosidase which hydrolyzes cellobiose to glucose, thereby preventing inhibition of cellobiohydrolases by cellobiose (Breuil et al., 1992). Increasing cellulase loading, removing sugars during hydrolysis by filtration (Gan et al., 2005) or using Simultaneous Saccharification and Fermentation (SSF) (Vinzant et al., 1999) are strategies designed to resolve this problem.
Enzyme synergism: Synergism seems to be particularly important for crystalline cellulose hydrolysis. Amorphous cellulose can be hydrolyzed by both endoglucanases and cellobiohydrolases while crystalline cellulose is largely hydrolyzed by cellobiohydrolases. Thus, crystallinity probably influences hydrolysis when synergism is lacking due to an incomplete cellulase system or an insufficient enzyme loading (Mansfield et al., 1999).
Ethanol fermentation: The lignocellulose hydrolysis yields D-glucose from cellulose and a mixture of hexoses (D-glucose, D-mannose, D-galactose, L-rhamanose), pentoses (D-xylose, L-arabinose) and uronic acids from hemicelluloses. As a result, the ability of the fermenting microorganisms to use the whole range of sugars available from the hydrolysate is vital to increase the economic competitiveness of cellulosic ethanol. The general requirements on an organism to be used in ethanol production is that it should give a high ethanol yield, a high productivity and be able to withstand high ethanol concentrations in order to keep distillation costs low (Von Sivers and Zacchi, 1996). In addition to these general requirements, inhibitor tolerance, temperature tolerance and the ability to utilize multiple sugars are essential. Tolerance towards low pH-values will minimize the risk of contamination.
Baker’s yeast (Saccharomyces cerevisiae) has long been employed for the industrial production of ethanol from hexose sugars (Kuyper et al., 2005). S. cerevisiae has many positive attributes which makes it suitable for industrial ethanol production such as a high rate of ethanol production from glucose. However this yeast species also has the limitation of not being able to ferment pentoses such as xylose or arabinose. Even though certain types of yeast, such as Pachysolen tannophilus, Pichia stipitis, or Candida shehatae, are xylose-fermenting, they have poor ethanol yields and low ethanol tolerance compared with the common glucose-fermenting yeasts, such as S. cerevisiae.
Thermophilic anaerobes have several advantages that make them suitable for an industrial conversion of lignocellulosic biomass to ethanol. First, there are organisms, such as Clostridium thermocellum that already have the ability to convert lignocellulosic biomass directly into ethanol. Second, the fermenters do not need aeration, what is an expensive part of industrial fermentation, because the organisms are anaerobes. Third, the optimal growth temperature of e.g., C. thermocellum is about 60°C; a high temperature simplifies the removal and recovery of ethanol by evaporation and distillation, renders expensive cooling systems unnecessary and furthermore it lessens problems with contaminations. Fourth, anaerobes have in general a low cell yield, so more of the substrate is converted to the endproduct (Demain et al., 2005). Despite the advantages of these organisms, one of the major drawbacks is that the achieved ethanol yields are still rather low compared to good ethanol producers like S. cerevisiae. This is to some part because during the fermentation of glucose not only ethanol is produced but also other compounds such as lactate and acetate. Another problem is the relative low tolerance of these organisms to ethanol. The tolerance to inhibitors that can be produced during the pretreatment can also be a problem. However, C. thermocellum lacks the ability to convert pentose sugars. Therefore the use of a coculture (or mixed culture) of C. thermocellum with another thermophilic anaerobe that is capable of producing ethanol out of pentoses is of great interest.
Zymomonas mobilis, an obligately anaerobic bacterium which gives a lower biomass yield and a higher ethanol yield on glucose compared to S. cerevisiae (Rogers et al., 1979). However, wild-type Z. mobilis lacks the ability to ferment pentose sugars and a major drawback is furthermore that it is not a very robust organism. In general, bacteria appear to be less tolerant to lignocellulose-derived inhibitors (Hahn-Hagerdal et al., 2007) and a detoxification step may be needed prior to the fermentation.
The inability of S. cerevisiae and Z. mobilis to utilize pentose sugars necessitates the search for pentose utilizing strains. Two groups of micro-organisms, i.e., enteric bacteria (E. coli and Klebsiella oxytoca) and some yeasts, are able to ferment pentoses, but with low ethanol yields. Furthermore, xylose fermenting yeasts (Pachysolen tannophilus, Candida shehatae and Pichia stipitis) are sensitive to high concentrations of ethanol (≥40 g L-1), require micro-aerophilic conditions, are highly sensitive to inhibitors and are not capable of fermenting xylose at low pH (Zaldivar et al., 2001).
As no naturally occurring organism can satisfy all necessary specifications (e.g., high ethanol yield, temperature tolerance, wide-substrate range, ethanol tolerance, tolerance to inhibitors and tolerance towards low pH-values), this has to come by the utilization of genetic engineering techniques for the construction of organisms that have most of the desirable characteristics for bioethanol production. Metabolic engineering for microorganisms used in fuel ethanol production has generated recombinant organisms displaying attractive features for the bioconversion of lignocellulose to ethanol (Jeffries and Jin, 2004). The three most promising microbial species that have been developed by metabolic engineering in the last two decades are S. cerevisiae, Z. mobilis and Escherichia coli. By genetic engineering it is possible to transfer cellulase genes from Trichoderma to Saccharomyces cerevisiae to produce “superstrains” via genetic enginnering capable of hydrolyzing cellulose and xylan along with fermentation of glucose and xylose into ethanol.
Due to the very attractive properties of S. cerevisiae in industrial fermentations, there have been significant efforts made to design recombinant S. cerevisiae strains to co-ferment the pentose sugars xylose and arabinose (Karhumaa et al., 2006). Furthermore, strains of S. cerevisiae were engineered to co-ferment mixtures of xylose and cellobiose, using a xylose fermenting strain and an intracellular β-glucosidase to hydrolyse cellobiose (Ha et al., 2011). It was shown that intracellular hydrolysis of cellobiose minimised glucose repression of xylose fermentation allowing co-consumption of cellobiose and xylose that improved ethanol yields.
According to the reactions, the theoretical maximum yield is 0.51 and 0.49 kg of ethanol kg-1 of C6 or C5 sugar (Qin, 2010):
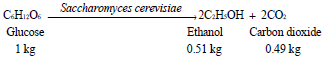

Hydrolysis and fermentation strategies: In the process of ethanol production out of lignocellulosic biomass 4 steps are driven by biological processes. (1) The production of the cellulase enzymes, (2) The hydrolysis of the cellulosic material, (3) The fermentation of hexose sugars and (4) The fermentation of pentose sugars.
Separate enzymatic hydrolysis and fermentation (SHF): In this process, pretreated lignocelluloses are hydrolyzed to glucose and subsequently fermented to ethanol in separate units (Fig. 4). The major advantage of this method is that it is possible to carry out the cellulose hydrolysis and fermentation at their own optimum conditions. The optimum temperature for cellulase is usually between 45 and 50°C, depending on the cellulose-producing microorganism (Saha et al., 2005; Olsson et al., 2006). However, the optimum temperature for most of the ethanol-producing microorganisms is between 30 and 37°C. Inhibition of cellulase activity by the released sugars, mainly cellobiose and glucose, is the main drawback of SHF. At a cellobiose concentration as low as 6 g L-1, the activity of cellulase is reduced by 60%. Although glucose decreases the cellulase activity as well, the inhibitory effect of this sugar is lower than that of cellobiose. On the other hand, glucose is a strong inhibitor for β-glucosidase. At a level of 3 g L-1 of glucose, the activity of β-glucosidase is reduced by 75% (Philippidis and Smith, 1995).
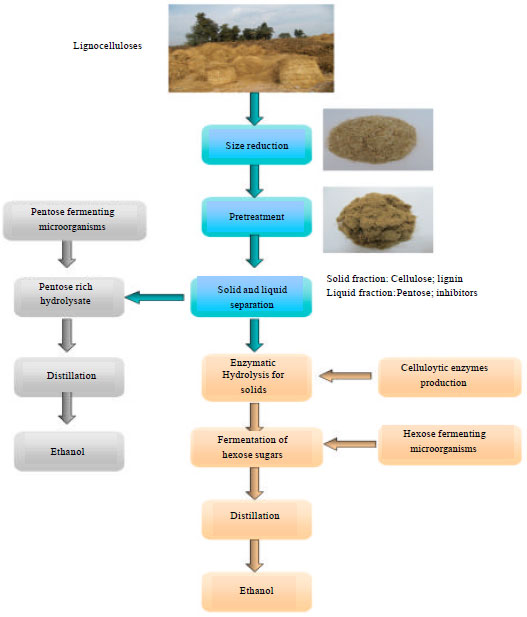 | |
| Fig. 4: | Schematic demonstration of bioethanol production using separate enzymatic hydrolysis and fermentation (SHF) |
Another possible problem in SHF is that of contaminations. The hydrolysis process is rather long, e.g., 1-4 days and a dilute solution of sugar always has a risk of microbial contaminations, even at rather high temperature such as 45-50°C. A possible source of contamination could be the enzymes. In practice, it is difficult to sterilize the cellulase in large scale, since it should be filtered because of its deactivation in an autoclave (Taherzadeh and Karimi, 2007b).
Simultaneous saccharification and fermentation (SSF): The Simultaneous Saccharification and Fermentation (SSF) combines the hydrolysis of pretreated lignocelluloses and the hexose fermentation in one step (Fig. 5). This configuration has the advantage that the sugars do not inhibit the cellulases during the hydrolysis because they are immediately used up by the fermenting organisms. In SSF process, the lignocellulosic biomass is first pretreated with a dilute acid (1.1% sulfuric acid at 160°C for 10 min) to breakdown lignin-hemicellulose complex. The resulting broth is filtered to drain the liquid from the system. The drained liquid containing pentose sugars is neutralized with lime and processed via., xylose fermentation process. Remaining solids containing cellulose and lignin is then hydrolyzed and fermented simultaneously by using cellulase enzymes and yeast.
The glucose produced by the hydrolyzing enzymes is consumed immediately by the fermenting microorganism present in the culture (Krishna et al., 2001).
 | |
| Fig. 5: | Schematic demonstration of bioethanol production using simultaneous saccharification and fermentation (SSF) |
This is a great advantage for SSF compared to SHF, since the inhibition effects of cellobiose and glucose to the enzymes are minimized by keeping a low concentration of these sugars in the media. SSF gives higher reported ethanol yields from cellulose than SHF and requires lower amounts of enzyme (Sun and Cheng, 2002; Demain et al., 2005; Karimi et al., 2006). The risk of contamination in SSF is lower than in the SHF process, since the presence of ethanol reduces the possibility of contamination. Furthermore, the number of vessels required for SSF is reduced in comparison to SHF, resulting in lower capital cost of the process (Table 4).
An important strategy in SSF is to have the optimum conditions for the enzymatic hydrolysis and fermentation as close as possible, particularly with respect to pH and temperature. However, the difference between optimum temperatures of the hydrolyzing enzymes and fermenting microorganisms is still a drawback of SSF. The optimum temperature for cellulases is usually between 45 and 50°C, whereas S. cerevisiae has an optimum temperature between 30 and 35°C and is practically inactive at more than 40°C. The optimum temperature for SSF by using T. reesei cellulase and S. cerevisiae was reported to be around 38°C which is a compromise between the optimal temperatures for hydrolysis and fermentation (Tengborg, 2000). Several thermotolerant bacteria and yeasts, e.g., Candida acidothermophilum and Kluyveromyces marxianus have been proposed for use in SSF to raise the temperature close to the optimal temperature of hydrolysis (Golias et al., 2002; Ballesteros et al., 2004; Hong et al., 2007).
| Table 4: | Comparison of the different fermentation process configurations |
 | |
 | |
| Fig. 6: | Schematic demonstration of bioethanol production using simultaneous saccharification and cofermentation (SSCF) |
The currently most promising ethanologenic bacteria for industrial exploitation are E. coli, Klebsiella oxytoca and Z. mobilis. Genetic engineering made it possible to transfer cellulose genes from Trichoderma to S. cerevisiae (Shoemaker, 1984). However, the cellulases were produced at a concentration too low to be useful.
Inhibition of cellulase by produced ethanol might be also a problem in SSF. It was reported that 30 g L-1 ethanol reduces the enzyme activity by 25% (Wyman, 1996). Ethanol inhibition may be a limiting factor in producing high ethanol concentration. Despite the mentioned problems, SSF is the preferred method in many laboratory studies and pilot scale studies for ethanol production. In the case of ethanol production from hardwood and agriculture residues, the hemicellulose mainly contains pentoses. If the pentose is separated during the pretreatment, the pentose-rich hydrolyzate (hemicellulosic hydrolyzate) can be converted to ethanol in a separate pentose-fermenting bioreactor (Fig. 5).
Simultaneous saccharification and cofermentation (SSCF): In Simultaneous Saccharification and Co-Fermentation (SSCF), the pretreated lignocellulosic biomass is neutralized and directly exposed to different enzymes and microorganisms that are capable of hydrolyzing cellulose and hemicelluloses to fermentable sugars as well as ferment hexoses and pentoses in one step to ethanol (Teixeira et al., 2000) (Fig. 6). One of the main problems of this approach is that the pentose utilizing organisms also prefer hexoses as substrate. So, if they are together with an organism as e.g., S. cerevisiae there is a competition between them, what usually results in lower ethanol yields. To overcome this effect sequential fermentation of hexoses and pentoses has been proposed, where the pentose fermenting organisms are added to the substrate after the hexose fermentation is completed, but in the same device. However ethanol yields of sequential fermentation are still low.
The process is carried out using genetically engineered microorganisms. Lawford and Rousseau (1998) used a metabolically engineered strain of Zymomonas mobilis that can coferment glucose and xylose. McMillan et al. (1999) used an adapted variant of the NREL xylose-fermenting Z. mobilis for ethanol production from dilute-acid- pretreated yellow poplar by SSCF.
 | |
| Fig. 7: | Schematic demonstration of bioethanol production using consolidated bioprocessing (CBP) |
The integrated system produced more than 30 g L-1 ethanol and achieved 54% conversion of all potentially available sugars in the biomass (total sugars) entering SSCF. This technology is superior to SSF technology in terms of cost effectiveness, better yields and shorter processing time (Table 4) (Lynd et al., 2005).
Consolidated bioprocessing (CBP): Consolidated bioprocessing (CBP) is a system in which cellulase production, substrate hydrolysis and fermentation are accomplished in a single step by cellulolytic microorganisms in microbial community and in one reactor (Fig. 7) (Lynd et al., 2005; Cardona and Sanchez, 2007). It is important to note that, in CBP, only one microbial consortium is employed for both the production of cellulase and fermentation. CBP offers the potential of lower production costs due to simpler lignocellulosic material processing, lower energy inputs and higher conversion efficiencies than SSF or SSCF based processes (Table 4). CBP is an economically attractive near-term goal for processes involving “third generation” biofuel production (Demain et al., 2005; Hamelinck et al., 2005). There are two strategies for enabling consolidated bioprocessing; (1) Modification of excellent ethanol producers, so that they also become efficient cellulase producers and (2) Modification of excellent cellulase producers, so that they also become efficient ethanol producers (Lynd et al., 2005).
There are as yet no organisms or compatible combinations of microorganisms available that produce cellulase and other enzymes at the required high levels and also produce ethanol at the required high yields and concentrations, although various organisms already combine multiple functions (Hamelinck et al., 2005). Den Haan et al. (2007) developed a recombinant strain of S. cerevisiae which can be used for CBP. Two cellulose-encoding genes, an endoglucanase of T. reesei and the β-glucosidase of Saccharomycopsis fibuligera, in combination, were expressed in S. cerevisiae. The resulting strain was able to grow on cellulose by simultaneous production of sufficient extracellular endoglucanase and β-glucosidase. They demonstrated the construction of a yeast strain capable of growing and of converting cellulose to ethanol in one step, representing significant progress towards realization of one-step processing of lignocellulose in a CBP configuration.
REFERENCES
- Alriksson, B., A. Cavka and L.J. Jonsson, 2011. Improving the fermentability of enzymatic hydrolysates of lignocellulose through chemical in-situ detoxification with reducing agents. Bioresour. Technol., 102: 1254-1263.
CrossRefDirect Link - Ballesteros, M., J.M. Oliva, M.J. Negro, P. Manzanares and I. Ballesteros, 2004. Ethanol from lignocellulosic materials by a simultaneous saccharification and fermentation process (SFS) with Kluyveromyces marxianus CECT 10875. Process Biochem., 39: 1843-1848.
CrossRefDirect Link - Berlin, A., M. Balakshin, N. Gilkes, J. Kadla, V. Maximenko, S. Kubo and J. Saddler, 2006. Inhibition of cellulase, xylanase and β-glucosidase activities by softwood lignin preparations. J. Biotechnol., 125: 198-209.
CrossRefDirect Link - Breuil, C., M. Chan, M. Gilbert and J.N. Saddler, 1992. Influence of β-glucosidase on the filter paper activity and hydrolysis of lignocellulosic substrates. Bioresour. Technol., 39: 139-142.
CrossRefDirect Link - Cardona, C.A. and O.J. Sanchez, 2007. Fuel ethanol production: Process design trends and integration opportunities. Bioresour. Technol., 98: 2415-2457.
CrossRefDirect Link - Chandel, A.K., M.L. Narasu, R. Rudravaram, R. Pogaku and L.V. Rao, 2009. Bioconversion of De-Oiled Rice Bran (DORB) hemicellulosic hydrolysate into ethanol by Pichia stipitis NCM3499 under optimized conditions. Int. J. Food Eng., Vol. 5.
CrossRef - Chandra, R.P., R. Bura, W.E. Mabee, A. Berlin, X. Pan and J.N. Saddler, 2007. Substrate pretreatment the key to effective enzymatic hydrolysis of lignocellulosics. Adv. Biochem. Eng. Biotechnol., 108: 67-93.
CrossRef - Chang, V.S. and M.T. Holtzapple, 2000. Fundamental factors affecting biomass enzymatic reactivity. Applied Biochem. Biotechnol., 84: 5-37.
CrossRefDirect Link - Chang, V.S., B. Burr and M.T. Holtzapple, 1997. Lime Pretreatment of Switchgrass. Proceedings of the 18th Symposium on Biotechnology for Fuels and Chemicals, May 5-9, 1996, Gatlinburg, Tennessee, pp: 3-19.
CrossRef - Chen, M., L. Xia and P. Xue, 2007. Enzymatic hydrolysis of corncob and ethanol production from cellulosic hydrolysate. Int. Biodeterior. Biodegrad., 59: 85-89.
CrossRef - Chundawat, S.P.S., B. Venkatesh and B.E. Dale, 2007. Effect of particle size based separation of milled corn stover on AFEX pretreatment and enzymatic digestibility. Biotechnol. Bioeng., 96: 219-231.
CrossRefDirect Link - Demain, A.L., M. Newcomb and J.D. Wu, 2005. Cellulase, clostridia and ethanol. Microbiol. Mol. Biol. Rev., 69: 124-154.
CrossRefDirect Link - Den Haan, R., S.H. Rose, L.R. Lynd and W.H. van Zyl, 2007. Hydrolysis and fermentation of amorphous cellulose by recombinant Saccharomyces cerevisiae. Metab. Eng., 9: 87-94.
CrossRefDirect Link - Draude, K.M., C.B. Kurniawan and S.J.B. Duff, 2001. Effect of oxygen delignification on the rate and extent of enzymatic hydrolysis of lignocellulosic material. Bioresour. Technol., 79: 113-120.
CrossRefDirect Link - El-Naggar, N.E.A., A.A. Sherief and S.S. Hamza, 2011. Bioconversion process of rice straw by thermotolerant cellulolytic Streptomyces viridiochromogenes for production of bioethanol under SSF conditions for bioethanol production. Afr. J. Biotechnol., 10: 11998-12011.
Direct Link - El-Naggar, N.E. and N.A.M. Abdelwahed, 2012. Optimization of process parameters for the production of alkali-tolerant carboxymethyl cellulase by newly isolated Streptomyces sp. strain NEAE-D. Afr. J. Biotechnol., 11: 1185-1196.
Direct Link - Eriksson, K.E.L., N. Habu and M. Samejima, 1993. Recent advances in fungal cellobiose oxidoreductases. Enzyme Microbial Technol., 15: 1002-1008.
CrossRefDirect Link - Eriksson, T., J. Borjesson and F. Tjerneld, 2002. Mechanism of surfactant effect in enzymatic hydrolysis of lignocellulose. Enzyme Microbial Technol., 31: 353-364.
CrossRef - Fan, L.T., Y.H. Lee and M.M. Gharpuray, 1982. The nature of lignocellulosics and their pretreatments for enzymatic hydrolysis. Adv. Biochem. Eng., 23: 157-187.
CrossRef - Fan, L.T., Y.H. Lee and D.H. Beardmore, 1980. Mechanism of the enzymatic hydrolysis of cellulose: Effects of major structural features of cellulose on enzymatic hydrolysis. Biotechnol. Bioeng., 22: 177-199.
CrossRef - Fan, L.T., Y.H. Lee and D.R. Beardmore, 1981. The influence of major structural features of cellulose on rate of enzymatic hydrolysis. Biotechnol. Bioeng., 23: 419-424.
CrossRef - Fonseca, B.G., R. de Oliveira Moutta, F. de Oliveira Ferraz, E.R. Vieira and A.S. Nogueira et al., 2011. Biological detoxification of different hemicellulosic hydrolysates using Issatchenkia occidentalis CCTCC M 206097 yeast. J. Ind. Microbiol. Biotechnol., 38: 199-207.
CrossRef - Galbe, M. and G. Zacchi, 2002. A review of the production of ethanol from softwood. Applied Microbiol. Biotechnol., 59: 618-628.
PubMed - Gama, F.M., M. Vilanova and M. Mota, 1998. Exo-and Endo-glucanolytic activity of cellulases purified from Trichoderma reesei. Biotechnol. Tech., 12: 677-681.
CrossRefDirect Link - Gan, Q., S. Allen and G. Taylor, 2005. Analysis of process integration and intensification of enzymatic cellulose hydrolysis in a membrane bioreactor. J. Chem. Technol. Biotechnol., 80: 688-698.
CrossRefDirect Link - Gaspar, M., G. Kalman and K. Reczey, 2007. Corn fiber as a raw material for hemicellulose and ethanol production. Process Biochem., 42: 1135-1139.
CrossRefDirect Link - Gharpuray, M.M., Y.H. Lee and L.T. Fan, 1983. Structural modification of lignocellulosics by treatment to enhance enzymatic hydrolysis. Biotechnol. Bioeng., 25: 157-172.
Direct Link - Golias, H., G.J. Dumsday, G.A. Stanley and N.B. Pamment, 2002. Evaluation of a recombinant Klebsiella oxytoca strain for ethanol production from cellulose by simultaneous saccharification and fermentation: Comparison with native Cellobiose-utilising yeast strains and performance in co-culture with thermotolerant yeast and Zymomonas mobilis. J. Biotechnol., 96: 155-168.
CrossRefDirect Link - Grethlein, H.E., 1985. The effect of pore size distribution on the rate of enzymatic hydrolysis of cellulosic substrates. Nature Biotechnol., 3: 155-160.
CrossRefDirect Link - Grethlein, H.E. and A.O. Converse, 1991. Common aspects of acid prehydrolysis and steam explosion for pretreating wood. Bioresour. Technol., 36: 77-82.
CrossRefDirect Link - Ha, S.J., J.M. Galazka, S.R. Kim, J.H. Choi and X. Yang et al., 2011. Engineered Saccharomyces cerevisiae capable of simultaneous cellobiose and xylose fermentation. Proc. Natl. Acad. Sci., 108: 504-509.
Direct Link - Hahn-Hagerdal, B., K. Karhumaa, C. Fonseca, I. Spencer-Martins and M.F. Gorwa-Grauslund, 2007. Towards industrial Pentose-fermenting yeast strains. Applied Microbiol. Biotechnol., 74: 937-953.
CrossRefDirect Link - Hamelinck, C.N., G.V. Hooijdonk and A.P. Faaij, 2005. Ethanol from lignocellulosic biomass: Techno-economic performance in Short-, middle-and Long-term. Biomass Bioenergy, 28: 384-410.
CrossRefDirect Link - Holtzapple, M.T., J.H. Jun, G. Ashok, S.L. Patibandla and B.E. Dale, 1991. The ammonia freeze explosion (AFEX) process. Applied Biochem. Biotechnol., 28: 59-74.
CrossRefDirect Link - Hong, J., Y. Wang, H. Kumagai and H. Tamaki, 2007. Construction of thermotolerant yeast expressing thermostable cellulase genes. J. Biotechnol., 130: 114-123.
CrossRefDirect Link - Howard, R.L., E. Abotsi, E.L.J. van Rensburg and S. Howard, 2003. Lignocellulose biotechnology: Issues of bioconversion and enzyme production. Afr. J. Biotechnol., 2: 602-619.
Direct Link - Hu, G., J.A. Heitmann and O.J. Rojas, 2008. Feedstock pretreatment strategies for producing ethanol from wood, bark and forest residues. BioResources, 3: 270-294.
Direct Link - Itoh, H., M. Wada, Y. Honda, M. Kuwahara and T. Watanabe, 2003. Bioorganosolve pretreatments for simultaneous saccharification and fermentation of beech wood by ethanolysis and white rot fungi. J. Biotechnol., 103: 273-280.
CrossRefDirect Link - Jeffries, T.W. and Y.S. Jin, 2004. Metabolic engineering for improved fermentation of pentoses by yeasts. Applied Microbiol. Biotechnol., 63: 495-509.
CrossRefDirect Link - Jorgensen, H., T. Eriksson, J. Borjesson, F. Tjerneld and L. Olsson, 2003. Purification and characterization of five cellulases and one xylanase from Penicillium brasilianum IBT 20888. Enzyme Microbial Technol., 32: 851-861.
CrossRefDirect Link - Jurado, M., A. Prieto, A. Martinez-Alcala, A.T. Martinez and M.J. Martinez, 2009. Laccase detoxification of Steam-exploded wheat straw for second generation bioethanol. Bioresour. Technol., 100: 6378-6384.
CrossRefDirect Link - Karhumaa, K., B. Wiedemann, B. Hahn-Hagerdal, E. Boles and M.F. Gorwa-Grauslund, 2006. Co-utilization of L-arabinose and D-xylose by laboratory and industrial Saccharomyces cerevisiae strains. Microbial Cell Factories, Vol. 5.
CrossRef - Karimi, K., G. Emtiazi and M.J. Taherzadeh, 2006. Ethanol production from dilute-acid pretreated rice straw by simultaneous saccharification and fermentation with Mucor indicus, Rhizopus oryzae and S. cerevisiae. Enzyme Microbial. Technol., 40: 138-144.
CrossRef - Kaylen, M., D.L. van Dyne, Y.S. Choi and M. Blase, 2000. Economic feasibility of producing ethanol from lignocellulosic feedstocks. Bioresour. Technol., 72: 19-32.
CrossRefDirect Link - Kheshgi, H.S., R.C. Prince and G. Marland, 2000. The potential of biomass fuels in the context of global climate change: Focus on transportation fuels. Ann. Rev. Energy Environ., 25: 199-244.
CrossRefDirect Link - Kim, T.H., J.S. Kim, C. Sunwoo and Y.Y. Lee, 2003. Pretreatment of corn stover by aqueous ammonia. Bioresour. Technol., 90: 39-47.
CrossRefDirect Link - Kirk, T.K. and R.L. Farrell, 1987. Enzymatic combustion: The microbial degradation of lignin. Annu. Rev. Microbiol., 41: 465-505.
CrossRef - Kong, F., C.R. Engler and E.J. Soltes, 1992. Effects of cell-wall acetate, xylan backbone and lignin on enzymatic hydrolysis of aspen wood. Applied Biochem. Biotechnol., 34-35: 23-35.
CrossRefDirect Link - Krause, D.O., S.E. Denman, R.I. Mackie, M. Morrison, A.L. Rae, G.T. Attwood and C.S. McSweeney, 2003. Opportunities to improve fiber degradation in the rumen: Microbiology, ecology, and genomics. FEMS Microbiol. Rev., 27: 663-693.
CrossRefPubMedDirect Link - Krishna, S.H., T.J. Reddy and G.V. Chowdary, 2001. Simultaneous saccharification and fermentation of lignocellulosic wastes to ethanol using a thermotolerant yeast. Bioresour. Technol., 77: 193-196.
CrossRef - Kumakura, M. and I. Kaetsu, 1983. Effect of radiation pretreatment of bagasse on enzymatic and acid hydrolysis. Biomass, 3: 199-208.
CrossRefDirect Link - Kumar, P., D.M. Barrett, M.J. Delwiche and P. Stroeve, 2009. Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Ind. Eng. Chem. Res., 48: 3713-3729.
CrossRefDirect Link - Kurakake, M., N. Ide and T. Komaki, 2007. Biological pretreatment with two bacterial strains for enzymatic hydrolysis of office paper. Current Microbiol., 54: 424-428.
CrossRefDirect Link - Kuyper, M., M.M. Hartog, M.J. Toirkens, M.J. Almering, A.A. Winkler, J.P. Dijken and J.T. Pronk, 2005. Metabolic engineering of a xylose‐isomerase‐expressing Saccharomyces cerevisiae strain for rapid anaerobic xylose fermentation. FEMS Yeast Res., 5: 399-409.
CrossRef - Laivins, G.V. and A.M. Scallan, 1996. The influence of drying and beating on the swelling of fines. J. Pulp Paper Sci., 22: 178-184.
Direct Link - Lawford, H.G. and J.D. Rousseau, 1998. Improving fermentation performance of recombinant zymomonas in acetic acid-containing media. Applied Biochem. Biotechnol., 70: 161-172.
CrossRefDirect Link - Lynd, L.R., W.H.V. Zyl, J.E. McBride and M. Laser, 2005. Consolidated bioprocessing of cellulosic biomass: An update. Curr. Opin. Biotechnol., 16: 577-583.
CrossRefDirect Link - Lynd, L.R., P.J. Weimer, W.H. van Zyl and I.S. Pretorius, 2002. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev., 66: 506-577.
CrossRefPubMedDirect Link - Mais, U., A.R. Esteghlalian, J.N. Saddler and S.D. Mansfield, 2002. Enhancing the enzymatic hydrolysis of cellulosic materials using simultaneous ball milling. Applied Biochem. Biotechnol., 98: 815-832.
CrossRefDirect Link - Malherbe, S. and T.E. Cloete, 2003. Lignocellulose biodegradation: Fundamentals and applications. Rev. Environ. Sci. Biotechnol., 1: 105-114.
CrossRef - Mamar, S. and A. Hadjadj, 1990. Radiation pretreatments of cellulose materials for the enhancement of enzymatic hydrolysis. Int. J. Radiat. Appl. Instrum. C, 35: 451-455.
CrossRef - McMillan, J.D., M.M. Newman, D.W. Templeton and A. Mohagheghi, 1999. Simultaneous saccharification and cofermentation of dilute-acid pretreated yellow poplar hardwood to ethanol using xylose-fermenting Zymomonas mobilis. Proceedings of the 20th Symposium on Biotechnology for Fuels and Chemicals, May 3-7, 1998, Gatlinburg, Tennessee, pp: 649-665.
- Mooney, C.A., S.D. Mansfield, M.G. Touhy and J.N. Saddler, 1998. The effect of initial pore volume and lignin content on the enzymatic hydrolysis of softwoods. Bioresour. Technol., 64: 113-119.
CrossRef - Mosier, N., C. Wyman, B. Dale, R. Elander, Y.Y. Lee, M. Holtzapple and M. Ladisch, 2005. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol., 96: 673-686.
CrossRefDirect Link - Neely, W.C., 1984. Factors affecting the pretreatment of biomass with gaseous ozone. Biotechnol. Bioeng., 26: 59-65.
CrossRef - Ohgren, K., M. Galbe and G. Zacchi, 2005. Optimization of steam pretreatment of SO2-impregnated corn stover for fuel ethanol production. Applied Biochem. Biotechnol., 124: 1055-1067.
CrossRef - Olsson, L., H.R. Soerensen, B.P. Dam, H. Christensen, K.M. Krogh and A.S. Meyer, 2006. Separate and simultaneous enzymatic hydrolysis and fermentation of wheat hemicellulose with recombinant xylose utilizing Saccharomyces cerevisiae. Applied Biochem. Biotechnol., 129: 117-129.
CrossRef - Ooshima, H., M. Kurakake, J. Kato and Y. Harano, 1991. Enzymatic activity of cellulase adsorbed on cellulose and its change during hydrolysis. Applied Biochem. Biotechnol., 31: 253-266.
CrossRef - Parawira, W. and M. Tekere, 2011. Biotechnological strategies to overcome inhibitors in lignocellulose hydrolysates for ethanol production: Review. Crit. Rev. Biotechnol., 31: 20-31.
CrossRef - Philippidis, G.P. and T.K. Smith, 1995. Limiting factors in the simultaneous saccharification and fermentation process for conversion of cellulosic biomass to fuel ethanol. Applied Biochem. Biotechnol., 51: 117-124.
CrossRef - Prates, J.A., N. Tarbouriech, S.J. Charnock, C.M. Fontes, L. Ferreira and G.J. Davies, 2001. The structure of the feruloyl esterase module of xylanase 10B from Clostridium thermocellum provides insights into substrate recognition. Structure, 9: 1183-1190.
CrossRef - Puri, V.P., 1984. Effect of crystallinity and degree of polymerization of cellulose on enzymatic saccharification. Biotechnol. Bioeng., 26: 1219-1222.
CrossRefDirect Link - Rabinovich, M.L., M.S. Melnick and A.V. Bolobova, 2002. The structure and mechanism of action of cellulolytic enzymes. Biochemistry (Moscow), 67: 850-871.
CrossRef - Ramos, L.P., M.M. Nazhad and J.N. Saddler, 1993. Effect of enzymatic hydrolysis on the morphology and fine structure of pretreated cellulosic residues. Enzyme Microb. Technol., 15: 821-831.
CrossRef - Rogers, P.L., K.J. Lee and D.E. Tribe, 1979. Kinetics of alcohol production by Zymomonas mobilis at high sugar concentrations. Biotechnol. Lett., 1: 165-170.
CrossRef - Saha, B.C., 2003. Hemicellulose bioconversion. J. Ind. Microbiol. Biotechnol., 30: 279-291.
CrossRefDirect Link - Saha, B.C., L.B. Iten, M.A. Cotta and Y.V. Wu, 2005. Dilute acid pretreatment, enzymatic saccharification and fermentation of rice hulls to ethanol. Biotechnol. Prog., 21: 816-822.
CrossRefDirect Link - Sameera, V., C. Sameera and Y. Ravi Teja, 2011. Current strategies involved in biofuel production from plants and algae. J. Microbial. Biochem. Technol.
Direct Link - Scott, C.D. and C.E. Wyman, 2004. Biotechnology for fuels and Chemicals-past, present and future. Proceedings of the 25th Symposium on Biotechnology for Fuels and Chemicals, May 4-7, 2003, Breckenridge, Co., pp: 761-763.
CrossRef - Sewalt, V.J.H., W.G. Glasser and K.A. Beauchemin, 1997. Lignin impact on fiber degradation. 3. Reversal inhibition of enzymatic hydrolysis by chemical modification of lignin and by additives. J. Agric. Food Chem., 45: 1823-1828.
CrossRefDirect Link - Sherief, A.A., N.E.A. El-Naggar and S.S. Hamza, 2010. Bioprocessing of lignocellulosic biomass for production of bioethanol using thermotolerant Aspergillus fumigates under solid state fermentation conditions. Biotechnology, 9: 513-522.
CrossRefDirect Link - Sinitsyn, A.P., A.V. Gusakov and E.Y. Vlasenko, 1991. Effect of structural and Physico-chemical features of cellulosic substrates on the efficiency of enzymatic hydrolysis. Applied Biochem. Biotechnol., 30: 43-59.
CrossRefDirect Link - Sun, Y. and J. Cheng, 2002. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresour. Technol., 83: 1-11.
CrossRefDirect Link - Sun, Y. and J.J. Cheng, 2005. Dilute acid pretreatment of rye straw and bermudagrass for ethanol production. Bioresour. Technol., 96: 1599-1606.
CrossRefDirect Link - Taherzadeh, M.J. and K. Karimi, 2007. Acid-based hydrolysis processes for ethanol from lignocellulosic materials: A review. BioResources, 2: 472-499.
Direct Link - Taherzadeh, M.J. and K. Karimi, 2007. Enzymatic-based hydrolysis processes for ethanol from lignocellulosic materials: A review. Bioresources, 2: 707-738.
Direct Link - Teixeira, L.C., J.C. Linden and H.A. Schroeder, 2000. Simultaneous saccharification and cofermentation of peracetic Acid-pretreated biomass. Applied Biochem. Biotechnol., 84: 111-127.
CrossRefDirect Link - Tran, A.V. and R.P. Chambers, 1986. Ethanol fermentation of red oak acid prehydrolysate by the yeast Pichia stipitis CBS 5776. Enzyme Microbial Technol., 8: 439-444.
CrossRefDirect Link - Vidal, P.F. and J. Molinier, 1988. Ozonolysis of lignin-Improvement of in vitro digestibility of poplar sawdust. Biomass, 16: 1-17.
CrossRefDirect Link - Vinzant, T.B., L. Ponfick, N.J. Nagle, C.I. Ehrman, J.B. Reynolds and M.E. Himmel, 1999. SSF comparison of selected woods from Southern Sawmills. Applied Biochem. Biotechnol., 45-46: 611-626.
CrossRefDirect Link - Von Sivers, M. and G. Zacchi, 1996. Ethanol from lignocellulosics: A review of the economy. Bioresour. Technol., 56: 131-140.
CrossRefDirect Link - Wyman, C.E., 2001. Twenty years of trials, tribulations and research progress in bioethanol technology: Selected key events along the way. Applied Biochem. Biotechnol., 91-93: 5-21.
CrossRefPubMedDirect Link - Xiao, Z., X. Zhang, D.J. Gregg and J.N. Saddler, 2004. Effects of sugar inhibition on cellulases and β-glucosidase during enzymatic hydrolysis of softwood substrates. Applied Biochem. Biotechnol., 115: 1115-1126.
CrossRef - Yang, B. and C.E. Wyman, 2004. Effect of xylan and lignin removal by batch and flowthrough pretreatment on the enzymatic digestibility of corn stover cellulose. Biotechnol. Bioeng., 86: 88-98.
CrossRefDirect Link - Yu, Y., Y. Feng, C. Xu, J. Liu and D. Li, 2011. Onsite Bio-detoxification of Steam-exploded corn stover for cellulosic ethanol production. Bioresour. Technol., 102: 5123-5128.
CrossRefDirect Link - Zaldivar, J., J. Nielsen and L. Olsson, 2001. Fuel ethanol production from lignocellulose: a challenge for metabolic engineering and process integration. Applied Microbiol. Biotechnol., 56: 17-34.
CrossRefDirect Link - Zeng, M., N.S. Mosier, C.P. Huang, D.M. Sherman and M.R. Ladisch, 2007. Microscopic examination of changes of plant cell structure in corn stover due to hot water pretreatment and enzymatic hydrolysis. Biotechnol. Bioeng., 97: 265-278.
CrossRef - Zhang, Y.H.P., M.E. Himmel and J.R. Mielenz, 2006. Outlook for cellulase improvement: Screening and selection strategies. Biotechnol. Adv., 24: 452-481.
CrossRefPubMedDirect Link - Zhu, J.Y., X.J. Pan, G.S. Wang and R. Gleisner, 2009. Sulfite pretreatment (SPORL) for robust enzymatic saccharification of spruce and red pine. Bioresour. Technol., 100: 2411-2418.
CrossRefDirect Link - Zhu, L., J.P. O'Dwyer, V.S. Chang, C.B. Granda and M.T. Holtzapple, 2008. Structural features affecting biomass enzymatic digestibility. Bioresour. Technol., 99: 3817-3828.
CrossRefDirect Link