
Fakher Rahim
Physiology Research Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Biotechnology
Year: 2010 | Volume: 9 | Issue: 2 | Page No.: 157-163







ABSTRACT
This study aimed to design a reliable homology model of human histamine H1 and H4 receptors (hH1R, hH4R), that would guide future biochemical and genetic efforts in its evaluation as a potential therapeutic target. Furthermore, these accurate models could aid in the structure-based inhibitor design for antagonists against the histamine H1 and H4 receptors. The homologous protein sequences of histamine receptors were retrieved from the NCBI REFSEQ which by using the sequence alignment program ClustalW alignment of the human histamine receptors sequence with Bovine Rhodopsin was conducted to locate the homology aligned regions. The present study found that Asp107 and Asn198 are in favorable positions for anchoring histamine. Identification of novel interaction sites for antagonist binding mutational data suggest a crucial role for Asp107, Trp158, Phe 432 and Phe 435 in antagonist binding. This study identified several novel amino acids at the binding site. Binding mode analysis of known H1 antagonists four known H1 antagonists (mepyramine, acrivastine, desloratadine, loratadine) were docked successfully to the binding site of the hH1R model by FlexiDock. The ligand used for optimizing the receptor model, the pharmacophore constraints and the different scoring functions applied in high throughput docking had all significant effect on the results. This research identified 16 compounds with 7 significant H4 activities representing an overall hit rate of 5.2%. To the best of our knowledge, this is one of the largest structure-based virtual screenings, where the virtual hits were confirmed by an in vitro assay. Moreover, this is the first structure-based drug design study reported on the hH4R. After the virtual screening, we identified several novel ligands with significant H4 affinity. These scaffolds can serve as starting points in the development of potent and selective H4 ligands in future.
PDF Abstract XML References Citation
How to cite this article
Fakher Rahim, 2010. An in Silico Development of Selective Inhibitor for Histamine Receptors. Biotechnology, 9: 157-163.
DOI: 10.3923/biotech.2010.157.163
URL: https://scialert.net/abstract/?doi=biotech.2010.157.163
DOI: 10.3923/biotech.2010.157.163
URL: https://scialert.net/abstract/?doi=biotech.2010.157.163
INTRODUCTION
In silico tools offer an attractive alternative strategy to the cumbersome experimental approaches (Gowthaman and Agrewala, 2009; Rahim, 2008). These computational tools have metamorphosed over the years into complex algorithms that attempt to efficiently predict the binding of peptides to receptors (Talele et al., 2009). Histamine receptors belong to class A of the G-Protein Coupled Receptor family. They are currently sub-classified into the four subtypes H1, H2, H3 and H4. These subtypes can be distinguished on the basis of their sensitivity to specific agonists and antagonists (Brown et al., 2001; Oda et al., 2000; Fox et al., 2005; Leurs et al., 2000) and by their molecular weight, which ranges from 45-60 kDa. Histamine has one of the broadest spectra among signaling molecules in the human body, ranging from involvement in allergies to contributing to the regulation of circadian rhythm in the brain (McEwen et al., 1997; Drzezga et al., 2001). Most of its well-defined actions are mediated by the H1, H2, H3 (Hill et al., 1997) and H4 receptor that has only recently been identified (Leurs et al., 2009; Zampeli and Tiligada, 2009).
Literature data suggest that histamine H1 and H4 receptors are potential therapeutic targets against allergy (Kiss and Keseru, 2009). H1 and H4 antagonists may be used separately or in combination representing an effective therapeutic option for allergy and other immunological diseases (Kiss and Keseru, 2009). The H1 receptor is most prominent in smooth muscle effects, especially those caused by IgE-mediated responses (Mitsuchashi and Payan, 1989). Bronchoconstriction and vasodilation, are typical retracts, opening gaps in the permeability barrier and resulting in the formation of local edema. These effects are manifest in allergic reactions and in mastocytosis, a rare neoplasm of mast cells.
H2 receptor mediates gastric acid secretion by parietal cells in the stomach. It also has a cardiac stimulant effect. A third action is to induce negative feedback upon histamine release from mast cells. The H3 receptor appears to be involved mainly in presynaptic modulation of histaminergic neurotransmission in the central nervous system. In the periphery, it appears to be a presynaptic heteroreceptor, which modulates the release of neurotransmitters other than those that stimulate it with modulatory effects on the release of other transmitters. Develop selective histamine receptor agonists and antagonists received considerable attention because of their potential effect as pharmaceutical agents in various human pathologies, including stomach acid disorders, duodenal ulcers, asthma, allergies. Biotechnology using molecular biology, biophysics, bioinformatics provides alternative methods for showing the classic drug interactions ligand-receptor characteristics, which the design should allow selective drugs based on detailed structural principles.
Hence, we felt that a reliable homology model of human histamine H1 and H4 receptors (hH1R, hH4R) would guide future biochemical and genetic efforts in its evaluation as a potential therapeutic target. Furthermore, these accurate models could aid in the structure-based inhibitor design for antagonists against the histamine H1 and H4 receptors.
MATERIALS AND METHODS
Data set and sequence alignment: This study was conducted from the 2008 to 2009. Amino acid sequence of human histamine receptors were downloaded in Gen Bank Flat File (GBFF) format at Los Alamos National Laboratory website (Pruitt et al., 2009). The homologous protein sequences of histamine receptors were retrieved from the NCBI REFSEQ using protein BLAST (Altschul et al., 1997; Pruitt et al., 2007). Using the sequence alignment program ClustalW (Version 1.81) alignment of the human histamine receptors sequence with Bovine Rhodopsin was conducted to locate the homology aligned regions (Fig. 1).
Homology modeling and docking: Homology models of hH1R and hH4R were built by SWISS-MODEL (Schwede et al., 2003). As a template, we used the crystal structure of bovine rhodopsin determined in 2000 (PDB ID: 1HZX). Using Mutate monomers each of the hH1R helices were mutated to the aligned sequences shown in Fig. 2 to generate the three different alignment models. The remaining loop sections of the hH1R model were completed by searching the PDB database using the Build loops function for matching amino acid sequence to the hH1R loop sequences.
 | |
| Fig. 1: | Alignment of the human histamine receptors sequence with Bovine Rhodopsin was conducted to locate the homology aligned regions |
 | |
| Fig. 2: | Phylogenetic relationships between the members of the human histamine receptor family. PHYLIP was used to generate the unrooted tree. PHYLODENDRON, a freestanding program was used to create the tree diagram |
The highest scored loops were inserted into the model and manual selection of the best fitting loops was employed. The final models were checked for amino acid chirality, proline ring integrity and were finally screened against the original hH1R sequence for monomer sequence. The three models were then subjected to energy minimisation within Sybyl® using 100 steepest descent iterations followed by 500 conjugate gradient steps using the standard Sybyl® setup for each minimisation. The three models, produced by the homology modelling described, are shown in Fig. 3a-c. The amino acid structures displayed in ball and stick style are the key amino acids previously shown to be important in ligand-receptor binding. The models were validated by the available mutagenesis data from literature and by a set of structural validation programs (PROCHECK, WHATIF, PROSA, HARMONY).
It is clear from these plots that all three structures have amino acid backbones conformations that are typically found in many other protein structures. Alignment 3, possessed the most favourable conformational space according to the Ramachandran analysis with 81.8% of the residues lying in the most favoured region (14.4% in allowed regions; 2.8% in generously allowed regions; 1.1% in disallowed regions) (Ramachandran et al., 1963).
Ligand docking with flexible protein side chains was carried out by FlexiDock. FlexX was used for docking with rigid protein side chains. High throughput docking on the hH4R model was exclusively done using the ClusterGrid production grid system developed by the National Information Infrastructure Development Institute (NIIF). Binding poses were scored by the own score of FlexX as well as the scoring functions available in the CScore package (Sybyl). The H4 activity of the virtual hits were evaluated by radioligand binding assay on a SK-N-MC cell line stably transfected with hH4R.
Phylogenetic analysis: An unrooted phylogenetic tree for the human Smad family based on MH1 sequences was constructed using algorithms contained within the PHYLIP Phylogeny Inference Package, version 3.5c (Felsenstein, 2001). PROTDIST was used on these sequences to calculate a distance matrix according to the Dayhoff PAM probability model (Dayhoff et al., 1983). The calculated distances represent the expected fraction of amino acid substitutions between each sequence pair. This distance matrix was then used to estimate the phylogenies using the Neighborhood Joining (NJ) method (Saitou and Nei, 1987). Bootstrapping was carried out using SEQBOOT (1000 replicates for the PAM substitution model). CONSENSE was then used to generate the consensus tree by the majority-rule method. Figure 2 shows the final unrooted tree diagram was generated using PHYLODENDRON (Hou et al., 1994).
RESULTS
The human histamine H1 Receptor (hH1R) model: According to the available mutational data, Asp107 and Asn198 have crucial roles in histamine binding, while Lys191 is mainly responsible for receptor activation. This study found that Asp107 and Asn198 are in favorable positions for anchoring histamine. On the other hand, Lys191 (5.39) is not able to form an H-bond with the imidazole N (1) of histamine. This study speculates that after histamine binding Lys191 can approach TM3. Consequently, the EC part of TM5 moves to the interior of the receptor, while the IC part, that is the G-protein binding site of hH1R, moves in the opposite direction. This can result in the activation of the receptor (Fig. 1).
Identification of novel interaction sites for antagonist binding Mutational data suggest a crucial role for Asp107, Trp158, Phe432 and Phe435 in antagonist binding. According to our model, we established that these residues are in favorable position to form interactions with ligands. We identified several novel amino acids at the binding site (Tyr108, Phe184, Phe190, Phe199 and Tyr431). The role of these residues in ligand binding was not described in the literature so far. Binding mode analysis of known H1 antagonists four known H1 antagonists (acrivastine, mepyramine, loratadine, desloratadine) were docked successfully to the binding site of the hH1R model by FlexiDock. Acrivastine, a second generation, zwitterionic antihistamine formed two ionic interactions with the side chains of Asp107 and Lys191 (Fig. 3).
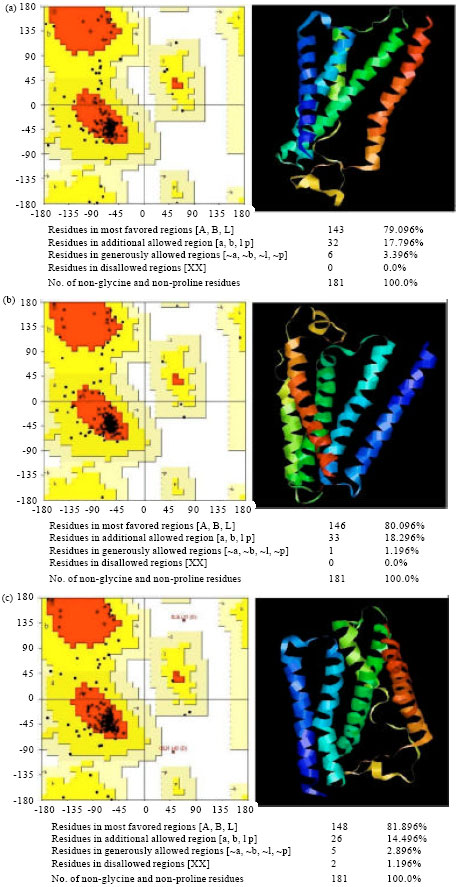 | |
| Fig. 3: | The three human histamine 1 receptors models (a-c) built by homology modelling to Bovine Rhodopsin. Ramachandran plot-darker colours represent favourable regions of amide bond torsions with black dots representing amino acids from the hH1 receptor model |
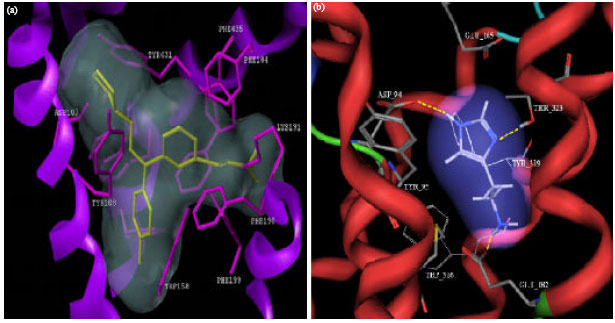 | |
| Fig. 4: | (a) Proposed binding mode of acrivastine at the hH1R binding site; (b) Proposed binding mode of histamine at the hH4R binding site |
The present study found that histamine and OUP-16 form complementary interactions with Asp94, Glu182 and Thr323, whereas JNJ7777120 interacts with Asp94 and Glu182 only.
Enrichment tests: We analyzed the applicability of six hH4R models developed by different methodologies for virtual screening by enrichment tests. We found that different inactive sets have only marginal effect on the highest achievable enrichment factors. On the other hand, the ligand used for optimizing the receptor model, the pharmacophore constraints and the different scoring functions applied in high throughput docking had all significant effect on the results.
Virtual screening: The database containing 7.8 million 3D-structures of small molecules was screened virtually on one of the developed hH4R models by FlexX. After the virtual screening we selected compounds for in vitro testing by two different methods using visual inspection and automatic filtering. In summary, 248 virtual hits were evaluated by radioligand binding assay. We identified sixteen compounds with 7 significant H4 activities representing an overall hit rate of 5.2% (Fig. 4a, b).
DISCUSSION
The present study has developed the structural model of hH1R by homology modeling as well as analyzed the role of amino acids at the binding site that were proved to take part in agonist or antagonist binding. Also, this study work identified several aromatic residues in suitable positions for antagonist binding. These novel interaction sites can be exploited in the design of new H1 antagonists. Bembenek et al. (2008) detail described, how the use of the available crystal structure information, pharmacophore modeling and docking lead to the identification of an inhibitor-histamine H(3) receptor antagonist. Spiegel et al. (2006) used decision analysis with budget impact modeling to measure the clinical and economic outcomes of these competing modes of administration. The present research builds the in silico homology model of hH4R that were refined with considering ligand information, docking and subsequent optimization.
Horr et al. (2006) investigated the role of the H4 receptor on STAT1/STAT6 responses in atopic and non-atopic lymphocytes by using the H4 receptor antagonist JNJ7777120 ex vivo.
The present study found that histamine and OUP-16 form complementary interactions with Asp94, Glu182 and Thr323, whereas JNJ7777120 interacts with Asp94 and Glu182 only, which shows the cost effective method to illustrate the interaction in silico.
Many previous and recent studies demonstrated that histamine binds to hH4R in a different conformation that was previously proposed in the literature (Schneider et al., 2009; Oda and Matsumoto, 2001; Strakhova et al., 2009). This study focused on the amino acids in the binding site and shows the binding property in the in silico manner.
This study found that ligand information can significantly influence the performance of the models in virtual screenings. On the other hand, the application of different inactive sets did not considerably affect the maximal achievable enrichment factors. According to the calculated enrichment factors, some of our hH4R models are suitable for virtual screening and therefore can be used to identify novel H4 ligands.
Finally, this study carried out a virtual screening on one of our hH4R models by docking all available 3D-structures of small molecules. To the best of our knowledge, this is one of the largest structure-based virtual screenings, where the virtual hits were confirmed by an in vitro assay. Moreover, this is the first structure-based drug design study reported on the hH4R. After the virtual screening, we identified several novel ligands with significant H4 affinity. These scaffolds can serve as starting points in the development of potent and selective H4 ligands in future.
CONCLUSION
Combination therapy will soon become a reality, particularly for those patients requiring poly-therapy to treat co-existing disease states. This becomes all the more important with the increasing cost, time and complexity of the drug discovery process prompting one to look at new delivery systems to increase the efficacy, safety and patient compliance of existing drugs. Along this line, we attempted to design in silico systems for simultaneous selective inhibitors for histamine receptors and model their and release kinetics. The attempts to model the in silico systems for simultaneous selective inhibitors for histamine receptors were successful, offering a possibility to use such models leading to high throughput screening of drugs and their nature, alone or in combination for receptors.
REFERENCES
- Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Bembenek, S.D., J.M. Keith, M.A. Letavic, R. Apodaca and A.J. Barbier et al., 2008. Lead identification of acetylcholinesterase inhibitors-histamine H3 receptor antagonists from molecular modeling. Bioorg. Med. Chem., 16: 2968-2973.
PubMed - Brown, R.E., D.R. Stevens and H.L. Hass, 2001. The physiology of brain histamine. Prog. Neurobiol., 63: 637-672.
Direct Link - Dayhoff, M.O., W.C. Barker and L.T. Hunt, 1983. Establishing homologies in protein sequences. Methods Enzymol., 91: 524-545.
PubMedDirect Link - Drzezga, A., U. Darsow, R.D. Treede, H. Siebner and M. Frisch et al., 2001. Central activation by histamine-induced itch: analogies to pain processing: A correlational analysis of O-15 H2O positron emission tomography studies. Pain, 92: 295-305.
PubMedDirect Link - Felsenstein, J., 2001. Taking variation of evolutionary rates between sites into account in inferring phylogenies. J. Mol. Evol., 53: 447-455.
PubMed - Fox, G.B., T.A. Esbenshade, J.B. Pan, R.J. Radek and K.M. Krueger et al., 2005. Pharmacological properties of ABT-239 [4-(2-{2-[(2R)-2-Methylpyrrolidinyl]ethyl}-benzofuran-5-yl)benzonitrile]: II. Neurophysiological characterization and broad preclinical efficacy in cognition and schizophrenia of a potent and selective histamine H3 receptor antagonist. J. Pharmacol. Exp. Ther., 313: 176-190.
PubMed - Gowthaman, U. and J.N. Agrewala, 2009. In silico methods for predicting T-cell epitopes: Dr Jekyll or Mr Hyde?. Expert Rev. Proteomics, 6: 527-537.
PubMed - Hill, S.J., C.R. Ganellin, H. Timmerman, J.C. Schwartz and N.P. Shankley et al., 1997. International union of pharmacology. XIII. classification of histamine receptors. Pharmacol. Rev., 49: 253-278.
PubMedDirect Link - Horr, B., H. Borck, J. Diel, C. Wackes, K. Nierich, R.L. Thurmond and F. Diel, 2006. The role of the histamine H4 receptor in signal transduction of atopic and non-atopic human lymphocytes ex vivo. Inflamm. Res., 55: S53-S54.
CrossRef - Hou, T.Z., J.Y. Luan, J.Y. Wang and M.D. Li, 1994. Experimental evidence of a plant meridian system: III. The sound characteristics of phylodendron (Alocasia) and effects of acupuncture on those properties. Am. J. Chin. Med., 22: 205-214.
PubMed - Kiss, R. and G.M. Keseru, 2009. Histamine H4 receptor ligands and their potential therapeutic applications. Expert Opin. Ther. Pat., 19: 119-135.
PubMed - Leurs, R., P.L. Chazot, F.C. Shenton, H.D. Lim and I.J. De-Esch, 2009. Molecular and biochemical pharmacology of the histamine H4 receptor. Br. J. Pharmacol., 157: 14-23.
PubMed - Leurs, R., M.S.R. Pena, R.A. Bakker, A.E. Alewijnse and H. Timmerman, 2000. Constitutive activity of G protein coupled receptors and drug action. Pharm. Acta Helv., 74: 327-331.
PubMed - McEwen, B.S., C.A. Biron, K.W. Brunson, K. Bulloch and W.H. Chamberset al., 1997. The role of adrenocorticoids as modulators of immune function in health and disease: Neural, endocrine and immune interactions. Brain Res. Rev., 23: 79-133.
PubMed - Mitsuchashi, M. and D.G. Payan, 1989. Molecular and cellular analysis of histamine H1 receptors on cultured smooth muscle cells. J. Cell Biochem., 40: 183-192.
PubMed - Oda, T., N. Morikawa, Y. Saito, Y. Masuho and S. Matsumoto, 2000. Molecular cloning and characterization of novel type of histamine receptor preferentially expressed in leukocytes. J. Biol. Chem., 275: 36781-36786.
PubMed - Oda, T. and S. Matsumoto, 2001. Identification and characterization of histamine H4 receptor. Nippon Yakurigaku Zasshi., 118: 36-42.
PubMed - Pruitt, K.D., T. Tatusova, W. Klimke and D.R. Maglott, 2009. NCBI reference sequences: current status, policy and new initiatives. Nucleic Acids Res., 37: D32-D36.
PubMed - Pruitt, K.D., T. Tatusova and D.R. Maglott, 2007. NCBI reference sequences (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res., 35: D61-D65.
Direct Link - Rahim, F., 2008. In silico comparison of simple sequence repeats in high nucleotides-rich genomes of microorganism. Pak. J. Biol. Sci., 11: 2372-2381.
CrossRefPubMedDirect Link - Ramachandran, G.N., C. Ramakrishnan and V. Sasiekharan, 1963. Stereochemistry of polypeptide chain configuarations. J. Mol. Biol., 7: 95-99.
PubMed - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Schneider, E.H., D. Schnell, D. Papa and R. Seifert, 2009. High constitutive activity and a G-protein-independent high-affinity state of the human histamine H(4)-receptor. Biochemistry, 48: 1424-1438.
PubMed - Schwede, T., J. Kopp, N. Guex and M.C. Peitsch, 2003. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res., 31: 3381-3385.
CrossRefPubMedDirect Link - Spiegel, B.M., G.S. Dulai, B.S. Lim, N. Mann, F. Kanwal and I.M. Gralnek, 2006. The cost-effectiveness and budget impact of intravenous versus oral proton pump inhibitors in peptic ulcer hemorrhage. Clin. Gastroenterol. Hepatol., 4: 988-997.
PubMed - Strakhova, M.I., C.A. Cuff, A.M. Manelli, T.L. Carr and D.G. Witte et al., 2009. In vitro and in vivo characterization of A-940894: A potent histamine H4 receptor antagonist with anti-inflammatory properties. Br. J. Pharmacol., 157: 44-54.
PubMed - Zampeli, E. and E. Tiligada, 2009. The role of histamine H4 receptor in immune and inflammatory disorders. Br. J. Pharmacol., 157: 24-33.
PubMed