
T. Taechowisan
Department of Microbiology, Faculty of Science, Silpakorn University, Nakorn Pathom 73000, Thailand
N. Chuaychot
Department of Microbiology, Faculty of Science, Silpakorn University, Nakorn Pathom 73000, Thailand
S. Chanaphat
Department of Microbiology, Faculty of Science, Silpakorn University, Nakorn Pathom 73000, Thailand
A. Wanbanjob
Department of Chemistry, Faculty of Science, Silpakorn University, Nakorn Pathom 73000, Thailand
P. Tantiwachwutikul
Department of Chemistry, Faculty of Science, Silpakorn University, Nakorn Pathom 73000, Thailand







ABSTRACT
In an attempt to understand the mode of action of this antagonist in nature, the interaction between Streptomyces sp. SRM1 and C. musae was studied by dual culture on agar plates. Evidence for the antibiosis of Streptomyces sp. SRM1 was demonstrated by inhibition zones in in vitro plate assay. The crude extract from the culture of Streptomyces sp. SRM1 also produced antifungal activity, which showed antagonistic effects against C. musae such as swelling, distortion and excessive branching of hyphae and inhibition of spore germination. An indirect method was used to show antagonistic effect of Streptomyces sp. SRM1 against C. musae in soil. This study suggests the potential of developing Streptomyces sp. SRM1 for the biological control of anthracnose disease of banana caused by C. musae.
PDF Abstract XML References Citation
How to cite this article
T. Taechowisan, N. Chuaychot, S. Chanaphat, A. Wanbanjob and P. Tantiwachwutikul, 2009. Antagonistic Effects of Streptomyces sp. SRM1 on Colletotrichum musae. Biotechnology, 8: 86-92.
DOI: 10.3923/biotech.2009.86.92
URL: https://scialert.net/abstract/?doi=biotech.2009.86.92
DOI: 10.3923/biotech.2009.86.92
URL: https://scialert.net/abstract/?doi=biotech.2009.86.92
INTRODUCTION
Wound anthracnose is the main disease affecting the quality of export banana fruits from many country including Thailand. This postharvest disease is caused by the pathogenic fungus Colletotrichum musae. In plantations, C. musae conidia contaminate banana fruits during the month after flowering (Chillet et al., 2000). These conidia quickly germinate and form melanised appressoria, which are quiescent structures of the pathogen (De Lapeyre de Bellaire et al., 2000). These appressoria germinate as the banana ripen and form infected hyphae that colonise the peel and penetrate into the fruit pulp (Chillet et al., 2007), leading to the development of anthracnose. Although a rather tedious procedure of injecting the fungicides thiabendazole or imazalil into plantation has been attempted, there are no effective chemical control measures for this disease. Strains of C. musae exist which are resistant to thiabendazole (Johanson and Blazquez, 1992). Some microorganisms, including Mycoparasites; Gliocladium sp., Trichoderma sp., Pythium sp., Verticillium sp. and Clonostachys rosea (Krauss et al., 1998), have been reported to be potential antagonists of C. musae. These microorganisms can be developed as potential biocontrol agents against C. musae by understanding the type of interactions between the fungal pathogen and the control agent (Krauss and Soberanis, 2001). Generally, mechanisms through which microorganisms inhibit fungal pathogens in the rhizosphere are: (i) competition for nutrients, oxygen or space; (ii) parasitism or the physical destruction of fungal cell walls by the action of hydrolytic enzymes produced by the antagonist; (iii) antibiosis or the inhibition of one microorganism by diffusible compound(s) produced by another and/or, by a synergistic combination of these modes of action (Benyagoub et al., 1998).
Actinomycetes of the genus Streptomyces are well known for their ability to suppress growth of a wide variety of fungal pathogens (Taechowisan et al., 2003a; Trejo-Estrada et al., 1998a). Streptomyces species have been used extensively in the biological control of several phytopathogenic fungi (El-Raheem et al., 1995). Many antibiotics produced by actinomycetes have been used directly or have been assumed to be responsible for the biocontrol potential of the producing strain (Smith et al., 1990; Trejo-Estrada et al., 1998b). Streptomyces are also known for their ability to produce fungal cell wall-degrading enzymes such as cellulases, hemicellulases, glucanases and chitinases (Taechowisan et al., 2003b; Trejo-Estrada et al., 1998b). The role of these hydrolytic enzymes in antifungal activity and biocontrol ability of Streptomyces has been investigated (Taechowisan et al., 2003b; Valois et al., 1996). The earlier studies, an endophytic Streptomyces aureofaciens CMUAc130 was shown to exhibit strong in vitro antagonism toward several plant pathogenic fungi at least two mechanisms, one was chitinolytic enzyme and other was secondary metabolites (Taechowisan et al., 2003b; Taechowisan et al., 2005). The isolate, referred to as strain SRM1, showed strong antagonism toward C. musae. The isolation, selection and identification of strain SRM1, as well as the antagonistic effects of this strain against C. musae on hyphal extension and pathogenesis and also spore germination were investigated.
MATERIALS AND METHODS
Fungal pathogen: Fungal strain used in the in vitro antagonism assay was: Colletotrichum musae, the causative agents of anthracnose of banana (provided by Dr. Wipornpan Photita, Department of Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai, Thailand). It was grown on Potato Dextrose Agar (PDA) plates at 30°C and stored on PDA slants at 4°C.
Isolation of actinomycetes: Actinomycetes were isolated from soil samples collected from the environs of Nakorn Pathom, Thailand, during the period April-July 2007. Samples were dried in room temperature for 5 days and then suspended in sterile saline (0.9% NaCl) solution. The serial dilution spread plate technique was used, where dilution 10-4 was plated onto humic acid-vitamin (HV) agar (Otoguro et al., 2001) containing 100 μg mL-1 nystatin and cycloeximide and incubated at 30°C for 1 week. The colonies were inoculated onto International Streptomyces Project-2 (ISP-2) medium (Shirling and Gottlieb, 1966) for purification and stock cultures.
Identification of endophytic actinomycetes: The characteristics of the actinomycetes isolates were observed. For morphological characteristics, presence of aerial mycelium, spore mass colour, distinctive reverse colony colour, diffusible pigment, sporophore and sporechain morphology were recorded after 10 days incubation on ISP-2 medium. Diaminopimelic acid isomers and sugars from whole-cell extract were analysed for chemotaxonomic studies (Becker et al., 1964; Boone and Pine, 1968).
In vitro assay for antagonism: An in vitro plate-assay technique was developed to test the inhibitory effect of actinomycete isolates on C. musae growth. Test for inhibitory activity was made on ISP-2 in Petri dishes. A 0.8 cm diameter ISP-2 agar plug, covered fully with a lawn of actinomycete isolate, was placed 1.5 cm from the edge of the Petri dish and incubated at 30°C for 5 days. This was done to allow the culture to be established on the agar surface and to sporulate prior to inoculation of the plates with fungi. In order to investigate the inhibitory effect of actinomycetes on the fungal growth, a 0.8 cm diameter PDA plug covered with activity growing mycelium was placed about 6 cm from actinomycete colony. The inoculated plates were placed in an incubator at 30°C for 4 days. The inhibition zone was determined by measuring the distance between the fungi and actinomycete colony in dual cultures.
Extraction of secondary metabolites produced by Streptomyces sp. SRM1: Among the 39 isolates of actinomycetes, the isolate SRM1 was found to be the best producer of antifungal substances. This isolate was selected for extraction the secondary metabolites. Spores of Streptomyces sp. SRM1 were used to inoculate 250 plates of ISP-2 and incubated for 14 days at 30°C. The culture medium was then cut into small pieces that were extracted with ethyl acetate (3x300 mL). This organic solvent was pooled and then taken to dryness under rotary evaporation to give a dark brown solid (250 mg).
In vitro antagonistic effects of Streptomyces sp. SRM1 metabolites on mycelial development and spore germination of C. musae: Crude extract from Streptomyces sp. SRM1 was tested for antibiosis using the disk diffusion assay as described by Milici et al. (2007). The crude extract was dissolved in methanol (25, 50 and 100 μg mL-1) and 50 μL was applied to sterile (6 mm diameter) paper disks (Advantec, Toyo Roshi Kaisha, LTD., Japan), dried and then placed on PDA plate, each plate was then incubated with an agar block (8 mm diameter) containing mycelial mats of the fungi in the center of the plate (the paper discs were 2.2 cm from the fungi). Plates were incubated at 30°C for 72 h and observed for the presence of an inhibition zone. After 72 h, morphological changes on fungal mycelial occurring in the margin of the inhibition zones were examined under a compound microscope at 400x magnification. The experiment was repeated twice.
Fungal spore germination assays were performed according to the procedure described by Lorito et al. (1993) with minor variations. A stock suspension of ca. 106 conidia of the fungus mL-1 in 30% glycerol was prepared and kept at -20°C. Equal volumes (400 μL) of spore suspension, 3x potato dextrose broth and the crude extract (384 μg mL-1 in 10% (v/v) methanol) were mixed in sterile microcentrifuge tube. The test solutions were replaced with sterile water in control samples. Tubes were incubated at 30°C for 24 h. A drop of the mixture from each tube was placed on a microscope slide and the percentage of conidial germination was determined from the first 100 spores chosen at random. Data were transformed into values representing the treatments as a percentage of the control (in which % inhibition of control = 0) by the following equation:
where, I(%) represented the percent inhibition, St(%) represented the percentage of spores germinating in the treatment of interest and Sc(%) represented the percentage of spores germinating in the control. The results of each experiment are reported as the average of three replicates.
Minimum Inhibitory Concentrations (MICs): MICs of crude extract was determined by microbroth dilution methods (Milici et al., 2007).
The fungal pathogen C. musae was tested for its response to the crude extract using a Potato Dextrose Agar (PDA) dilution technique. The crude extract (5.12 mg) was dissolved in DMSO (1 mL), then serially diluted two-fold to obtain final concentration ranges of 0.50-256 μg mL-1 in PDA. The medium (5 mL) was added to a 5 cm diameter Petri dish. An 8 mm diameter plug of the fungi, removed from the margin of a 4 day old colony on PDA, was placed 1.5 cm from the edge of the plate. Linear growth of the fungi at 30°C was recorded 2 days after treatment. Each treatment consisted of three replicates. The experiment was repeated twice.
Direct observation of the antagonistic effects of strain SRM1 on spore germination of C. musae in soil environment: A buried slide technique (Stevenson, 1956) was used. About 50 g of soil (pH = 7.6) was sieved, air-dried and sterilized in 250 mL plastic bags by autoclaving at 121°C for 30 min. A 5 mL spore suspension of strain SRM1 containing 3x108 colony forming units mL-1 was prepared from ISP-2 plates. Spores were washed thoroughly with sterile distilled water, inoculated into the sterile soil and incubated at 30°C for 7 days. A 10 mL spore suspension of C. musae containing 2x106 spores mL-1 was mixed in 100 mL of sterile 1.8% molten agar and coated on sterile microscope glass slides by dipping them. When the agar layer had set, slides were carefully inserted vertically into plastic bags containing sterile soil alone (control) or strain SRM1-inoculated soil and incubated at 30°C. At the end of each specified incubation period (2, 4 and 7 days), the slides were removed carefully and examined immediately under a compound microscope at 400X magnification. Each observation period involved analysis of two plastic bags for each treatment. Each plastic bag contained three spore-coated glass slides. After the second day of incubation, the number of spores that germinated and formed germ tubes on the control and test slides was compared visually under a light microscope. The percentage of spore germination was calculated in 30 microscopic fields.
RESULTS AND DISCUSSION
Thirty-nine strains were isolated and classified into Streptomycete -like strains by colony and morphological observation. All isolates were tested for the in vitro antagonistic assay against C. musae. Six isolates (J72, KJ5, NC1, NC2, SRM1 and WTA2) inhibited the growth of C. musae. The inhibition zones of this tested fungi was over 10 mm when cultured dually with these six isolates for 4 days at 30°C. One isolate, SRM1, showed strong antagonism towards C. musae.
Morphological observation of 10 day old culture of SRM1 revealed that sporophores to be monopodially branched and flexuous, producing open spirals of oval-shaped spores (1x1.5 μm) with smooth surfaces. The substrate mycelium was extensively branched with non-fragmenting hyphae. The aerial mycelium was white changing to ash-grey with yellow soluble pigment occasionally discernable. Based on results in morphological observation as well as on the presence of LL-diaminopimelic acid in the whole-cell extract, the isolate SRM1 was identified as belong to the genus Streptomyces (Fig. 1).
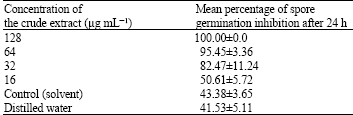
Since, the isolate SRM1 producing the most active substance, it was cultured in the large scale to get more culture and its secondary metabolites were extracted by ethyl acetate. The crude extract of SRM1 at a concentration of 1.25, 2.5 and 5 μg disc-1 produced inhibition zones with a diameter of about 12-28 mm on paper disc assay (Fig. 2a). When observed under a dissecting microscope, fungal mycelium along the edges of the colonies facing the crude extract of SRM1 appeared thickened, with bulbous-like formations along the ends (Fig. 2b, c). On the control disc, however, fungal mycelium showed regular, radial growth. The different concentration of the crude extract showed efficacy in suppressing C. musae. The minimum concentration of the crude extract for inhibition in hyphal extension of C. musae was 25 μg mL-1. The crude extract of SRM1, at a concentration 128 μg mL-1 completely inhibited spore germination of C. musae (Table 1). After 24 h of incubation, spore germination of C. musae was reduced when increasing the concentration of the crude extract, in contrast, a high degree of germination occurred in both experiments when spores were mixed with either distilled water or with 10% (v/v) methanol. The presence of solvent in the crude extract sample did not have any negative effect on spore germination when compared with distilled water (Fig. 3).
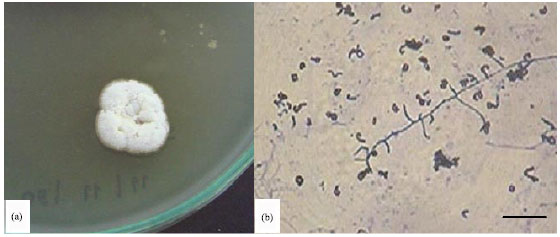 | |
| Fig. 1: | Morphology of Streptomyces sp. SRM1 colony (a) and mycelium (b). A white colony , changing to orange with faint yellowish soluble pigment. An open spirals, spores spherical to-oval-shaped under light microscopy observation. Bar = 10 μm |
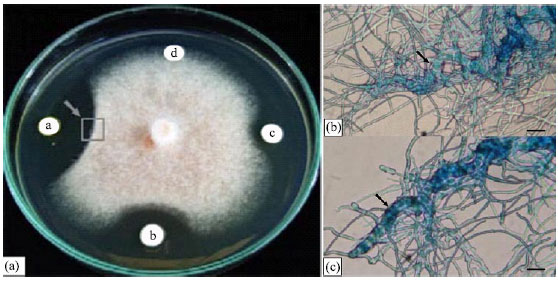 | |
| Fig. 2: | Antagonism of the crude extract of Streptomyces sp. SRM1 to Colletotrichum musae on ISP-2 medium. (a) In vitro plate assay after 72 h incubation at difference concentration, a = 5, b = 2.5, c = 1.25 μg disc-1 and d = control (arrow shows the source of fungal-mycelium samples for microscopic studies). (b and c) Hyphae showing thickened and bulbous structures (arrow) at the edges of the inhibited fungal colonies in the in vitro antagonistic assay. Bar = 10 μm |
| Table 1: | Effect of crude extract of Streptomyces sp. SRM1 on spore germination inhibition of Colletotrichum musae |
 | |
Results from the buried slide technique showed that the spores of C. musae on glass slides buried in uninoculated sterile soil germinated normally and the germ tubes developed into long hyphal strands. By the 7th day, healthy and extensive hyphal growth with sporulation was evident on the control slides (Fig. 4). However, in the sterile soil in which the isolate SRM1 was grown, fungal spore germination was reduced compared to the control slides. The percentage of spore germination was observed 27.62 and 18.24% in the percent of SRM1 on the second and fourth day while there are no observation of fungal spore germination in the present of SRM1 on the seventh day. A lytic effect caused by SRM1 was first detected in the fungal hyphae after 2 days of incubation. Hyphal distortion, like swelling or bulbous growth, was also observed. By the fourth day, distortion and lysis of the hyphae were more frequently noted.
 | |
| Fig. 3: | Spore germination inhibition after incubation at 30°C for 24 h with: (a) 20 mL sterile distilled water, (b) 10% (v/v) methanol, (c) 64 μg mL-1 of crude extract and (d) 128 μg mL-1 of crude extract. Bar = 10 μm |
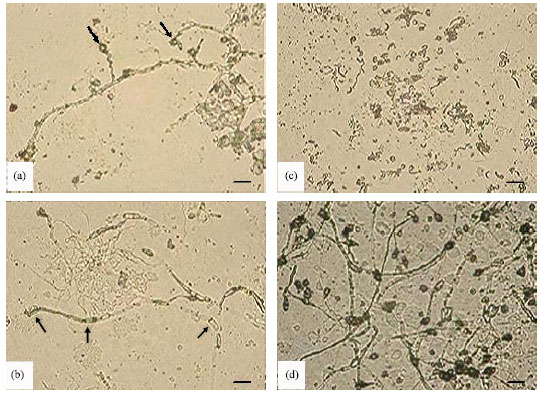 | |
| Fig. 4: | Hyphal development of Colletotrichum musae in SRM1-inoculated control soil (x400). (a) Early stages of hyphal distortion and lysis (arrowed) after 2 days in soil culture. (b) Distorted and swellings (arrow) of hyphae after 4 days in soil culture. (c) Massive lysis of hyphae after 7 days in soil culture, hyphae is less while the spore of SRM1 is abundant. (d) Healthy hyphal growth with abundant sporulation after 7 days in uninoculated control soil. Bar = 10 μm |
All of actinomycetes isolated in this study belong to the Streptomycete-like group. The predominance of Streptomycetes in actinomycete population in soil sample is distributed at a high frequency. This could also be due to the fact that no special pretreatment techniques were adapted to enhance the isolation of rare actinomycetes for example; HV agar was amended with antibacterial agents was used in conjunction with physical (dry heating) and chemical (phenol, chlorhexidine gluconate, benzethonium chloride and γ-collidine) pretreatments to achieve the highly selective isolation of the Micromonospora, Microbispora, Streptosporangium, Dactylosporangium and Actinoplanes (Hayakawa et al., 1991a-c).
In the in vitro antagonistic assay showed promising results for the use of streptomycetes as antifungal agents against phytopathogenic fungi. Similar results have been reported previously in actinomycetes screening studies (Crawford et al., 1993). It was noted that some actinomycetes which inhibited pathogens on agar plate also did so, in soil (Fravel, 1988). Thus, in vitro assays appear to be useful in identifying which actinomycete antagonists might function in soil.
The isolate SRM1 was selected for one trait of ten associated with biocontrol agents, its ability to produce the secondary metabolites active against C. musae. In the in vitro assay, preinoculation of the isolate SRM1 was done to allow growth and spore germination of the culture prior to inoculation with C. musae. Thus, the antagonism between the isolate SRM1 and the C. musae may have involved production of secondary metabolites in agar. The in vitro studies further indicated that culture of the isolation SRM1 in agar medium produced the secondary metabolites that showed clear zones of inhibition against C. musae and induced morphological changes such as swollen and distorted germ tubes that branched more freely than normal conidia. The secondary metabolites from the crude extract also inhibited spore germination and hyphal development of C. musae. The isolate SRM1 also showed the inhibitory effects on spore gemination and hyphal lysis of C. musae in soil by the buried slide technique. These results indicated that using the secondary metabolites of SRM1 could be controlling fungal growth and might be a promising method of biocontrol of some plant pathogens.
Antibiosis is particularly considered to provide an advantage in biological disease control because compounds mediating antibiosis can diffuse rapidly in nature and direct contact between the antagonist and pathogen is nor necessary (Hajlaou et al., 1994).
Antibiotics have been implicated in the antagonism of fungi by actinomycetes. These are many reports related to antibiotic substances that induced malformations such as stunting, distortion, swelling, hyphal protuberances or the highly branched appearance of fungal germ tubes (Gunji et al., 1983). Previously, a strain of Streptomyces hygroscopicus var. geldanus produced geldanamycin in culture and in soil was able to biocontrol diseases caused by Rhizoctonia solani (Rothrock and Gottlieb, 1984) and other studies, a strain of Streptomyces violaceusniger that produced at least three antifungal compounds has been reported to suppress damping off of lettuce caused by Pythium ultimum (Trejo-Estrada et al., 1998b). On the other hand, Streptomyces are also known for their ability to cause lysis of fungal hyphae by producing chitinases and glucanases (Taechowisan et al., 2003b; Valois et al., 1996).
This study demonstrated that antibiosis mediated by diffusible metabolites was involved in the antagonism of strain SRM1 against C. musae in sterile soil. However, further studies are needed to determine whether inhibition of fungal spore germination and hyphal swelling, distortion and lysis will be detected when C. musae is placed in strain SRM1-inoculated nonsterile soils. In addition further studies are needed to characterize the antifungal substances of strain SRM1, for example bioactive compound(s) and hydrolytic enzyme production. These studies will be useful in the manipulation and development of strain SRM1 as a potential biological control agent for anthracnose of banana caused by C. musae.
ACKNOWLEDGMENT
This study was supported by Thailand Research Fund MRG5180173 and the Faculty of Science, Silpakorn University, Nakorn Pathom, Thailand.
REFERENCES
- Becker, B., M.P. Lechevalier, R.E. Gordon and H.A. Lechevalier, 1964. Rapid differentiation between Nocardia and Streptomyces by paper chromatography of whole-cell hydrolysates. Applied Microbiol., 12: 421-423.
PubMed - Boone, C.J. and L. Pine, 1968. Rapid method for characterization of actinomycetes by cell wall composition. Applied Microbiol., 16: 279-284.
Direct Link - Chillet, M., L. de Lapeyre de Bellaire, M. Dorel, J. Joas, C. Dubois, J. Marchal and X. Perrier, 2000. Evidence for the variation in susceptibility of bananas to wound anthracnose due to Colletotrichum musae and the influence of edaphic conditions. Sci. Hortic., 86: 33-47.
CrossRef - Chillet, M., O. Hubert, and L. de Lapeyre de Bellaire, 2007. Relationship between physiological age, ripening and susceptibility of banana to wound anthracnose. Crop Protect., 26: 1078-1082.
CrossRef - Crawford, D.L., J.M. Lynch, J.M. Whipps and M.A. Ousley, 1993. Isolation and characterization of actinomycete antagonists of a fungal root pathogen. Applied Environ. Microbiol., 59: 3899-3905.
Direct Link - De Lapeyre de Bellaire, L., M. Chillet, C. Dubois and X. Mourichon, 2000. Importance of different source of inoculum and dispersal methods of conidia of Colletotrichum musae, the causal agent of banana anthracnose, for fruit contamination. Plant Pathol., 49: 782-790.
CrossRef - El-Raheem, A., R. El-Shanshoury, S.M. El-Sououd, O.A. Awadalla and N.B. El-Bandy, 1995. Formation of tomatine in tomato plants infected with Streptomyces species and treated with herbicides, correlated with reduction of Pseudomonas solanacearum and Fusarium oxysporum f. sp. lycopersici. Acta Microbiol. Pol., 44: 255-266.
PubMed - Fravel, D.R., 1988. Role of antibiosis in the biocontrol of plant diseases. Annu. Rev. Phytopathol., 26: 75-91.
CrossRef - Gunji, S., K. Arima and T. Beppu, 1983. Screening of antifungal antibiotics according to activities inducing morphological abnormalities. Agric. Biol. Chem., 47: 2061-2069.
CrossRefDirect Link - Hajlaou, M.R., J.A. Traquair, W.R. Jarvis and R.R. Belanger, 1994. Antifungal activity of extracellular metabolites produced by Sporothrix flocculosa. Biocontrol Sci. Technol., 4: 229-237.
CrossRef - Hayakawa, M., T. Kajiura and H. Nonomura, 1991. New methods for the highly selective isolation of Streptosporangium and Dactylosporangium from soil. J. Ferment. Bioeng., 72: 327-333.
CrossRefDirect Link - Hayakawa, M., T. Sadakata, T. Kajiura and H. Nonomura, 1991. New methods for the highly selective isolation of Micromonospora and Microbispora from soil. J. Ferment Bioeng., 72: 320-326.
CrossRefDirect Link - Hayakawa, M., T. Tamura and H. Nonomura, 1991. Selective isolation of Actinoplanes and Dactylosporangium from soil by using g-collidine as the chemoattractant. J. Ferment Bioeng., 72: 426-432.
CrossRef - Johanson, A. and B. Blazquez, 1992. Fungi associated with banana crown rot on field-packed fruit from the Windward Islands and assessment of their sensitivity to the fungicides thiabendazole, prochloraz and imazalil. Crop Prot., 11: 79-83.
CrossRef - Krauss, U., R. Bidwell and J. Ince, 1998. Isolation and preliminary evaluation of mycoparasites as biocontrol agents of crown rot of banana. Biol. Control, 13: 111-119.
CrossRef - Krauss, U. and W. Soberanis, 2001. Biocontrol of cocoa pod diseases with mycoparasite mixtures. Biol. Control, 22: 149-158.
CrossRef - Lorito, M., A. Di Pietro, C.K. Hayes, S.L. Woo and G.E. Harman, 1993. Antifungal, synergistic interaction between chitinolytic enzymes from Trichoderma harzianum and Enterobacter cloacae. Phytopathology, 83: 721-728.
CrossRefDirect Link - Otoguro, M., M. Hayakawa, T. Yamazaki and Y. Iimura, 2001. An integrated method for the enrichment and selective isolation of Actinokineospora spp. in soil and plant litter. J. Appl. Microbiol., 91: 118-130.
CrossRefDirect Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Smith, J., A. Putnam and M. Nair, 1990. In vitro control of Fusarium diseases of Asparagus officinalis L. with a Streptomyces or its polyene antibiotic, faeriefungin. J. Agric. Food Chem., 38: 1729-1733.
CrossRefDirect Link - Stevenson, I.L., 1956. Antibiotic activity of actinomycetes in soil as demonstrated by direct observation techniques. J. Gen. Microbiol., 15: 372-380.
PubMed - Taechowisan, T., C. Lu, Y. Shen and S. Lumyong, 2005. 4-Arylcoumarins from endophytics Streptomyces aureofaciens CMUAc130 and their antifungal activity. Ann. Microbiol., 55: 63-66.
Direct Link - Taechowisan, T., J.F. Peberdy and S. Lumyong, 2003. Isolation of endophytic actinomycetes from selected plants and their antifungal activity. World J. Microbiol. Biotechnol., 19: 381-385.
CrossRefDirect Link - Taechowisan, T. J.F. Peberdy and S. Lumyong, 2003. Chitinase production by endophytic Streptomyces aureofaciens CMUAc130 and its antagonism against phytopathogenic fungi. Ann. Microbiol., 53: 447-461.
Direct Link - Trejo-Estrada, S.R., A. Paszczynski and D.L. Crawford, 1998. Antibiotics and enzymes produced by the biocontrol agent Streptomyces violaceusniger YCED-9. J. Ind. Microbiol. Biotechnol., 21: 81-90.
CrossRefDirect Link - Trejo-Estrada, S.R., I.R. Sepulveda and D.L. Crawford, 1998. In vitro and in vivo antagonism of Streptomyces violaceusniger YCED9 against fungal pathogens of turfgrass. World J. Microbiol. Biotechnol., 14: 865-872.
CrossRefDirect Link - Valois, D., K. Fayad, T. Barasubiye, M. Garon, C. Dery, R. Brzezinski and C. Beaulieu, 1996. Glucanolytic actinomycetes antagonistic to Phytophthora fragariae var. rubi, the causal agent of raspberry root rot. Applied Environ Microbiol., 62: 1630-1635.
Direct Link - Benyagoub, M., N. Benhamou and O. Carisse, 1998. Cytochemical investigation of the antagonistic between a Microsphaeropsis sp. (isolate P130A) and Venturia inaequalis. Phytopathology, 88: 605-613.
CrossRef