
M. Miransari
Department of Soil Science, College of Agricultural Sciences, Shahed University, P.O. Box 18151/159, Tehran-Qom Highway, Tehran, Iran
D. Smith
Department of Plant Science, Macdonald Campus of McGill University, 21111 Lakeshore Road, Ste-Anne-de-Bellevue, Quebec, H9X 3V9, Canada
Biotechnology
Year: 2009 | Volume: 8 | Issue: 2 | Page No.: 270-275







ABSTRACT
Gibberellins are plant hormones, enhancing seed germination. The bacterium-to-plant signal, lipo-chitooligosaccharides (LCOs) or Nod factors, are of great importance for roots organogenesis and hence, nodule formation and N fixation. Hence, we hypothesized that LCOs like gibberellins may also enhance barley (Hordeum vulgare L.) germination. The objectives were to test the effects of gibberellins on barley germination and to test the hypothesis that LCOs may increase seed germination in barley. The concentrations, tested were 10-5 M for gibberellins and 10-6 M and 10-7 and 10-8 M LCOs. Although, gibberellins were able to numerically increase barley germination (up to 18%), the LCOs seemed to be more effective on barley germination as they significantly increased seed germination (up to 44%). Hence, the novel finding indicates that for LCOs may also be very effective on barley seed germination, through inducing morphogenesis and physiological changes in seeds. This finding can have very important agricultural implications.
PDF Abstract XML References Citation
How to cite this article
M. Miransari and D. Smith, 2009. Rhizobial Lipo-Chitooligosaccharides and Gibberellins Enhance Barley (Hordeum vulgare L.) Seed Germination. Biotechnology, 8: 270-275.
DOI: 10.3923/biotech.2009.270.275
URL: https://scialert.net/abstract/?doi=biotech.2009.270.275
DOI: 10.3923/biotech.2009.270.275
URL: https://scialert.net/abstract/?doi=biotech.2009.270.275
INTRODUCTION
The process of symbiosis between Bradyrhizobium japonicum and soybean (Glycine max L. Merr.) begins with the excretion of the plant-to-bacterium signal, genistein, which is a bacterial nod gene inducer (Miransari and Smith, 2007, 2008), followed by the production of the rhizobia-to-plant signal, lipo-chitooligosaccharides (LCOs), or Nod factors (Bai et al., 2002). Lipo-chitooligosaccharides are those molecules, responsible for morphogenesis changes in the roots and hence, nodule formation (Miransari et al., 2006; Khan et al., 2008).
The combination of the oligosaccharides of β-1,4-linked N-acetyl-o-glucosamine and of some functional groups form LCOs. According to Perret et al. (2000) the production of LCOs is related to some complicated biochemical processes taking place with the help of enzymes, produced and activated as a result of nod genes expression. Different strains of rhizobium produce various mixtures of LCO and the strain, Bradyrhizobium japonicum 532C, produces nod Bj V (C18:1; MeFuc) (Prithiviraj et al., 2000; Khan et al., 2008).
Submicromolar concentrations of LCOs may result in physiological changes in legumes (Goedhart et al., 2003) and non-legumes (Souleimanov et al., 2002; Prithiviraj et al., 2003). Although, it is not clear yet how plants perceive LCOs (Macchiavelli and Brelles-Marino, 2004), it has been suggested that there are more than one LCO perception system (Cullimore et al., 2001) including two different LCO binding sites for Medicago sp. called NFBS1 and NFBS2, with different affinities (Gressent et al., 1999).
The LCOs are able to form (Macchiavelli and Brelles-Marino, 2004) and deform root hairs (Miransari et al., 2006), depolarize membrane, alkalinize the internal and external cellular space, induce the expression of nod genes, alternate ion fluxes and form nodules (Broughton et al., 2000). It has also been reported that LCOs may enhance seed germination in some crops (Prithiviraj et al., 2003), though there has not yet been any report on the enhancing effect of LCOs on seed germination in barley.
Macchiavelli and Brelles-Marino (2004) have stated that treating seeds of Medicago trancatula with LCOs may more effectively increase the number of nodules than when roots are not treated and have attributed this to the existence of a perception system with a high affinity for LCO in the seed and embryo of M. trancatula.
Seeds of cereals include endosperm providing the necessary nutrients, carbohydrates and proteins for seed growth before the beginning of seedling photosynthesis. It is made of two parts, the central starchy part and the surrounding aleurone (Bosnes et al., 1992). The barley (Hordeum vulgare) endosperm becomes available to the embryo as a result of hydrolases production during germination (Jones and Jacobsen, 1991).
While, the plant hormone, gibberellins, stimulates the synthesis and excretion of these enzymes (mainly α-amylases) and hence seed germination, abscisic acid (ABA) adversely affects these processes (Jones and Jacobsen, 1991). Hence, barley aleurone is of great significance for the study of signal transduction pathways in response to the plant hormones, gibberellins and ABA and some of these pathways have yet to be elucidated (Ritchie and Gilroy, 1998).
Gibberellins are involved in the induction of several genes, necessary for the synthesis and secretion of α-amylase before germination. The embryo synthesizes gibberellins, residing in the aleurone (Appleford and Lenton, 1997), where they are programmed for the expression of genes, responsible for the synthesis and secretion of α-amylases, proteases and β-glucanases.
Although, there are findings on the effects of exogenously applied gibberellins on barley seed germination, but since to our knowledge there are not any findings on the effects of LCO on barley seed germination, we hypothesized that with respect to their properties, LCOs are also able to induce and enhance seed germination in barley. We proposed these experiments to further exhibit the great importance of microbiological products in the stability and production of ecosystems and indicate that legumes can also be very useful to non-legumes per se in a intercropping or rotational cropping. The objectives were (1) to test the effects of gibberellins on barley seed germination and (2) to test the hypothesis that LCO may increase seed germination in barley, though it may be concentration dependent.
MATERIALS AND METHODS
Two experiments were conducted in which the following stages were taken. Seeds of barley (Hordum vulgare L.) were surface sterilized in sodium hypochlorite (2% solution containing 4 ml L-1 Tween 20) and rinsed several times with distilled water (Bhuvaneswari et al., 1980). There were eight seeds per treatment in the first experiment and to increase the experimental precision 20 seeds per treatment in the second experiment. In both experiments seeds were grown in Petri dishes in the laboratory, at room temperature, with 16 h of fluorescence light in the month of October at the laboratory of Plant Science Department, Macdonald College of McGill University, Montreal, Canada.
In the first experiment distilled water was used as the control treatment and concentrations of 10-5 and 10-6 M of gibberellins (Sigma-Aldrich Canada Ltd., Oakville, Canada) and LCO (produced, isolated and purified as explained below) were applied to the seeds in Petri dishes (9 for each treatment) containing filter papers and 5 mL of the solution (Benech-Arnold et al., 2003).
In the second experiment also similar stages were taken but in addition to the treatments, used in the first experiment, concentrations of 10-7 and 10-8 M of LCO were also applied to ten Petri dishes for each treatment. Twenty hours after applying the treatments the seed germination rates were determined.
Production, extraction and purification of LCO: LCOs were produced, extracted and purified taking the following stages (Miransari et al., 2006).
Bacterial culture: Bradyrhizobium japonicum strain 532C was grown at 28°C in 100-125 mL of sterile yeast mannitol medium (YEM) (pH 6.8) containing 10 g mannitol, 0.5 g K2HPO4, 0.0977 g MgSO4, 0.1 g NaCl, 0.4 g yeast extract and 1000 mL distilled water. The culture was shaken at 150 rpm until it achieved an OD620 equal to 0.4-0.6 (4-6 days).
Thereafter, 2 L bacterial subculture was prepared by adding 5 mL of the culture to 250 mL of YEM medium and 0.25 mL of 50 μM genistein was added to each 250 mL of bacterial subculture. The subculture was grown at 28°C and shaken at 150 rpm until the OD620, reached 0.8-1.0 (5-7 days). Concentration of 5 μM genistein in the subculture was produced, while looking white-grayish. The LCO biosynthesis is induced by genistein over the course of 48-96 h.
Lipo-chitooligosaccharide extraction: To the bacterial subculture 0.8 L of HPLC grade 1-butanol was added and it was shaken for 5 min and then left over night to separate into organic and water phases. Using a separatory funnel the 2 layers were carefully separated. The lower layer was discarded and the upper one was collected and stored at 4°C before it was concentrated by evaporation. A rotary evaporator (Yamato RE500) and temperature of 80°C were used to evaporate the collected layer. The remaining material was dissolved in 4 mL of 18% acetonitrile and kept in the dark in a glass vial sealed with parafilm.
Purification of LCO by HPLC: For the HPLC analysis a Vydac C18 reversed-phase column (0.46x25 cm; 5 μM) with a flow rate of 1.0 mL min-1 and a Vydac guard column were used. To establish a baseline, 18% acetonitrile (AcN/H2O; w/w) was used and this baseline was run for at least 10 min. The baseline should be about 0.01. If the injection volume was greater than 200 μL, the auxiliary loop was installed for Waters 712 WISP Autosampler. After injecting the sample, isocratic elution by 18% of AcN for 45 min was started. This step removes all non-polar junk light fractions. Thereafter, gradient elution for 90 min was applied using 18-82% of AcN. LCO should begin to appear at the detector at 94-96 min after starting the run (identified by using a standard) (Souleimanov et al., 2002). When the water fraction collector (collects a new fraction every 2 min) was used, LCO was found in fractions, numbered 47 and 48.
Statistical analysis: Using the SAS (1988) system the analysis of variance was performed for the data and the effect of the model and the main effect in the model, treatments, were statistically determined. Mean values were compared using the GLM method and the least significant difference test at p = 0.05 (Steel and Torrie, 1980).
RESULTS AND DISCUSSION
In the first experiment, adding LCO at 10-6 M increased seed germination (by 44%) significantly, compared to the control, while adding gibberellins at 10-5 M did not. Both the effects of model and treatment were significant (p = 0.05) (Table 1).
In the second experiment, the effects of both the model and the treatment were also significant. In the second experiment, adding LCO at 10-7 M increased seed germination (by 25%) significantly compared to the control while adding gibberellins at 10-5 M or LCO at either 10-6 or 10-8 M did not. The effect of LCO 10-8 M was not different from control (Table 1).
The effects of LCOs on barley germination: Although, the key role of LCOs in nodule formation has been determined, there are other plant morphogenesis activities, attributed to LCOs including the stimulation of genes, involved in cell cycling in the cultures of suspension cell and stimulation of mitosis division in the cultures of protoplast in both legumes and non-legumes (Souleimanov et al., 2002) and also Ca2+ spiking in the cytoplasm (Mitra et al., 2004; Kalo et al., 2005). After 1 min of addition, Nod factors can depolarize the membrane of legume root hairs (Ehrhardt et al., 1992; Miwa et al., 2006), as a result of Ca2+ spiking at the cytoplasm of root hair tip (Cardenas et al., 1998; Shaw and Long, 2003).
| Table 1: | The effects of different concentrations of LCO and gibberellins on barley seed germination (± SD) in the first and second experiment |
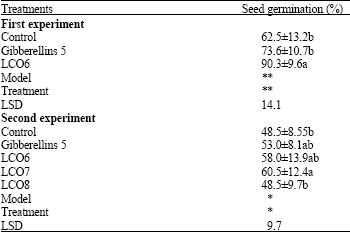 | |
| Gibberellins 5: Gibberellins at 10-5 M; LCO6, 7 and 8: LCO at 10-6, 10-7 and 10-8 M, respectively. LSD: Least significant difference test. Values within the same column, followed by different letter(s) are significantly different at p = 0.05; * = p<0.05; ** = p<0.01 | |
Present results show that LCOs are capable of enhancing barley seed germination, which is in agreement with Bai et al. (2002), who indicated that LCOs might induce seed germination in soybean. In the soil, LCOs may have many enhancing effects on plant growth through inducing seed germination (Zhang and Smith, 2001), enhancing plant growth, development and yield for both legumes and non-legumes, increasing the photosynthetic rates when sprayed onto the leaves (Smith et al., 2002; Almaraz et al., 2006), stimulating effects on flavonoid genes (Spaink and Lugtenberg, 1994), enhancing the ability of arbuscular mycorrhiza in root colonization (Xie et al., 1995) and cell cycling and embryogenesis. In addition LCO can also later the auxin transfer (Souleimanov et al., 2002), which its balance with cytokinin can affect plant growth (Relic et al., 1993). Although, it has yet to be specified whether there are LCOs receptors on the seed and embryos, however scientists have suggested that there are LCOs receptors on the roots of developing embryos (Macchiavelli and Brelles-Marino, 2004).
Based on present results, LCO 10-7 M may be the most effective concentration to induce seed germination in barley, which is in accordance with Zhang and Smith (2001), Smith et al. (2002) and Supanjani et al. (2005). This stimulating effect is also in agreement with the idea that LCOs are capable of adjusting plant growth through restoration and continuation of cell cycling and embryogenesis in mutated plants and cultures of somatic embryo (De Jong et al., 1993; Egertsdotter and Von Arnold, 1998) when the plant hormones, auxins and cytokinins are not present (Daychok et al., 2000).
Also, foliar application of LCOs at 10-6, 10-8 and 10-10 M onto the leaves increased the photosynthetic rates from 10 to 20% in soybean, common bean, maize (Khan et al., 2008), rice, canola, apple and grape plants. This enhanced photosynthetic rate resulted in an increase of 40% in soybean yield under field conditions (Smith et al., 2002). LCOs in rhizobia may be overexpressed by phenolic compounds, which are abundant in plant root exudates and residues (Dakora, 2003). When presented at high concentrations in the soil, these signal molecules may result in the production of large amounts of LCOs in the rhizosphere, with high inducing effects on plant growth (Dakora, 2003).
Gibberellins and barley germination: Present results show that exogenously applied gibberellins enhanced the germination of barley seeds, which is in agreement with Steinbach et al. (1997) and Benech-Arnold et al. (2000), who stated that exogenous gibberellins increased germination in developing seeds. Also, application of gibberellins may enhance seed germination in dormant seeds as the biosynthesis of gibberellins is suppressed in dormant seeds compared with non-dormant seeds (Perez-Flores et al., 2003). The balance between gibberellins and ABA is a determining factor in the germination capacity of seeds (Benech-Arnold et al., 2003).
To prove the hypothesis that gibberellins are necessary for the germination of immature embryo, White and Rivin (2000) applied paclobutrazol and ancymidol, the inhibitors of gibberellins synthesis, to the cultured embryo at different stages of development. The dissection of embryos took place at different stages and also the ratio and the part of the embryos, germinated contingently decreased. Addition of exogenous gibberellins could suppress the effects of gibberellins inhibitors enabling embryos to germinate. Also, when gibberellins inhibitors are not present, exogenous gibberellins could significantly enhance embryos germination.
CONCLUSION
With regard to the results of these experiments it is very much clear that LCOs are able to enhance barley seed germination even more effectively than gibberellins through morphogenesis and physiological changes. This can indicate the great importance of biological (i.e., secondary metabolites) products in enhancing barley seed germinating and growth. Although, barely is a non-legume plant, however the legume LCOs products in a rotation or intercropping (Prithiviraj et al., 2003; Khan et al., 2008) can very much contribute to the increased level of productivity (Long, 2005). It should also be mentioned that to our knowledge data regarding the effects of LCOs and barley germination are rare.
REFERENCES
- Almaraz, J., X. Zhou, A. Souleimanov and D.L. Smith, 2006. Gas exchange characteristics and dry matter accumulation of soybean treated with Nod factors. J. Plant Physiol., 164: 1391-1393.
Direct Link - Appleford, N.E.J. and J.R. Lenton, 1997. Hormonal regulation of α-amylase gene expression in germinating wheat (Triticum aestivum) grains. Physiol. Plantarum, 100: 534-542.
CrossRefDirect Link - Bai, Y., A. Souleimanov and D.L. Smith, 2002. An inducible activator produced by a Serratia proteamaculans strain and its soybean growth-promoting activity under greenhouse conditions. J. Exp. Bot., 53: 1495-1502.
CrossRefDirect Link - Benech-Arnold, R.L., S. Enciso, R.A. Sanchez, F. Carrari and L. Perez-Flores et al., 2000. Involvement of ABA and GAs in the Regulation of Dormancy in Developing Sorghum Seeds. In: Seed Biology: Advances and Applications, Black, M., K.J. Bradford and J. Vazquez Ramos (Eds.). CAB International, Wallingford, UK., pp: 101-111.
- Benech-Arnold, R.L., S. Enciso, R.A. Sanchez and M.V. Rodriguez, 2003. On the hormonal nature of the stimulatory effect of high incubation temperatures on germination of dormant sorghum (S. bicolor) caryopses. New Phytol., 160: 371-377.
Direct Link - Broughton, W.J., S. Jabbouri and X. Perret, 2000. Keys to symbiotic harmony. J. Bacteriol., 182: 5641-5652.
CrossRefDirect Link - Dakora, F., 2003. Defining new roles for plant and rhizobial molecules in sole and mixed plant cultures involving symbiotic legumes. New Phytol., 158: 39-49.
Direct Link - Daychok, J.V., A.E. Tobin, N.P.J. Price and S. von Arnold, 2000. Rhizobial Nod factors stimulate somatic embryo development in Picea abies. Plant Cell Rep., 19: 290-297.
Direct Link - Ehrhardt, D.W., E.M. Atkinson and S.R. Long, 1992. Depolarization of alfalfa root hair membrane potential by Rhizobium meliloti Nod factors. Science, 256: 998-1000.
Direct Link - Goedhart, J., J.J. Bono, T. Bisseling and Jr. T.W. Gadella, 2003. Identical accumulation and immobilization of sulfated and non-sulfated Nod factors in host and non-host root hair cell walls. Mol. Plant Microbe Interact., 16: 884-892.
Direct Link - Gressent, F., S. Drouillard and N. Mantegazza, 1999. Ligand specificity of a high-affinity binding site for lipo-chitooligosaccharidic Nod factors in Medicago cell suspension cultures. Proc. Natl. Acad. Sci. USA., 96: 4704-4709.
CrossRefDirect Link - Jones, R.L. and J.V. Jacobsen, 1991. Regulation of synthesis and transport of secreted proteins in cereal aleurone. Int. Rev. Cytol., 126: 49-88.
CrossRefDirect Link - Kalo, P., C. Gleason, A. Edwards, J. Marsh and R.M. Mitra et al., 2005. Nodulation signaling in legumes requires NSP2, a member of the GRAS family of transcriptional regulators. Science, 308: 1786-1789.
Direct Link - Khan, W., B. Prithiviraj and D.L. Smith, 2008. Nod factor [Nod Bj V (C18:1, MeFuc)] and lumichrome enhance photosynthesis and growth of corn and soybean. J. Plant Physiol., 185: 1342-1351.
Direct Link - Macchiavelli, R.E. and G. Brelles-Marino, 2004. Nod factor-treated Medicago truncatula roots and seeds show an increased number of nodules when inoculated with a limiting population of Sinorhizobium meliloti. J. Exp. Bot., 55: 2635-2640.
Direct Link - Miransari, M., D.L. Smith, A.F. Mackenzie, H.A. Bahrami, M.J. Malakouti and F. Rejali, 2006. Overcoming the stressful effect of low pH on soybean root hair curling using lipochitooligosaccahrides. Commun. Soil Sci. Plant Anal., 37: 1103-1110.
CrossRef - Miransari, M. and D.L. Smith, 2007. Overcoming the stressful effects of salinity and acidity on soybean [Glycine max (L.) Merr.] nodulation and yields using signal molecule genistein under field conditions. J. Plant Nutr., 30: 1967-1992.
CrossRefDirect Link - Miransari, M. and D. Smith, 2008. Using signal molecule genistein to alleviate the stress of suboptimal root zone temperature on soybean-Bradyrhizobium symbiosis under different soil textures. J. Plant Interact., 3: 287-295.
CrossRefDirect Link - Mitra, R.M., C.A. Gleason, A. Edwards, J. Hadfield, J.A. Downie, G.E.D. Oldroyd and S.R. Long, 2004. A Ca2+/calmodulin-dependent protein kinase required for symbiotic nodule development: Gene identification by transcript-based cloning. Proc. Natl. Acad. Sci. USA., 101: 4701-4705.
Direct Link - Miwa, H., J.E.D. Jongho, G.E.D. Oldroyd and J.A. Downie, 2006. Analysis of calcium spiking using a cameleon calcium sensor reveals that nodulation gene expression is regulated by calcium spike number and the developmental status of the cell. Plant J., 48: 883-894.
Direct Link - Perret, X., C. Staehelin and W.J. Broughton, 2000. Molecular basis of symbiotic promiscuity. Microbiol. Mol. Biol. Rev., 64: 180-201.
Direct Link - Perez-Flores, L., F. Carrari, R. Osuna-Fernandez, M.V. Rodriguez and S. Enciso et al., 2003. Expression analysis of a GA 20-oxidase in embryos from two sorghum lines with contrasting dormancy: Possible participation of this gene in the hormonal control of germination. J. Exp. Bot., 390: 2071-2079.
Direct Link - Prithiviraj, B., A. Souleimanov, X. Zhou and D.L. Smith, 2000. Differential response of soybean (Glycine max (L.) Merr.) genotypes to lipo-chitin-oligosacchride Nod Bj V (C18:1 MeFuc). J. Exp. Bot., 51: 2045-2051.
CrossRefDirect Link - Prithiviraj, B., X. Zhou, A. Souleimanov, W.M. Kahn and D.L. Smith, 2003. A host-specific bacteria-to-plant signal molecule (Nod factor) enhances germination and early growth of diverse crop plants. Planta, 21: 437-445.
Direct Link - Relic, B., F. Talmont, J. Korsinska, W. Golonowski, J.C. Prome and W.J. Broughton, 1993. Biological activity of Rhizobium sp. NGR234 Nod-factors on Macroptilium atropurpureum. Mol. Plant Microbe Interact., 6: 764-774.
Direct Link - Ritchie, S. and S. Gilroy, 1998. Gibberellins: regulating genes and germination. New Phytol., 140: 363-383.
CrossRefDirect Link - Shaw, S.L. and S.R. Long, 2003. Nod factor elicits two separable calcium responses in Medicago truncatula root hair cells. Plant Physiol., 13: 976-984.
Direct Link - Souleimanov, A., B. Prithiviraj and D.L. Smith, 2002. The major Nod factor of Bradyrhizobium japonicum promotes early growth of soybean and corn. J. Exp. Bot., 53: 1929-1934.
Direct Link - Spaink, H.P. and B.J.J. Lugtenberg, 1994. Role of rhizobial lipo-chitin oligosaccharide signal molecules in root nodule organogenesis. Plant Mol. Biol., 26: 1413-1422.
PubMed - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Steinbach, H.S., R.L. Benech-Arnold and R.A. Sanchez, 1997. Hormonal regulation of dormancy in developing Sorghum seeds. Plant Physiol., 113: 149-154.
CrossRefDirect Link - Supanjani, F. Mabood, A. Souleimanov, K.D. Lee and D.L. Smith, 2005. Stability and activity of the major nod factor produced by Bradyrhizobium japonicum following purification, sterilization and storage. Crop Sci., 45: 1281-1285.
Direct Link - White, C.N. and C.J. Rivin, 2000. Gibberellins and seed development in maize. II. Gibberellin synthesis inhibition enhances abscisic acid signaling in cultured embryos. Plant Physiol., 122: 1089-1098.
Direct Link - Xie, Z.P., C. Staehelin, H. Vieheilig, A. Wiemken and S. Jabbouri et al., 1995. Rhizobial nodulation factors stimulate mycorrhizal colonization of nodulating and nonnodulating soybean. Plant Physiol., 108: 1519-1525.
PubMedDirect Link - Zhang, F. and D.L. Smith, 2001. Interorganismal signaling in suboptimum environments: The legume-rhizobia symbiosis. Adv. Agron., 76: 125-161.
CrossRefDirect Link
murudkar smita r Reply
great job
lakshmi Reply
please tell physiological change in seed germination
kanti priy bhardwaj Reply
this is a great artical.
Prof. Dr. M. Miransari Reply
Thank you very much for your great comments. The physiological changes during seed germination are mostly related to the enzymes activities changes and nutrients availability in the seed.
Prof. Dr. Mohammad Miransari Reply
I will be happy to respond to the other comments too.
With respect,
Prof. Dr. Mohammad Miransari