
Reza Halabian
Department of Animal Science, Faculty of Agriculture, University of Zanjan, Zanjan, Iran
Morad Pasha Eskandari Nasab
Department of Animal Science, Faculty of Agriculture, University of Zanjan, Zanjan, Iran
Mohammad Reza Nassiry
Center of Excellence in Animal Science, Faculty of Agriculture, Ferdowsi University of Mashhad, Mashhad, Iran
Ali Reza Heravi Mossavi
Center of Excellence in Animal Science, Faculty of Agriculture, Ferdowsi University of Mashhad, Mashhad, Iran
Seyed Abolfazl Hosseini
Department of Biology, Faculty of Science, STT University, Sabzevar, Iran
Saber Qanbari
Department of Animal Science, Faculty of Agriculture, University of Zanjan, Zanjan, Iran
Biotechnology
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 118-123







ABSTRACT
In this study, DNA isolated from blood samples of Iranian Holstein cows (N = 120) and PCR products (156 bp) were analyzed by SSCP technique with MDE gel. Then 4 patterns (Z1, Z2, ...Z4) of provided by SSCP were sequenced and then using POLYPHRED 5.04 software, 4 SNPs were detected in positions 6237, 6263, 6268 and 6297. The genotypes were TC and TG in position 6237, TT in position 6263, AG in position 6268 and AC in position 6297. From of detected SNPs, they that located in positions 6237 and 6268 caused change in the amino acids sequence of protein. These data provide evidence that bPRL gene is a good polymorphic source for single nucleotides that can be used for association with milk yield and investigate whether mutations in this sequences might be responsible for quantitative variations in milk yield and composition.
PDF Abstract XML References Citation
How to cite this article
Reza Halabian, Morad Pasha Eskandari Nasab, Mohammad Reza Nassiry, Ali Reza Heravi Mossavi, Seyed Abolfazl Hosseini and Saber Qanbari, 2008. Characterization of SNPs of Bovine Prolcatin Gene of Holstein Cattle. Biotechnology, 7: 118-123.
DOI: 10.3923/biotech.2008.118.123
URL: https://scialert.net/abstract/?doi=biotech.2008.118.123
DOI: 10.3923/biotech.2008.118.123
URL: https://scialert.net/abstract/?doi=biotech.2008.118.123
INTRODUCTION
Prolactin is a polypeptide hormone that is synthesized in and secreted from specialized cells of the anterior pituitary gland, the lactotrophs (Bole-Feysot et al., 1998). The varied effects of prolactin on mammary gland include growth and development of the mammary gland (mammogenesis), synthesis of milk (lactogenesis), maintenance of milk secretion (galactopoiesis), expression milk protein genes, lactose and lipid (Bern and Nicol, 1968; Akers et al., 1981; Mitra et al., 1993; Dybus, 2002). However, has been shown that prolactin has over 300 separate biological activities (Bole-Feysot et al., 1998). Indeed, not only does prolactin subserve multiple roles in reproduction other than lactation, but it also plays multiple homeostatic roles in organism (Grosvenor and Whitworth, 1974; Bern, 1975; Shennan, 1994; Buskila and Shoenfeid, 1996; Goffin et al., 1998; Neidhart, 1998).
The detection and genotyping of sequence variations is at the core of all genetic analysis and all approaches to disease gene mapping ultimately lead to variation discovery across a candidate region or gene(s) to identify genetic variants that affect the trait of interest (Nickerson et al., 1997).
SNPs are considered to be useful polymorphic and ideal markers for genetic studies of polygenic traits and a worldwide effort to collect SNPs has achieved an accumulation of millions of them in public databases (Tomonari et al., 2001; Jalving et al., 2004). SNPs are most abundant type of polymorphism in the genome. Therefore, SNPs are becoming the preferred markers for high density mapping (Vignal et al., 2002).
In human, agricultural (Bovethuts and Weller, 1994; Rothschild et al., 1994) and experimental populations (Mackay and Langley, 1990), polymorphisms at candidate genes have been associated with quantitative genetic variation. Therefore, bPRL gene can be an important and excellent candidate for linkage analysis with Quantitative Trait Loci (QTL) affecting milk production traits (Brym et al., 2005) and for mapping hSNP (haplotype SNP) and their association with milk performance trait.
The bPRL found on the chromosome 23, in the bovine genome (Hallerman et al., 1998). The bPRL gene is about 10 kb in size and is composed of 5 exons and 4 introns (Camper et al., 1984). This encodes the 199 amino acids mature protein (Wallis, 1974). In relation to survey of polymorphisms of bPRL gene, several study have been carried out by SSCP and PCR-RFLP methods (Cowan et al., 1989; Hart et al., 1993; Zhang et al., 1994; Mitra et al., 1993; Chung et al., 1996; Chrenek et al., 1999; Udina et al., 2001). By analysis of four different mRNA clones, four silent nucleotide substitutions were identified (Sasavage et al., 1982). On the basis of sequencing of PCR product representing different SSCP pattern, six SNPs were detected (Brym et al., 2005). Also by using Milkprotchip, 5 SNPs in bPRL gene were genotyped (Kaminski et al., 2005).
The aim of the present study was detection and genotyping of SNPs of bPRL gene by SSCP method with MDE gel, sequencing of different SSCP patterns and analysis by POLYPHRED software.
MATERIALS AND METHODS
Sampling and DNA extraction: Blood Samples (N = 120) were collected from Iranian Holstein cattles in Astan Ghods farm located at Mashhad. Whole blood (100 μL) was used as source for DNA, which was extracted by a modified Guanidine Isothiocyanate-Silica gel method (Boom et al., 1989). The concentration and purity of the DNA obtained were assessed by spectrophotometery and electrophoresis in 1% agarose gels, respectively.
PCR amplification: Polymerase Chain Reaction (PCR) was carried out with Genepak PCR universal kit that was containing 10 μL PCR diluents, 3 μL mixture of primers (5 pm μL-1), 6 μL of ddH2O and 1 μL of genomic DNA. To detect of SNPs of the bPRL gene, a fragment of exon 3 (156 bp) was amplified with primers (PRL-Forward; 5´-CGAGTCCTTATGAGCTTGATTCT-3` and PRL-Reverse; 5`-GCCTTCCAGAAGTCGTTTTT-3). PCR with primers was performed under following condition: initial denaturation (94°C for 5 min), 30 cycles of denaturation (94°C for 30 sec), annealing (59°C for 40 sec), extension (72°C for 20 sec) and final synthesis (72°C for 3 min). The yield and specificity of PCR products were evaluated after electrophoresis in 2% agarose gel with ethidium bromide.
SSCP-MDE analysis: In order to detect mutations, SSCP analysis (Orita et al., 1989; Travis, 1995) with MDE® gel solution (Camberx Bio Science Rockland) was performed according to the guidelines described by Camberx Bio Science Rockland. Briefly, 2.0 μL of PCR product was mixed with 18 μL of the denaturing solution (EDTA 0.5 molar, 95% formamide, 0.25% bromophenol blue and 0.25% xylenecyanol), then denatured for 5 min at 95°C, rapidly quenching on ice and, then 4 μL of sample loaded on precast gels (20-25 cm, 0.5 mm) containing 20 mL 2X MDE gel solution, 4.8 mL 10X TBE buffer, 55.2 mL ddH2O, polymerized by the addition of 500 μL of 10% w/v ammonium persulfate and 50 μL TEMED. The gel was run at a constant power of 6W at room temperature for 8 h. The gels were stained by sliver staining method (Benboza et al., 2006).
Sequencing and sequencing analysis: PCR products representing different SSCP were sequenced with an ABI 3730 XL sequencer. All sequences were base-called and assembled using the phred/phrap/consed (Ewing et al., 1998; Ewing and Green, 1998; Gordon et al., 1998) suit of programs. A related programs polyphred version 5.04 (Nickerson et al., 1997) was used to identify SNPs in the assembled sequences. Differences between individual PCR products as well as between PCR products and reference sequences available in GenBank (AF 426315) classified as SNPs.
RESULTS AND DISCUSSION
To detect of SNPs within bPRL gene, first we obtained a specific PCR product of expected size (156 bp) with high quality as shown by agarose gel electrophoresis (Fig. 1). This PCR product was consist of whole exon 3 and part of introns 2 and 3.
We used MDE-SSCP method in order to initial screening for polymorphism in 2 herds of Iranian Holstein cattle (N = 120). Four different SSCP patterns were observed: Z1, Z2... Z4 (Fig 2). The frequencies for patterns have been understood in Table 1, that Z1 pattern had higher frequency (0.68) than others.
After direct sequencing of 4 patterns of SSCP and analysis of assembled sequences by polyphred 5.04 revealed 4 SNPs (Table 2). Genotypes of each SNP correspond to SSCP patterns have shown in Table 3. The allelic frequencies for 4 SNPs have shown in Table 4, that G allele had lower frequency (0.0167) than others. Also part of results of direct sequencing that presented SNPs in the bPRL gene in different SSCP patterns in (Fig. 3).
| Table 1: | Frequencies of SSCP patterns in the Iranian Holstein cattle breed |
 | |
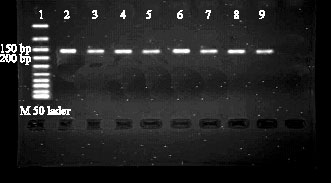 | |
| Fig. 1: | Electrophoregram in 2% agarose gel. Lane 1 is ladder M 50 and lanes 2-9 are 156 bp segment of the bPRL gene consist of whole exon 3 and part of introns 2 and 3 |
| Table 2: | Positions and properties of identified SNPs in this study |
 | |
| Table 3: | Genotypes of identified SNPs within PCR products representing SSCP patterns using resequencing and polyphred 5.04 software |
| |
| Table 4: | Allelic frequencies of detected SNPs |
 | |
 | |
| Fig. 2: | SSCP analysis of a 156-bp PCR product of the bPRL gene with MDE gel. Four SSCP patterns are visible: 1, Z2...Z4 |
SNPs in position 6237, 6263 and 6268 are located within exon 3 of bPRL (cSNP), that SNPs located in positions 6363 was silent with respect to amino acids sequence of the protein, but SNP located in positions 6237 and 6268 caused change in the amino acids sequence of protein as shown in Table 5. Also SNP in positions 6297 are located in intron3.
| Table 5: | List of changed amino acids correspond to detected SNPs |
 | |
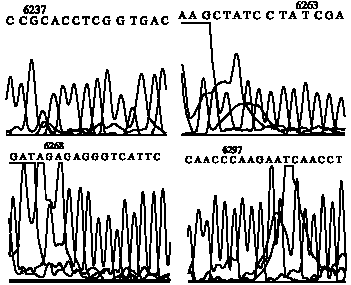 | |
| Fig. 3: | Result of direct sequencing of SSCP different patterns of a 156 bp fragment of the bPRL gene, showing four SNPs at positions: 6237, 6263, 6268 and 6297 that detected by POLYPHRED 5.04 software |
Because of bPRL plays a crucial role in mammary gland development, in the initiation and maintenance of lactation and because of milk production is under the indirect influence of it, thus we decide to detection SNPs of bPRL gene for next study in regard of their association with milk performance traits.
Since, MDE gel solution is very sensitive and efficient at detecting sequence polymorphisms in PCR amplicon and because MDE gel solution causes DNA separation to occur on the basis of both size and conformation, thus increasing the probability of detecting differences from as low as 15% in standard polyacrylamide gels, to approximately 80%; therefore, we used it in stand of standard polyacrylamide gel (Soto and Sukumar, 1992; Martins-Lopes et al., 2001).
With its decreasing costs and rapidly-expanding scale, DNA sequencing seems likely to play a key role in genetic analysis. As studies identify genomic regions associated with biologically or medically important phenotypes, this will inevitably be followed by sequencing of these regions in appropriate individuals.
Also because of resequencing of genes to identify DNA variations play a major role in the post-genomics analysis of animal and human biology, we resequenced different patterns of SSCP and analyzed their with POLYPHRED 5.04 software that automatically detects the presence of heterozygous single nucleotide substitutions by fluorescence based sequencing PCR product. POLYPHRED 5.04 focuses particularly on detecting SNPs through the identification of heterozygous individuals and is therefore well suited to SNP detecting in diploid samples obtained following DNA amplification. It is substantially more accurate than existing approaches and, importantly, provides a useful quantitative measure of its confidence in each potential SNP detected and in each genotype called. The essence of SNP identification is elimination of sequence errors and misaligned bases. therefore, because of quality bases were available, we used polyphred version 5.04. POLYPHRED was shown to be successful in this approach (Nickerson et al., 1997).
Within the bPRL gene several polymorphisms have been reported but most papers only have described RFLP and SSCP mutations without explaining its nature and locations. Zhang et al. (1994) observed three patterns by SSCP analysis of a 857 bp PCR segment of the bPRL gene consist of exon 1 and intron 1. Also Brym et al. (2005) reported three patterns by SSCP-PCR on exon 4 of the bPRL gene. Hart et al. (1993) observed a four-allele SSCP in the bPRL gene 5` flank. Among of all SNPs identified in earlier studies, SNP located in position 8398 of the bPRL (in exon 4) that recognizable by Rsa I endonuclease, has become a popular genetic marker for genetic characterization of cattle populations by mean of PCR-RFLP (Mitra et al., 1993; Chrenek et al., 1998; Brym et al., 2005). Udina et al. (2001) and Dybus (2002) analysed a 156 bp fragment of exon 3 of the bPRL gene by PCR-RFLP with Rsa I, but did not presented position of SNP. Because of this work with these techniques (MDE-SSCP, direct sequencing and different method of detection and genotyping SNPs by polyphred 5.04 software) was carried out for first time on exon 3 of bPRL gene, therefore we could not compared our results with others.
It is noteworthy that SNPs identified in this work can compose intragenic haplotypes (at least two SNPs within a single locus). Because of the limited number of animals genotyped, we were able to show only two haplotypes (Table 3). Intragenic haplotypes can be used as more informative markers in polygenic and association studies. Such linked SNPs may be used in the near future for construction of a bovine haplotype map applied in a new strategy of QTL mapping.
Present finding reveals that bPRL gene is a good polymorphic source for single nucleotides, that opens interesting prospects for future selection programs, specially marker-assistant selection for milk production traits.
ACKNOWLEDGMENTS
This research was carried out in Laboratory of Animal Biotechnology of Center of Excellence in Animal Science of Ferdowsi University of Mashhad.
ABBREVIATIONS
| MDE | : | Mutation Detection Enhancement. |
| SNP | : | Single Nucleotide Polymorphism. |
| SSCP | : | Single Strand Conformation Polymorphism. |
| bPRL | : | Bovine prolactin. |
| cSNP | : | Coding SNP. |
REFERENCES
- Akers, R.M., D.E. Bauman, A.V. Capuco, G.T. Goodman and H.A. Tucker, 1981. Prolactin regulation of milk secretion and biochemical differentiation of mammary epithelial cells in periparturient cow. Endocrinology, 109: 23-30.
CrossRefPubMedDirect Link - Benbouza, H., J.M. Jacquemin, J.P. Baudoin and G. Mergeai, 2006. Optimization of a reliable, fast, cheap and sensitive silver staining method to detect SSR markers in polyacrylamide gels. Biotechnol. Agron. Soc. Environ., 10: 77-81.
Direct Link - Bole-Feysot, C., V. Goffin, M. Edery, N. Binart and P. Kelly, 1998. Prolactin (PRL) and its receptor: Action, signal transduction pathways and phenotype observed in PRL receptor Knockout mice. Endocrine Rev., 19: 225-268.
Direct Link - Boom, R., C.J. Sol, M.M. Salimans, C.L. Jansen, P.M. Wertheim-van Dillen and J. van der Noordaa, 1990. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol., 28: 495-503.
PubMedDirect Link - Bovenhuis, H. and J.I. Weller, 1994. Mapping and analysis of dairy cattle quantitative trait loci by maximum likelihood methodology using milk protein genes as genetic markers. Genetics, 136: 267-280.
PubMedDirect Link - Brym, P., S. Kaminski and E. Wojcik, 2005. Nucleotide sequence polymorphism within exon 4 of the bovine prolactin gene and its associations with milk performance traits. J. Applied Genet., 46: 179-185.
PubMedDirect Link - Buskila, D. and Y. Shoenfeld, 1996. Prolactin, bromocriptine and autoimmune diseased. Isr J. Med. Sci., 32: 23-27.
PubMedDirect Link - Camper, S.A. D.N. Luck, Y. Yao, R.P. Woychik, R.G. Goodwin, R.H.L. Jr. Lyons and F.M. Rottman, 1984. Characterization of the bovine prolactin gene. DNA., 3: 237-249.
Direct Link - Chung, E. and W. Kim, 1997. DNA polymorphism of prolactin gene in dairy cattle. Korean J. Dairy sci., 19: 105-112.
Direct Link - Chrenek, P., D. Vasicek, M. Bauerova and J. Bulla, 1998. Simultaneous analysis of bovine growth hormone and prolactin alleles by multiplex PCR and RFLP. Czech J. Animal Sci., 43: 53-55.
Direct Link - Cowan, C.M., M.R. Dentine, R.L. Ax and L.A. Schuler, 1989. Restriction fragment length polymorphisms associated with growth hormone and prolactin genes in Holstein bulls: Evidence for a novel growth hormone allele. Animal Genet., 20: 157-165.
CrossRefDirect Link - Ewing, B., L. Hillier, M.C. Wendi and P. Green, 1998. Base-calling of automated sequencer traces using phred I. Accuracy assessment. Genome Res., 8: 175-185.
CrossRefDirect Link - Goffin, V., B. Bouchard, C.J. Ormandy, E. Weimann and F. Ferrag et al., 1998. Prolactin: A hormone at the crossroads of neuroimmunoendocrinology. Ann. NY. Acad. Sci., 840: 498-509.
CrossRefDirect Link - Gordon, D., C. Abajian and P. Green, 1998. Consed: A graphical tool for sequence finishing. Genome Res., 8: 195-202.
CrossRefDirect Link - Grosvenor, C. and N. Whitworth, 1974. Evidence for a steady rate of secretion of prolactin following suckling in the rat. J. Dairy Sci., 57: 900-904.
CrossRefDirect Link - Hallerman, E.M., J.L. Theilmann, J.S. Beckmann, M. Soller and J.E. Womack, 1998. Mapping of bovine prolactin and rhodopsin genes in hybrid somatic cells. Anim. Genet., 19: 123-131.
CrossRefDirect Link - Hart, G.L., J. Bastiaansen, M.R. Dentine and B.W. Kirkpatrick, 1993. Detection of a four-allele single strand conformation polymorphism (SSCP) in the bovine prolactin gene 5' flank. Anim. Genet., 24: 149-149.
CrossRefDirect Link - Hayashi, K. and D. Yandell, 1993. How sensitive is PCR-SSCP? Hum. Mutat., 2: 338-346.
CrossRefPubMedDirect Link - Horseman, N., W. Zhao, E. Montecino‐Rodriguez, M. Tanaka, K. Nakashima and S. Engle et al., 1997. Defective mammopoiesis, but normal hematopoiesis, in mice with a targeted disruption of the prolactin gene. EMBO J., 16: 6926-6935.
CrossRefDirect Link - Jalving, R., R. Van't Slot and B.A. Van Oost, 2004. Chiken single nucleotide polymorphism identification and selection for genetic mapping. Poultry Sci., 83: 1925-1931.
Direct Link - Kaminski, S., A. Ahman, A. Ruse, E. Wojcik and T. Nualewski, 2005. MilkProtChip-a microarray of SNPs in candidate genes associated with milk protein biosynthesis-development and validation. J. Applied Genet., 46: 45-58.
PubMedDirect Link - Lefcourt, A.M., R.M. Akers, D.L. Wood and J. Bitman, 1994. Circadian and ultradian rhythms of peripheral prolactin concentrations in lactating dairy cows. Am. J. Phsiol. Regulatory Integrat. Comp. Physiol., 297: 1461-1466.
Direct Link - Mackay, T.F. and C.H. Langley, 1990. Molecular and phenotypic variation in the achaete-scute region of Drosophila melanogaster. Nature, 348: 64-66.
CrossRefDirect Link - Martins-Lopes, P., H. Zhang and R. Koebner, 2001. Detection of single nucleotide mutations in wheat using single strand conformation polymorphism gels. Plant Mol. Biol. Report, 19: 159-162.
Direct Link - Mitra, A., P. Schlee, C.R. Balakrishnan and F. Pirchner, 1993. Polymorphisms at growth-hormone and prolactin loci in Indian cattle and buffalo. J. Anim. Breed. Genet., 112: 71-74.
CrossRefDirect Link - Neidhart, M., 1998. Prolactin in autoimmune diseases. Proc. Soc. Exp. Biol., 217: 408-419.
Direct Link - Nickerson, D.A., V.O. Tobe and S.L. Taylor, 1997. Polyphred: Automating the detection and genotyping of single nucleotide substitutions using fluorescence-based resequencing. Nucleic Acids Res., 25: 2745-2751.
CrossRefDirect Link - Sasavage, N.L., J.H. Nilson, S. Horowitz and F.M. Rottman, 1982. Nucleotide sequence of bovine prolactin messenger RNA. Evidence for sequence polymorphism. J. Biol. Chem., 257: 678-681.
Direct Link - Shennan, D., 1994. Regulation of water and solute transport across mammalian plasma cell membranes by prolactin. Dairy Res., 61: 155-166.
PubMed - Soto, D. and S. Sukumar, 1992. Improved detection of mutations in the P53 gene in human tumors as single-stranded conformation polymorphs and double-stranded heteroduplex DNA. PCR Methods Applied., 2: 96-98.
CrossRefDirect Link - Tomonari, S., T. Tahira, A. suzuki, A. Higasa, K. Kukita, S. Baba and K. Hayashi, 2001. Precise estimation of allele frequencies of single nucleotide polymorphism by a quantitative SSCP analysis of pooled DNA. Am. J. Hum. Genet., 68: 214-218.
Direct Link - Udina, I., S. Turkova, M. Kostuchenko, L. Lebedeva and G. Sulimova, 2001. Polymorphism of bovine prolactin gene, microsatellites, PCR-RFLP. Russian J. Genet., 4: 407-411.
Direct Link - Vignal, A., D. Milan, M. Sancristobal and A. Eggen, 2002. A review on SNP and others marker and their use in animal genetics. Genet. Sel. Evol., 34: 275-305.
Direct Link - Zhang, H., S. DeNise and R. Ax, 1994. Rapid communication: Diallelic single-stranded conformational polymorphism detected in the bovine prolactin gene. J. Anim. Sci., 72: 256-256.
Direct Link - Orita, M., H. Iwahana, H. Kanazawa, K. Hayashi and T. Sekiya, 1989. Detection of polymorphisms of human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc. Natl. Acad. Sci. USA., 86: 2766-2770.
PubMedDirect Link