
Gamal Enan Enan
King Khalid Military Academy, Saudi Arabia
Biotechnology
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 143-147







ABSTRACT
Nisin or plantaricin UG1 induced a listericidal action in vitro and in food followed by regrowth of the target cells. The regrowing cells of Listeria monocytogenes LMG10470 were proved to be resistant variants at a resistance frequency from 10-5 to 10-6. The mix of nisin and plantaricin UG1 avoided the regrowth of becteriocin resistant cells of L. monocytogenes LMG10470 in vitro and in food. No cross resistance was found between nisin and plantaricin UG1 as Listerial cells resistant to one bacteriocin were sensitive to the other one; indicating on a synergistic listericidal effect of nisin-plantaricin UG1 mixture which could extend the shelf life of dairy products, meat products and tomato paste.
PDF Abstract XML References
How to cite this article
Gamal Enan Enan, 2006. Control of the Regrowing Bacteriocin Resistant Variants of Listeria monocytogenes LMG 10470 in vitro and in Food by Nisin-plantaricin UG1 Mixture. Biotechnology, 5: 143-147.
DOI: 10.3923/biotech.2006.143.147
URL: https://scialert.net/abstract/?doi=biotech.2006.143.147
DOI: 10.3923/biotech.2006.143.147
URL: https://scialert.net/abstract/?doi=biotech.2006.143.147
INTRODUCTION
Listeria monocytogenes is a gram positive rod that is catalase positive and shows a characteristic tumbling motility. Listeria monocytogenes (L. monocytogenes) has long been recognized as a veterinary pathogen and in humans causes a disease known as listeriosis in pregnant women and neonates (Pucci et al., 1988). It is found in soil, decaying animals and dead vegetation and in intestinal tracts of over domestic and wild species of birds and animals including sheep, cattle, chickens and swine (Moura et al., 1993). The ability of listerias cells to grow at temperatures ranging from 1 to 45°C, their high salt tolerance and their ability to initiate growth at a relatively low pH make these pathogens particularly difficult to control in food (Vignolo et al., 2000). A novel approach to the control of L. monocytogenes in food is the use of antimicrobial proteins (bacteriocins) from lactic acid bacteria (Nettles and Barefoot, 1993). However, listerias cells can develop tolerance towards nisin and pediocin like bacteriocins in vitro and in foods (Rekhif et al., 1994). An improved control of the target organisms and inhibition of bacteriocin resistant strains and species can be obtained by using a combination of one or more bacteriocins (Vignolo et al., 2000).
The bacteriocin plantaricin UG1 produced by Lactobacillus plantarum UG1 (Enan et al., 1994a, b) inhibited L. monocytogenes in vitro and in food (Enan et al., 1996; Enan, 2002). However, few regrowth was observed thereafter (Enan, 2002). The regrowing listerias cells were characterized to be resistant variants at a resistance frequency around 10-6 to control the regrowth of bacteriocin resistant cells of L. monocytogenes in food system, combined effect of bacteriocins is important (Bouttefroy and Milliere, 2000). The present research was undertaken to study the combined effect of nisin and plantericin UG1 on the growth of the plantaricin UG1-resistant variants of L. monocytogenes LMG10470 in vitro and in some Saudi foods viz., minced meat, pastenrized milk, kareesh cheese and tomatoes paste.
MATERIALS AND METHODS
Microorganisms and media: L. monocytogenes LMG10470 was kindly provided from Department of Food Technology and Food Preservation, Faculty of Agricultural and Applied Biological Sciences, University of Gent, Belgium. It was used as the indicator organism. It was maintained as frozen stocks in trypticase soy broth (Difco) plus 10% glycerol and was subcultured every 2 weeks in trypticase soy broth. Growth values of L. monocytogenes were measured on Listeria selective agar (Oxoid) as cfu/ml. growth values were taken from three replicates. Lactobacillus plantarum UG1 was used as a bacteriocin producer (plantericin UG1 producer) (Enan et al., 1994a, b). It was maintained at-20°C in De Man, Rogosa and Sharpe medium (MRS) (De Man et al., 1960) and was propagated in MRS broth (Enan et al., 1996; Enan, 2000).
Food samples: Samples of minced meat were prepared by their grinding in a sterile mincer. About 100 g portions were placed aseptically in sterile plastic bags and were used in the experiments. Tomato paste (Sadafco Co., Saudi Arabia) were prepared. About 100 g samples of tomato paste were placed aseptically in 250 mL sterile screw capped bottles and were used in the experiments. Pasteurized milk (Nadec Co., Saudi Arabia) was added aseptically in sterile 250 mL Erlenmeyer flasks (100 mL for each) and was used in the experiments. Kareesh cheese is an Egyptian famous cheese. It was provided from Wella Marketing Company (Saudi Arabia) and was cutted aseptically by sterile knife. Hundred gram portions of Kareesh cheese were placed aseptically in sterile plastic bags and were used in the experiments. This investigation was carried out in Laboratory of Microbiology, Department of Sciences, King Khalid Military Academy, Saudi National Gaured, Saudi Arabia.
Preparation of bacteriocins and bacteriocin bloassay: The bacteriocin nisin was provided from Somatco Chemical Company (Saudi Arabia). Stock nisin solution was prepared by solubilizing appropriate amounts of powder in 0.02 HCl solution. The pH was adjusted to 3.5 with 1N NaOH. The solution was then filter-sterilized (0.22 μm, Milipore). The nisin bioassay was determined by the critical dilution assay (Biswas et al., 1991). The Listeria monocytogenes LMG10470 was used as the indicator organism. One IU (International Unit) of nisin was showed to be equivalent to 0.20 AU (Arbitarary Unit). So, 1 mL of nisin solution containing 104 IU was showed to contain 2000 Arbitarary Units (AU).
The bacteriocin plantaricin UG1 was prepared according to the procedures described previously (Enan, 2002, 2005). Briefly, a 16 h old culture in MRS broth of Lactobacillus plantarum UG1 was centrifuged at 10000xg for 10 min at 4°C. Cell free supernatants were subjected to ammonium sulphate precipitation. The precipitates were recovered in 10mM potassium phosphate buffer pH 6.5 and were sterilized by filtration (0.22 μm Milipore). One milliliter of partially purified plantaricin UG1 was assayed against Listeria monocytogenes LMG10470 and was showed to contain 1920 Arbitarary Units (AU).
Inhibition of L. monocytogenes LMG10470 by bacteriocins in vitro and in food: The food samples mentioned above were inoculated with 105 cfu mL-1 or gm final concentration. The above inoculated food samples were treated with either 4000 AU mL-1 of nisin or 4000 AU mL-1 plantaricin UG1. These bacteriocins were used individually or in combination. The inoculated and treated samples were shaken vigorously by hand to distribute both inocula and bacteriocins and were then incubated at 8°C for 4 days. The survivors of listerias cells were enumerated onto Listeria selective agar at different time intervals. Each test was run in duplicate and mean values were calculated.The above procedure was based on the procedure proposed by Vignolo et al. (2000). The regrowing resistant variants as well as resistance frequency were studied by Enan (2005).
RESULTS
The effect of nisin and plantaricin UG1 individually or in combination on the growth of the L. monocytogenes LMG10470 cells in BHI broth and in pasteurized milk is shown in Fig. 1. The action of nisin and plantaricin UG1 was characterized by a two phase response: at first, viable listerias populations decreased by 2-3 log cycles within almost 36-48 h in either BHI broth or pasteurized milk. However, no growth was observed in all samples tested after almost 3 days. Thereafter, a resurgence of growth was observed. The mix of nisin and plantaricin UG1 prevented regrowth of the L. monocytogenes LMG10470 cells in both BHI broth and pasteurized milk.
Figure 2 shows the effect of bacteriocins on growth of the L. monocytogenes LMG10470 strain in tomato paste. Nisin-plantaricin UG1 mixture displayed the fastest rate of bactericidal action recording 100% cell destruction within 12 h, followed by both nisin and plantaricin UG1 which destroyed 100% of listerias cells within 48-60 h. By further storage of the treated food samples, no regrowth was observed in tomato paste samples treated with nisin-plantaricin UG1 mixture, but a slight regrowth (around 1x102) was observed after one week in tomato paste samples treated with either nisin or plantaricin UG1.
The addition of nisin; plantaricin UG1; nisin-plantaricin UG1 mixture to either minced meat or kareesh cheese samples (Fig. 3) resulted in a marked bactericidal effect with reduction in viable counts of L. monocytogenes LMG10740 by 3 log cycles; 3 log cycles; 5 log cycles after almost 48-60 h in minced meat, respectively and by 3 log cycles; 2 log cycles; 5 log cycles after almost 48-60 h in kareesh cheese samples (Fig. 3). After three to four days, no growth of listerias cells was observed in the treated samples of minced meat and Kareesh cheese; but thereafter a slight regrowth was observed only in samples treated with either nisin or plantaricin UG1. However, no regrowth was observed in samples of both minced meat and Kareesh cheese treated with nisin-plantaricin UG1 mixture (Fig. 3).
A representatives of the survived cells of L. monocytogenes LMG10470 were isolated from all food samples treated with either nisin or plantaricin UG1 and were proved to be resistant variants as they were still resistant to the respective bacteriocins onto BHI agar plates after 7 successive transfers. Mutants of listerias cells resistant to either nisin or plantaricin UG1 showed a resistance frequency in the range of 10-5 to 10-6. When resistance frequency in the range of 10-5 to 10-6.
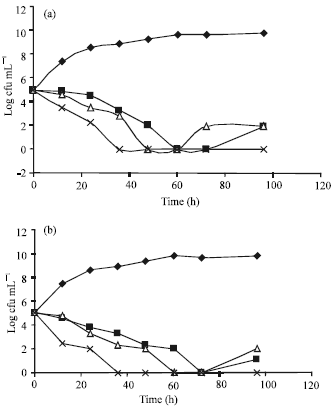 | |
| Fig. 1: | Growth of Listeria monocytogenes LMG 10470 in (a) brain heart infusion broth and in (b) pasteurized milk held at 8°C with or without bacteriocins. |
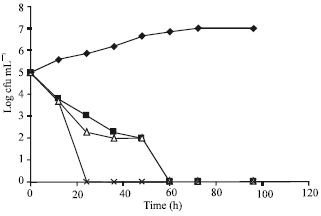 | |
| Fig. 2: | Growth of Listeria monocytogenes LMG 10470 in tomato paste held at 8°C with or without bacteriocins. |
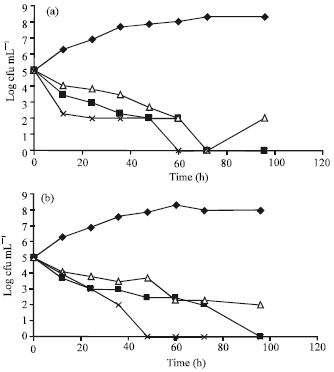 | |
| Fig. 3: | Growth of Listeria monocytogenes LMG 10470 in (a) minced meat and in (b) kareesh cheese held at 8°C with or without bacteriocins. |
When the sensitivity of the mutant listerias cells to the respective bacteriocins in media containing either nisin or plantericin UG1 was tested, the variants didn't show cross-resistance. The survivors isolated from samples containing nisin were resistant only to nisin but were sensitive to plantericin UG1. Also, the plantaricin UG1mutants were sensitive to nisin and and where insensitive to this bacteriocin.
DISCUSSION
Bacteriocins of lactic acid bacteria, natural antimicrobial peptides, have been proposed by many researchers for controlling L. monocytogenes in food products (Muriana, 1996). Nisin produced by some Lactococcus lactis subsp. Lactis strains, belongs to calss I bacteriocins (lantibiotic peptides) (Klaenhammer, 1993). This bacteriocin has been approved by the World Health Organization as a preservatique in the food industry (Ronk, 1988). Unfortunately, a spontaneous resistant variants of L. monocytogenes to nisin were detected and characterized Rekhif et al. (1994), Mazzotta and Montville (1997) and Vignolo et al. (2000). This clearly suggests that there is a need to continue research to explain the nature of resistance and to find out other bacteriocins having different modes of resistance to be mixed with nisin.
The effect of either nisin or plantaricin UG1 noticed in this study induced a bactericidal action on L. monocytogenes followed by regrowth of the target cells. This transitory inhibitory effect has been reported previously for nisin (Bouttefroy and Milliere, 2000) and plantaricin UG1 (Enan, 2002). Representatives of the regrowing cells were isolated and were proved to be resistant cells as they were still resistant to either nisin or plantaricin UG1 in BHI agar containing the same bacteriocins. The resistant variants of L. monocytogenes resistant to either nisin (Vignolo et al., 2000) or plantaricin UG1 (Enan, 2005) had been isolated and characterized. The results obtained have showed that the resistance frequency in L. monocytogenes to either nisin or plantaricin UG1 was showed to be 10-5 to 10-6. Many authors also (Davis and Adams, 1994; Mazzota and Montville, 1997) showed that nsin resistant varients of L. monocytogenes appear at frequencies of 10-5 to 10-8. Also, Enan (2005) showed a plantericin UG1 resistance in L. monocytogenes at a resistance frequency around 10-6. Nisin resistance response of L. monocytogenes was reported to be correlated with membrane fatty acid composition, a significant reduction in membranes phospholipids and, therefore, inability of nisin to form pores in more rigid membranes (Abee, 1995). In contrast the resistance phenotype of listerias cells to plantaricin UG1 was due to modification in adsorption site (s) of the target cells (Enan et al., 1996). A combination of two bacteriiocins differying in their modes of resistance have a higher protective ability for foods against L. monocytogenes strains than when used individually (Hanlin et al., 1993).
The results presented here indicate that nisin-plantaricin UG1 combinations avoided the regrowth of bacteriocin resistant cells of L. monocytogenes in vitro and in foods (pasteurized milk, tomato paste, minced meat, kareesh cheese). Such results supported many published results in this respect (Hanlin et al., 1993; Bouttefroy and Milliere, 2000; Vignolo et al., 2000). A mixture of two bacteriocins would be more bactericidal than one bacteriocin, since cells resistant to one bacteriocin would be killed by another. In addition mainly synergistic effects were reported when the interactions between pair of bacteriocins were used (Mulet-Powell et al., 1998). Moreever, a mixture containing two bacteriocins that belong to different classes would be more effective in food protection than one only. This is the case in this study, nisin is a lantibiotic (Moll et al., 1999) (Class I). However, plantaricin UG1 is probably class IIb bacteriocins (Enan et al., 2004). No resistant variants of L. monocytogenes were isolated from foods samples treated with nisin-plantaricin UG1mixture. This is a promising result to use nisin-plantaricin UG1 mixture as biopreservative for dairy products, tomato paste and minced products, tomato paste and minced meat in Saudi Arabia. No cross resistance was found between nisin and plantaricin UG1 whose synergistic listericidal effect prevented the emergence of spontaneous variants of L. monocytogenes in pasteurized milk, tomato paste kareesh cheese and minced meet.
Based on the results employed herein and in agreement with Vignolo et al. (2000), the industrial application of combined partically purified and/or purified bacteriocins in food products (dairy products, tomato paste and minced meat could be a useful approach to reduce the frequency at which resistant populations develop, to improve the hygiene standards and to extend the shelf life of dairy products, meat products and tomato paste. Although the use of nisin is permitted in more than 50 countries in a variety of foods, other bacteriocins with different and/or more effective antibacterial activity may be considered and approved to mix with nisin to be of more hazardous quality.
ACKNOWLEDGEMENT
G. Enan is indepted to Prof. Dr. J. Debevere: the Chairman of Department of Food Technology and Food Preservation at University of Gent, Belgium, for providing the experimental strains.
REFERENCES
- Abee, T., 1995. Pore forming bacteriocins of Gram-positive bacteria and self protection mechanism of producer organisms. FEMS Microbiol. Lett., 129: 1-9.
Direct Link - Biswas, S.R., P. Ray, M.C. Johnson and B. Ray, 1991. Influence of growth conditions on the production of a bacteriocin, pediocin AcH, by Pediococcus acidilactici H. Applied Environ. Microbiol., 57: 1265-1267.
PubMedDirect Link - Bouttefroy, A. and J.B. Milliere, 2000. Nisin-curvaticin 13 combinations for avoiding the regrowth of bacteriocin resistant cells of L. monocytogenes ATCC 15313. Int. J. Food Microbiol., 62: 65-75.
Direct Link - De Man, J.C., M. Rogosa and M.E. Sharpe, 1960. A medium for the cultivation of Lactobacilli. J. Applied Bacteriol., 23: 130-135.
CrossRefDirect Link - Enan, G., A.A. El-Essawy, M. Uyttendaele and J. Debevere, 1996. Antibacterial activity of Lactobacillus plantarum UG1: Production, characterization and bactericidal action of plantaricin UG1. Int. J. Food Microbiol., 30: 189-215.
Direct Link - Enan, G., 2000. Inhibition of Bacillus cereus ATCC 14579 by plantaricin UG1 in vitro and in food. Nahrung, 44: 364-367.
PubMed - Enan, G., S. Alayan, H.A. Abdel-Salam and J. Debevere, 2002. Inhibition of Listeria monocytogenes LMG 10470 by plantaricin UG1 in vitro and in beef meat. Nahrung, 46: 411-414.
Direct Link - Hanlin, M.B., N. Kalchayanand, P. Ray and B. Ray, 1993. Bacteriocins of lactic acid bacteria in combination have greater antibacterial activity. J. Food Prot., 56: 252-255.
Direct Link - Klaenhammer, T.R., 1993. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev., 12: 39-85.
CrossRefPubMedDirect Link - Moura, S.M., M.T. Destro and B.D. Franco, 1993. Incidence of Listeria species in raw and pasteurized milk produced in SaoPaulo, Brazil. Int. J. Food Microbiol., 19: 229-237.
PubMedDirect Link - Mulet-Powell, J., A.M. Lacoste-Armynote, M. Vinas and M. Simoen de Buochberg, 1998. Interaction between pairs of bacteriocins from lactic acid bacteria. J. Food Prot., 61: 1210-1212.
Direct Link - Pucci, M.J., E.R. Vedamuthu, B.S. Kunka and P.A. Vandenbergh, 1988. Inhibition of Listeria monocytogenes by using bacteriocin PA-1 produced by Pediococcus acidilactici PAC 1.0. Appl. Environ. Microbiol., 54: 2349-2353.
CrossRefDirect Link - Rekhif, N., A. Atrih and G. Lefevure, 1994. Selection and properties of spontaneous variants of L. monocytogenes ATCC 15313 resistant to different bacteriocins produced by lactic acid bacteria strains. Curr. Microbiol., 28: 237-241.
CrossRefDirect Link - Vignolo, G., J. Palacios, M.E. Farias, F. Sesma, U. Schillinger, W. Holzapfel and G. Oliver, 2000. Combined effect of bacteriocins on the survival of various Listeria species in broth and meat system. Curr. Microbiol., 41: 410-416.
CrossRefDirect Link