
Mahmoud M. El-Attrouny
Department of Animal Productions, Faculty of Agriculture, Benha University, Moshtohor, Toukh 13736, Qalyubia, Egypt
LiveDNA: 20.33208
Mahmoud M. Iraqi
Department of Animal Productions, Faculty of Agriculture, Benha University, Moshtohor, Toukh 13736, Qalyubia, Egypt
Asian Journal of Poultry Science
Year: 2020 | Volume: 14 | Issue: 2 | Page No.: 24-31







ABSTRACT
Background and Objective: Poultry industry aims mainly to produce genetically superior animals with high productivity of egg and meat. On a commercial scale, the purpose of most quail genetic improvement programs is to select genetically superior birds for meat and egg production. The objective of this study was to investigate the effect of short-term selection through four generations of selection in Japanese quail on egg number at the first 90 days of laying, estimate genetic parameters of egg production traits and estimate the correlated response to selection. Materials and Methods: Data of 1,352 female Japanese quails were collected through five consecutive generations (base and four selected generations) to estimate genetic gain, depending on breeding values, for Age at First Egg (AFE), Body Weight at Sexual Maturity (BWSM), Weight at First Egg (WFE), Egg Number at first 90 days (EN90D) and Average Egg Weight (AEW). Results: Estimates of heritability for egg production traits were ranged from 0.17 to 0.26. Moreover, positive genetic correlations were detected (0.18 to 0.43) between all studied egg production traits, except the correlation between AFE and EN90D (-0.32). The contrast among estimates was significant (p<0.05), favoring the selected generations over the base. The cumulative selection response was favorable for all unselected traits with the superiority of the selected generations to the base generation. Conclusion: It can be concluded that early selection based on breeding value for a cumulative egg number at first 90 days of egg production can effectively improve the total egg production and other egg traits in Japanese quail through four selected generations.
PDF Abstract XML References Citation
How to cite this article
Mahmoud M. El-Attrouny and Mahmoud M. Iraqi, 2020. Correlated Responses to Short-term Selection for Egg Production Traits in Japanese Quail. Asian Journal of Poultry Science, 14: 24-31.
DOI: 10.3923/ajpsaj.2020.24.31
URL: https://scialert.net/abstract/?doi=ajpsaj.2020.24.31
DOI: 10.3923/ajpsaj.2020.24.31
URL: https://scialert.net/abstract/?doi=ajpsaj.2020.24.31
INTRODUCTION
Worldwide, there is an expanding market for quail products, including quail eggs. Quail egg production is concentrated in East Asia, Latin America and some European countries, with more advances in the production technology of pickled egg beside fresh eggs1,2. For that, the chain industry of quail egg becomes an important segment as an animal protein source with high biological value for human consumption. Japanese quail has been considered as an economic animal model for research due to its short generation interval and incubation periods, early maturation, suitability for genetic studies, resistance against diseases and great egg production persistency on a high level3-6. Egg production is one of the important economic traits of Japanese quail, which is influenced by genetic and environmental factors. Therefore, more research particularly in the genetic improvement for egg production is required to support this important egg chain2.
Selection programs are the most important tool for achieving genetic gain in quail7,8. Previous studies showed that performing selection programs increased egg production of Japanese quail9-12. The genetic improvement programs by selection depend on genetic variability for productive traits13-15. For genetic evaluation, it is necessary to estimate heritability and genetic correlations between egg production traits.
Egg production performance is usually evaluated in poultry by other traits, which can be considered as selection criteria, such as age at first egg, body weight at sexual maturity, egg number, egg weight and rate of laying during certain production periods7,16. Selection based on partial egg production periods is a usual approach for improving egg production in poultry to achieve major genetic gain across generations17-19. The selection for partial periods shortens the generation interval, hence greater genetic gain could be obtained in egg production12,20,21.
The peak of egg production in quail occurred at part-period (3 months) egg number of laying with the egg production rate ranged from22-25 85 to 95%. It was illustrated that genetic improvement for increasing egg numbers would increase the production rate and profitability of Japanese quail12,26.
Therefore, this study using a selection experiment was conducted to, (i) estimate genetic parameters of egg production traits during the first 90 days of production, (ii) estimate response to selection for increase egg production and correlated responses of egg traits and (iii) investigate the breeding value estimates and the genetic trends for the egg traits across the four selected generations.
MATERIALS AND METHODS
Population structure and bird management: This study was conducted from September, 2018 to December, 2019 at the Poultry Research Farm, Faculty of Agriculture, Benha University, Egypt. Research on live animals met the guidelines approved by the Institutional Animal Care and Use Committee in Egypt. A selection experiment was carried out for five generations and used a total number of 1352 females (262 base population and 1090 for the selected generations). In the first generation, 133 males and 266 females birds were selected according to the breeding value of egg number at the first 90 days of laying from the base population (Table 1). The selected birds were housed (one selected male with two selected females) in each breeding cage with dimensions of 25×25×25 cm and with a sloping floor for egg collection. All birds were fed a standard diet containing 20% crude protein and 12.56 MJ kg–1 according to the nutrient requirements recommendations for Japanese quail27. The temperature of the hen house was around 22°C and the light was given 16 h per day and 8 h of darkness. Eggs were collected daily during the first 90 days of laying, in a pedigree system for each family depending on its sire and dam. Eggs from each dam/cage were identified by a specific pattern of egg surface. The structures of the collected data during the experiment were presented in (Table 1).
Hatched chicks from each sire and dam, were wing-banded by small size plastic bands, which were replaced by wing metal bands at 14 days of age. The chicks were housed in brooder cages from hatching to 14 days of age and then transferred into rearing cages. Bird comfort and housing conditions were carefully monitored daily. A feed consisting of 24% crude protein and 12.14 MJ of ME kg–1 based on the nutrient requirements recommendations for Japanese quail27 during the growth period (first 6 weeks). Feed and water were supplied ad libitum and illumination was available 24 h.
| Table 1: | Numbers of sires, dams and hens for different generations |
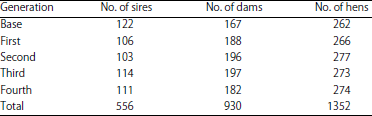 | |
After 6 weeks of age, all females were transferred to an individual laying cage where egg production was recorded. All birds were kept under the same managerial, hygienic and environmental conditions. Age at First Egg (AFE), Body Weight at Sexual Maturity (BWSM), Weight at First Egg (WFE), Days Needed to produce the first ten eggs (DN10), egg number at first 90 days of laying were recorded for each bird (egg number were collected daily during first 90 days) and Average Egg Weight (AEW) and all eggs laid by each bird were weighed individually 4 days/week.
Genetic analysis: Data of egg production traits were analyzed using the following multi-trait animal model:
y = Xb + Zaua + e
where, y is the vector of observations hen,; b is the vector of fixed effects of generation (five levels), X and Za is incidence matrices corresponding to fixed and additive random effects of the hen (ua), respectively, e is vector of random residual effects. The VCE6 software was used to estimate the variance components of random effects and heritabilities28. These estimates were used to solve the corresponding mixed model equations, obtaining solutions for the generation effects and their error variance-covariance matrix using the PEST software28.
The genetic (rg) and phenotypic (rp) correlations between egg production traits were also estimated according to the formula29,30:
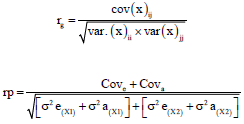 |
where, Cov (X)ij is The covariance between additive genetic effects for egg production traits, Xii and Xjj is the additive genetic (a) variances of ith and jth egg production traits. Cove is covariance of error between egg production traits,; Cova is covariance between egg production traits for the hen, σ2e(X1) is the variance of error for trait 1, σ2a(X1) is the additive variance of hen for trait 1, σ2e(X2) is the variance of error for trait 2, σ2a(X2) is the additive variance of hen for trait 2.
Estimates of the contrasts between the five generations of selection were obtained by the least-squares analysis using the VCE6 software. The Predicted Breeding Values (PBV) of the birds with and without records were estimated using the PEST program28 to evaluate genetic gain. The averages of the predicted breeding values in each generation were regressed on the generation number to estimate the genetic trend. The cumulated selection response (CSR) at generation was calculated by the following equation31:
CSR = iXhxhyrgσpy
where, CSRy is the correlated response in trait y, i is the selection intensity for trait x, hx and hy is square roots of heritability for traits x and y, respectively, rg is the genetic correlation of traits x and y and σpy is the phenotypic standard deviation of trait y.
RESULTS AND DISCUSSION
The statistical descriptions including mean, coefficient of variation, minimum and maximum were presented in Table 2. In the current study, the average for egg production traits was 49.9 and 13.2 days for AFE and DN10. For BWSM, WFE and AEW, the average recorded was 258.4, 11.5 and 13.8 g, respectively and the EN90D was 63.2 eggs in Japanese quail. These findings are in agreement with those for AFE26,32, BWSM16,33-35, WFE12,13 and EN90D36-38. However, values for AFE, BWSM and EN traits are in contrast with previous studies14,24,39. Coefficients of variation (CV%) for all studied traits were moderate, ranging from 7.02 to 11.0%, which are in agreement with previous studies40,41,4,26.
Genetic parameters including heritability, genetic and phenotypic correlations for egg production traits are presented in Table 3. Estimates of heritability were low and moderate for AFE (0.20), BWSM (0.24) and DN10 (0.17). These estimates are related to the decrease in the genetic variability of the studied population, as a result of the selection program, which could be due to the population structure. Similar heritability values were reported for AFE ranging from 0.21 to 0.2724,35,36. In addition, Abou Khadiga et al.32 reported a similar heritability for BWSM (0.25).
| Table 2: | Descriptive statistics for studies traits in Japanese quail |
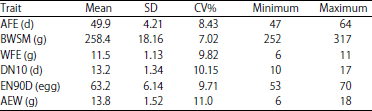 | |
AFE: Age at first egg, BWSM: Weight at sexual maturity, WFE:, Weight of first egg, DN10: Days needed to produce the first ten eggs, EN90D: Egg Number at first 90 days of laying and AEW: Average egg weight | |
| Table 3: | Estimates of heritabilities (diagonal), genetic (above the diagonal) and phenotypic (below the diagonal) correlations with their standard errors (±) of traits |
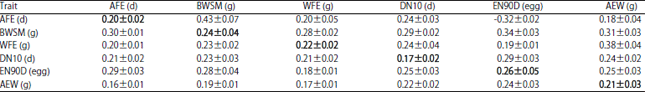 | |
AFE, age at first egg; BWSM, weight at sexual maturity; WFE, Weight of first egg; DN10, days needed to produce the first ten eggs; EN90D, Egg Number at first 90 days of laying and AEW, average egg weight | |
In contrast, Özsoy and Aktan4, reported higher estimates of heritability for BWSM (0.58) than those reported in the current study. Heritability for WFE (0.22) was higher than the estimates reported by Farrag et al.42, Daikwo et al.24, Mahmoud et al.11 ranging from 0.04 to 0.16.
Estimates of heritability were moderate for EN90D (0.26), in the present study, which indicated an increase in the additive genetic variance over generations. Valente et al.36 reported low heritability of EN (0.15) from 11 to 16 week of age. Narinc et al.43 reported high heritability estimates (0.43) for monthly egg production in laying quail. Okenyi et al.37 revealed that heritabilities estimates varied from low to high (0.12, 0.33 and 0.48) in three selected generations for egg production in Japanese quail. Moreover, Stivanin et al.12 detected moderate heritability estimates, for partial production of eggs, for the yellow (0.26 to 0.36) and Black (0.25 to 0.35) lineages of quail.
The current study showed that the heritability for AEW was moderate (0.21) as presented in Table 3. This agrees with previous studies, who showed similar moderate heritability estimates for AEW in Japanese quail. However, some studies showed a varied range of heritability estimates of AEW ranging from 0.14 to 0.83 in quail13,15,26,41.
Estimates of the genetic correlations between studied traits showed that AFE was positively correlated with all studied traits with low to moderate rg ranged from 0.18 to 0.43 (Table 3) and was negatively correlated with EN90D (-0.31). It means that selection for early AFE will improve egg production. Lotfi et al.26 found high negative rg estimated were found between AFE and EN from 9 to 12 week of age (-0.74 to -0.89) in Japanese quail. Also, in Japanese quail, negative and high magnitude genetic correlation between AFE and EN was reported by Hidalgo et al.15. Abou Khadiga et al.32 revealed high positive rg estimates between AFE and both BWSM and DN10 (0.53 and 0.45, respectively) in Japanese quail. Özsoy and Aktan4 reported that rg estimates between AFE and EW had a low magnitude of 0.18.
Phenotypically, in the current study, AFE positively correlated with all egg production traits ranged from low to moderate (0.16-0.30) as presented in Table 3. This agrees with the previous studies of Karabag et al.16 and Alkan et al.44, who reported a moderate positive rp between AFE and FEW being 0.320 and 0.224, respectively. Abou Khadiga et al.32 reported low positive rp estimates between AFE and each BWSM and DN10 of 0.18 and 0.08, respectively. Özsoy and Aktan4 reported low negative rp estimates between AFE and EW of -0.10. In addition, Farrag42 reported high negative rp estimates between AFE and EN of -0.697 in the selected line of Japanese quail. Daikwo et al.24 reported that estimates of rp between AFE and each WFE, EN and EW were 0.82, 0.71 and -0.85, respectively.
From the current findings, there were positive and high genetic correlations (ranged 0.28-0.34) estimates between BWSM and all egg production traits. This means that hens with heavier body weight at sexual maturity produce higher eggs number. Lotfi et al.26 found a positive rg between BWSM and AEW from 9 to 12 week of age (0.65 to 0.94). Özsoy and Aktan4 showed a low rg (0.12) between BWSM and EW.
Phenotypic correlation estimates between BWSM and all studied traits were positive and ranged from low to moderate (0.19-0.30). Özsoy and Aktan4 reported a high and positive phenotypic correlation estimate between BWSM and EW (0.51). In the same trend, there was a positive phenotypic correlation estimate between BWSM and WFE (0.48)11. Lotfi et al.26 reported low and negative rp between WSM and EN from 9 to 12 weeks of age (-0.02).
The estimate of the genetic correlation was 0.25 between EN90D and AEW and was 0.38 between WFE and AEW. Lotfi et al.26 found a genetic correlation between AEW and EN at different weeks of age ranged from negative to positive values (-0.28 to 0.45). Hidalgo et al.15 reported positive rg estimates between EN and AEW being 0.58 for yellow line, 0.09 for Blue line and 0.09 for red line whereas there was negative rg between the previous traits of -0.18 for meat line in Japanese quail. Stivanin et al.12 found that rg between total egg production for 150 days and average egg weight presented values ranging from 0.45 to 0.49 in both yellow and black lineages of quail.
The current findings revealed that the phenotypic correlation estimate between EN90D and AEW was moderate (0.24). This in contrast with the findings of Daikwo et al.24 who reported that the phenotypic correlation between EN and EW at 90 days of laying was negative and low (-0.10) in Japanese quail.
| Table 4: | Least-square means for egg production traits in different generations along with all possible generation contrasts and their standard errors (±SE) in Japanese quails |
 | |
AFE: Age at first egg, BWSM: Weight at sexual maturity, WFE: Weight of first egg, DN10: Days needed to produce the first ten eggs, EN90D: Egg Number at first 90 days of laying and AEW: Average egg weight, a-eMeans with the same letters within each trait are not significantly different (P×0.05)* Generation effect significantly different (P 0.05), -,+Sign is the mathematical result of the contrast operation which determine the superiority of generations and describe the direction of improvement | |
Similarly, Lotfi et al.26 found that the rp correlation between AEW and EN from 9 to 12 week of age was 0.14 in Japanese quail.
Least squares means of the studied traits for both base and selected generations are presented in Table 4. The mean for AFE in the base, 1st, 2nd, 3rd and 4th generations were 52.1, 50.7, 49.6, 48.7 and 48.5 days, respectively. The corresponding generational means for DN10 were 14.21, 13.33, 13.12, 12.42 and 12.35 days, respectively. The differences were significant (p<0.05) for AFE and DN10 between generations. Age at first egg and DN10 decreased progressively with advancing generations. The decreasing trend observed for AFE and DN10 over generation reflects a negative correlated response in these traits due to the selection for increased egg numbers. Our findings agree with those reported for AFE4,32,36,45. However, a different trend reported by Bahie El-Deen et al.46 who showed that the overall means of AFE were 52.61 and 50.45 d for control and selected lines in Japanese quail.
Least square means for BWSM were 244, 251, 259, 264 and 272 g of the base, 1st, 2nd, 3rd and 4th generations, respectively (Table 4). There were significant differences (p<0.05) for BWSM among generations with an increasing trend over generations. This result indicated a positive correlated response for BWSM when performing selection for high EN90D, which agreed with the findings of Karabağ et al.16, Özsoy and Aktan4 and Abou Khadiga et al.32.
The means for egg number and average egg weight are shown in Table 4. The least square means of EN90D in the base, 1st, 2nd, 3rd and 4th, generations were 58.9, 60.7, 63.5, 65.8 and 66.6 eggs, respectively. Means for AEW were 13.0, 13.5, 13.7, 14.2 and 14.3 g, respectively.
| Table 5: | Estimates of Cumulative Selection Response (CSR) and their (±standard errors) for the unselected traits through four generations of selection |
 | |
AFE: Age at first egg, BWSM: Weight at sexual maturity, WFE: Weight of first egg, DN10: Days needed to produce the first ten eggs, Average egg weight, *p<0.05 | |
There were significant differences (p<0.05) in EN90D and AEW between the generations. While EN90D and AEW increased progressively with generations. These higher values reflect a positive correlated response in AEW due to selection for improved EN, which agreed with the positive genetic correlation between EN and AEW. Asasi and Jaafar47 showed that egg weight ranged from 9.76 to 11.63 g in Japanese quail. However, Khaldari et al.14 reported that egg weight was 13.40 and 12.80 g in the selected and control populations, respectively, during selection for increased body weight at 4 week.
Estimates of contrast between selected and control generations for the egg production traits are given in Table 4. The contrast among estimates of these generations was significant (p<0.05) for AFE, BWSM, WFE, DN10, AEW, favoring the selected generations. Similar results were reported by Alkan et al.44 and Mahmoud et al.11.
Genetic gain to selection: Cumulative selection response after four selected generations was favorable for all unselected traits and showed the superiority of the selected generations compared to the base generation (Table 5).
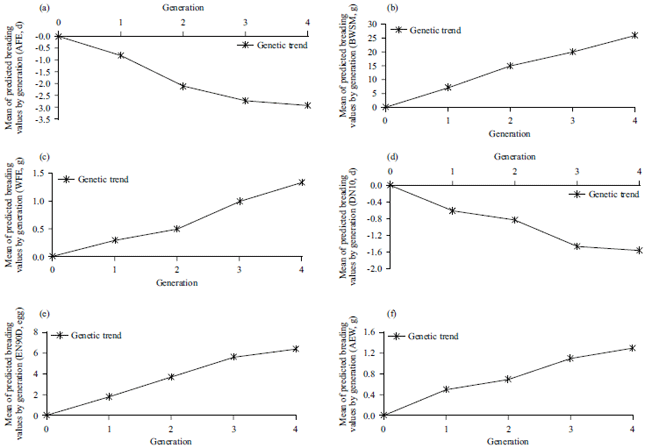 | |
| Fig. 1(a-f): | Means of the predicted breeding values by generation for unselected traits in Japanese quail |
Responses were significant (p<0.05) for AFE (-3.03 days), BWSM (26.8 g), WFE (1.4 g) and DN10, (-1.6 days) while was not significant for AEW (0.98 g) after four selected generations. These results reflect the positive responses to selection, for unselected traits for BWSM, WFE and AEW and negatively correlated responses for AFE and DN10. This demonstrated that, selection for egg number was associated with an increase in BWSM, WFE and AEW. On the other hand, the decline in AFE and DN10 indicates that selection for EN was associated with a decrease in AFE and DN10. Clearly, the realized genetic gains from this selection program depending on breeding value estimates could result in a great improvement in genetic gain. Similar results were reported for AFE, BWSM and DN1032,45, the same results have been shown for EN and AEW12. On the other hand, Okenyi et al.37 reported lower BWSM and EW in a selected line for short-term 30-day egg production for two generations. Ramzy et al.48 reported that cumulative selection response to selection for early sexual maturity decreased EW from 10.74 to 10.04 g but BWSM increased from 136.32 to 139.32 g after four selected generations in Japanese quail.
The genetic trends for egg production traits across the generations were presented in Fig. 1. Estimates of the genetic trend for BWSM, WFE, EN90D and AEW gradually increased as the generation advanced as shown in Fig. 1b, c, e and f. On the other hand, genetic trend estimates for AFE and DN10 gradually decreased in the subsequent selection generations as shown in Fig. 1a and d. This increased genetic trend in the selected generations illustrated the efficiency of selection for EN90D of laying. The estimation of the genetic trend was necessary for the evaluation of the genetic improvement program. The current results agree with Abou Khadiga et al.32 who observed a linear decrease trend of AFE and DN10 in selected populations as the generations progressed.
CONCLUSION
In conclusion, the construction of genetic improvement program through selection depending on the breeding value for EN90D of laying could result in a great improvement in genetic gain. Superiorities in the selected generations indicated that selection for EN90D in Japanese quail was associated with an improvement in other egg production traits. In addition, the cumulative correlated responses of selection for high EN90D of laying in Japanese quail through four selection generation increased with generations. Therefore, genetic improvement in egg production performance in Japanese quail could be achieved through practicing selection for egg number at first 90 days of laying and more improvement could be achieved through selection for more generations based on breeding value estimates. This saves effort, cost and increases the return and profitability of the Japanese quail’s breeder.
SIGNIFICANCE STATEMENT
Practicing short-term selection for egg number at first 90 days of laying based on breeding value estimate successfully improved egg production performance, saved time, effort, cost and increased egg profitability in Japanese quail.
ACKNOWLEDGMENT
The authors thank the personnel working at the Poultry Research Farm at Faculty of Agriculture, Benha University, Egypt.
REFERENCES
- Minvielle, F., 2004. The future of Japanese quail for research and production. Poult. Sci., 60: 500-507.
CrossRefDirect Link - Alkan, S., K. Karabag, A. Galic, T. Karslı and M.S. Balcıoglu, 2010. Determination of body weight and some carcass traits in japanese quails (Coturnix coturnix japonica) of different lines. Kafkas Univ. Vet. Fak. Derg., 16: 277-280.
CrossRefDirect Link - Narinc, D., E. Karaman, T. Aksoy and M.Z. Firat, 2014. Genetic parameter estimates of growth curve and reproduction traits in Japanese quail. Poultry Sci., 93: 24-30.
CrossRefDirect Link - Molino, A.B., E.A. Garcia, G.C. Santos, J.A.V. Filho, G.A.A. Baldo and I.C.L. Almeida Paz, 2015. Photostimulation of Japanese quail. Poult Sci., 94: 156-161.
CrossRefDirect Link - Durmus, I., S. Alkan, D. Narinc, K. Karabag and T. Karsli, 2017. Effects of mass selection on egg production on some reproductive traits in Japanese quail. Europ. Poult. Sci.,.
CrossRefDirect Link - Ahmad, S., S. Mehmood, K. Javed, A. Mahmud and M. Usman et al., 2018. Different selection strategies for the improvement of the growth performance and carcass traits of Japanese quails. Braz. J. Poult. Sci., 20: 497-506.
CrossRefDirect Link - Nestor, K.e., W.L. Bacon and A.L. Lambio, 1983. Divergent selection for egg production in Coturnix coturnix japonica. Poul. Sci., 62: 1548-1552.
CrossRefDirect Link - Maeda, Y., F. Minvielle, S. Okamoto and T. Hashiguchi, 1999. Changes of protein polymorphism in selection program for egg production in Japanese quail, Coturnix coturnix japonica. J. Poult. Sci., 36: 83-95.
CrossRefDirect Link - Mahmoud, B.Y.F., G.A. Khadiga and E.A. El-Full, 2015. Correlated response to selection for some egg performance traits in egg line of Japanese quail. Anim. Prod. Sci., 56: 913-917.
CrossRefDirect Link - Stivanin, T.E., F.C. Maia, E. Migliorini, S. Kluska, S.T. Amorim, F.S Lovatto, and E.N. Martins,, 2019. Evaluation of selection criteria in laying quail (Coturnix coturnix japonica). Livestock Res. Rural Dev.
Direct Link - Saatci, M., H. Omed and I.A. Dewi, 2006. Genetic parameters from univariate and bivariate analyses of egg and weight traits in Japanese quail. Poult. Sci., 85: 185-190.
CrossRefDirect Link - Khaldari, M., A. Pakdel, H.M. Yegane, A.N. Javaremi and P. Berg, 2010. Response to selection and genetic parameters of body and carcass weights in Japanese quail selected for 4-week body weight. Poult. Sci., 89: 1834-1841.
CrossRefDirect Link - Hidalgo, A.M., E.N. Martins, A.L. Dos Santos, T.C.O. De Quadros, A.P.S. Ton and R. Teixeira, 2011. Genetic characterization of egg weight, egg production and age at first egg in quails. R. Bras. Zootec., 40: 95-99.
CrossRefDirect Link - Karabag, K., 2010. The differences in some production and clutch traits in divergently selected Japanese Quails. J. Fac. Vet. Med. Univ. Kafkas, 16: 383-387.
Direct Link - Poggenpoel, D.G., G.F. Ferreira, J.P. Hayes and J.J. Du Preez, 1996. Response to long-term selection for egg production in laying hens. Br. Poul. Sci., 37: 743-756.
CrossRefDirect Link - Luo, P.T., R.Q. Yang and N. Yang, 2007. Estimation of genetic parameters for cumulative egg numbers in a broiler dam line by using a random regression model. Poult. Sci., 86: 30-36.
CrossRefDirect Link - Cruz, V.A.R., A.V. Pires, F.M. Gonçalves, R.A. Torres Filho, I.G. Pereira and C.V. Araújo, 2016. Egg production selection in partial and accumulated periods in a commercial broiler line. Arq. Bras. Med. Vet. Zootec., 68: 489-496.
CrossRefDirect Link - Karami, K., S. Zerehdaran, M. Tahmoorespur, B. Barzanooni and E. Lotfi, 2016. Genetic evaluation of weekly body weight in Japanese quail using random regression models. Br. Poultry Sci., 58: 13-18.
CrossRefDirect Link - Khadiga, G.A., B.Y.F. Mahmoud, G.S. Farahat, A.M. Emam and E.A. El-Full, 2017. Genetic analysis of partial egg production records in Japanese quail using random regression models. Poultry Sci., 96: 2569-2575.
CrossRefDirect Link - Nestor, K.E. and W.L. Bacon, 1982. Divergent selection for body weight and yolk precursor in Coturnix coturnix japanese. 3. Correlated responses in mortality, reproduction traits and adult body weight. Poult. Sci., 61: 2137-2142.
CrossRefDirect Link - Minvielle, F., G.Gandemer, Y. Maeda, G.Leborgoyen and M.Boulay, 2000. Carcass characteristics of a heavy Japanese quail line under introgressios with the roux gene. Br. Poul. Sci., 41: 41-45.
CrossRef - Daikwo, S.I., N.I. Dim and O.M. Momoh, 2014. Genetic parameters of some egg production traits in japanese quail in a tropical environment. J. agric. vet. sci., 7: 39-42.
CrossRefDirect Link - Lukanov, H., A. Genchev and P. Kolev, 2019. Egg quality traits in WG, GG and GL Japanese quail populations. Trakia J. Sci., 17: 49-55.
CrossRefDirect Link - Lotfi, E., S. Zerehdaran and M.A.M. Azari, 2012. Estimation of genetic parameters for egg production traits in Japanese quail (Coturnix cot. japonica). Arch. Gefl, 76: 131-135.
Direct Link - Beckar, W.A., 1984. Manual of Quantitative Genetics. 4th Edn., Academic Enterpises, Pullman Washington, U.S.A, Pages: 190.
Direct Link - Khadiga, G.A., B.Y.F. Mahmoud and E.A. El-Full, 2016. Genetic evaluation of early egg production and maturation traits using two different approaches in Japanese quail. Poultry Sci., 95: 774-779.
CrossRefDirect Link - Sakunthaladevi, K.S., B.R. Gupta, M.G. Prakash, S. Qudratullah and A.R. Reddy, 2010. Genetic studies on growth and production traits in two strains of Japanese quails. Anim. Sci., 6: 223-230.
Direct Link - Mahmoud, B.Y., G. Abou Khadiga and E. A. El-Full, 2014. Direct and correlated responses to short-term index selection for some economic traits of Japanese quail. Egypt Poult. Sci., 34: 761-778.
CrossRefDirect Link - Valente, B.D., G.J.M. Rosa, M.A. Silva, R.B. Teixeira, and R.A. Torres, 2011. Searching for phenotypic causal networks involving complex traits: an application to European quail. Genet. Sel. Evol., 43: 37-48.
CrossRefDirect Link - Okenyi, N., H.M. Ndofor-Foleng, C.C. Ogbu and C.I. Agu, 2013. Genetic parameters and consequences of selection for short-term egg production traits in Japanese quail in a tropical environment. Afr. J. Biotechnol., 12: 1357-1362.
CrossRefDirect Link - Manaa, E.A., K. El-Bayomi and G. Sosa, 2019. Genetic evaluation for egg production traits in Japanese quail. Benha Vet. Med. J., 28: 178-185.
CrossRefDirect Link - Reddish, J.M., K.E. Nestor and M.S. Lilburn, 2003. Effect of selection for growth on onset of sexual maturity in randombred and growth-selected lines of Japanese quail. Poult. Sci., 82: 187-191.
CrossRefDirect Link - Sezer, M., E. Berberoglu and Z. Ulutas, 2006. Genetic association between sexual maturity and weekly live-weights in laying-type Japanese quail. S. Afr. J. Anim. Sci., 36: 142-148.
CrossRefDirect Link - Sezer, M., 2007. Genetic parameters estimated for sexual maturity and weekly live weights of Japanese quail (Coturnix coturnix japonica). Asian-Australian J. Anim. Sci., 20: 19-24.
Direct Link - Farrag, S.A., A.B. Tanatarov, M.E. Soltan, M. Ismail and O.M. Zayed, 2011. Microsatellite analysis of genetic diversity in three populations of Japanese quail (Coturnix coturnix japonica) from Kazakhstan. J. Anim. Vet. Adv., 18: 2376-2383.
CrossRefDirect Link - Narinc, D., T. Aksoy, E. Karaman, A. Aygun, M.Z. Firat and M.K. Uslu, 2013. Japanese quail meat quality: Characteristics, heritabilities, and genetic correlations with some slaughter traits. Poultry Sci., 92: 1735-1744.
CrossRefDirect Link - Alkan, S., T. Karsli, K. Karabag and A. Galic, 2013. The effects of selection and season on clutch traits and egg production in Japanese quails (Coturnix coturnix japonica) of different lines. SDU J. Faculty Agric., 8: 71-77.
Direct Link - Nath, D.N., F.R. Sheriff, R. Prabakaran and R.A. Rajini, 2011. Response to short term index selection for economic traits in meat type Japanese quail. J. Indian Vet. Assoc., 9: 10-14.
Direct Link - El-Deen, M.B., A.A. Nour, W.S. El-Tahawy and H.A. Fouad, 2015. Effect of selection for weight of early eggs on egg production traits in Japanese quail. Asian J. Poult. Sci., 9: 179-190.
CrossRefDirect Link - Asasi, K. and A.J. Jaafar, 2000. The effect of sex ratio on egg production, fertility and hatchability of Japanese quail. Pajouhesh-Va-Sazandegi, 4: 128-131.
Direct Link - Ramzy, A., D. Aboul-Seoud and M. Aboul-Hassan, 2019. Direct and correlated responses from selection for early sexual maturity in Japanese quail. Arch. Agric. Sci. J., 2: 28-40.
CrossRefDirect Link