
Alaa E. Elkomy
Department of Livestock Research, Arid Lands Cultivating Research Institute, City of Scientific Research
Ahmed M. Abd El-hady
Department of Poultry Production, Faculty of Agriculture, Alexandria University, Alexandsia, Egypt
Osama A. Elghalid
Department of Poultry Production, Faculty of Agriculture, Alexandria University, Alexandsia, Egypt
Asian Journal of Poultry Science
Year: 2015 | Volume: 9 | Issue: 2 | Page No.: 85-96







ABSTRACT
This study was carried out to assess the effect of Boron supplementation on male rabbit’s reproductive performance. A total of 20 V. Line (V.L) adult rabbit bucks were divided into four equal experimental groups (5 bucks for each). Groups 2, 3 and 4 were fed rations supplemented with 17.5, 35 and 70 mg boron/kg feed in the form of boric acid (100, 200 and 400 mg boric acid, respectively) over 8 weeks. The first group was served as a control group. Results obtained by the Boron groups revealed that adding Boron to rabbit bucks rations at any tested doses resulted in a non-significant increase in the ejaculate volume of male rabbit by approximately 25% above the control group. Adding Boron at any dose exhibited a significant increase (p≤0.01) in sperm concentration, total sperm output, sperm motility, live sperm and normal sperm compared to the control group and the high Boron dose had the highest effect on previous measurements compared to the low and medium Boron doses. Opposite trend was shown in the reaction time (libido) and seminal plasma fructose concentration which showed a significant decrease in these parameters in the Boron treated groups compared to the control. Plasma total protein, glucose, total lipids and triglycerides concentration were boosted in the Boron groups compared to the control (p≤0.01) for glucose and triglycerides measurements. On the contrary, treated male rabbits with Boron at any tested doses resulted in a significant reduction in plasma cholesterol concentration. Blood and seminal plasma AST and ALT enzymes activities showed a tendency to decrease by Boron treatments compared to the control group. Treating bucks by different Boron doses resulted in a significant increase (p≤0.01) in Total Antioxidant Capacity (TAC) to reach 110, 128 and 163% of blood plasma and 114, 121 and 122% of seminal plasma compared to the control value. On the other side, adding Boron to bucks rations did not affect the activity of seminal plasma alkaline phosphatase enzyme. Blood testosterone hormone had a tendency to increase in the Boron groups compared to the control group but this increase was not significant. It could be concluded that feeding rabbit bucks on rations containing the studied Boron doses results in a significant improvement in semen quality characteristics and had a positive effect on their physiological status.
PDF Abstract XML References Citation
Received: December 19, 2014;
Accepted: March 07, 2015;
Published: May 15, 2015
How to cite this article
Alaa E. Elkomy, Ahmed M. Abd El-hady and Osama A. Elghalid, 2015. Dietary Boron Supplementation and its Impact on Semen Characteristics and Physiological Status of Adult Male Rabbits. Asian Journal of Poultry Science, 9: 85-96.
DOI: 10.3923/ajpsaj.2015.85.96
URL: https://scialert.net/abstract/?doi=ajpsaj.2015.85.96
DOI: 10.3923/ajpsaj.2015.85.96
URL: https://scialert.net/abstract/?doi=ajpsaj.2015.85.96
INTRODUCTION
Boric acid is derived from Boron, a naturally occurring element found in rocks, soil and water. Boron is universal in the environment and typically is found in the form of borates or combined with other chemicals (WHO., 1998; Woods, 1994; Eisler, 1990). Boric acid products are used on a variety of sites including sewage systems, food and non-food crops, outdoor residential areas and indoor sites such as homes, hospitals and commercial buildings (EPA., 1993).
In the last 20 years, several researches have shown that Boron may have some positive effects for human health (Ku et al., 1993; Brown et al., 1989). In addition, it has been shown that Boron has many regulative roles in macromineral metabolism (Brown et al., 1989; Hegsted et al., 1991), energy metabolism (Hunt et al., 1997) and on the immune system (Hunt and Idso, 1999).
In the field of animal nutrition, the role of some trace minerals is not completely elucidated and contradictory results are reported in the literature as well there is no concrete definition of the real function of Boron and its possible requirements for poultry (broilers, laying hens and breeders) (Fassani et al., 2004). Some functions and relationships of Boron were suggested including improvement of the growth rate, nutritional efficiency, calcium and phosphorus retention in broilers, besides reduction of vitamin D deficiency symptoms (Hunt and Nielsen, 1981). Although, Boron is not considered essential for poultry or most animals, the inclusion of 2 ppm in poultry feed was recommended by NRC (1984). At the molecular level, Boron influences the activities of at least 26 enzymes (Hunt, 1998) and many of these enzymes are essential in energy substrate metabolism. In vitamin D deficient chicks, dietary Boron decreased the abnormally elevated plasma concentrations of pyruvate, hydroxybutyrate and triglycerides that are typically associated with vitamin D deficiency (Hunt et al., 1994).
Boric acid is practically nontoxic to birds acutely (LD50>2510 mg kg-1) and the U.S. EPA. (1993) does not anticipate adverse effects to birds from the use patterns of boric acid products. Wilson and Ruszler (1997) studied the effects of Boron supplementation in broiler diets and suggested that the addition of 50 ppm Boron improved some bone characteristics (resistance to breaking, shear force and ash percentage) but did not influence weight gain. Moreover, low levels of Boron added to pig diets (5 and 10 ppm) were beneficial, resulting in improved weight gain, nutritional efficiency and calcium and phosphorus retention (Goihl, 2002).
It is known that semen profile is directly affected by various environmental, physical and chemical factors (Oliva et al., 2001). There is no enough knowledge about the effect of Boron on male reproduction and semen quality. Armstrong et al. (2002) studied the long-term effects of 5 mg kg-1 Boron supplementation on the reproductive characteristics of gilts, it was reported that Boron did not alter the fertilization rate or embryonic mortality (Armstrong et al., 2002). During the previous studies on humans residing in regions with high Boron content, no negative findings were obtained regarding fertility in a retrospective questionnaire study. According to Korkmaz et al. (2007), regarding male fertility did not report any value for chronic Boron exposure.
The aim of present study was to investigate the impact of dietary Boron supplementation to bucks’ rations on their semen quality and some blood and seminal plasma constituents.
MATERIALS AND METHODS
To investigate the impact of adding Boron to bucks ration on reproductive performance, a total of twenty V. line adult male rabbits (at 10 months of age) were used during spring season (from April to May). Bucks were randomly distributed into 4 homogeneous groups (5 bucks each). The rabbits were individually housed in a naturally ventilated building and kept in individual galvanized wire cages. Batteries were equipped with feeders for pellet rations and automatic drinkers. Animals were kept under similar management and hygienic conditions. All groups were fed ad libitum the same commercial pellet diet, containing 17.27% crude protein and 2640 kcal kg-1.
Groups 2, 3 and 4 were fed rations supplemented with 17.5, 35 and 70 mg boron/kg feed in the form of boric acid (100, 200 and 400 mg boric acid, respectively), over 8 weeks experimental period. The first group was served as a control group that was fed on a basal diet only.
Semen specimens were artificially collected biweekly using an artificial vagina. Reaction time, the time between introducing the teaser doe to the buck and ejaculation (RT/sec) was estimated. The volume of each ejaculate was recorded nearest 0.1 mL (using a graduated collection tube) after removal of the gel mass. Sperm motility was estimated using phase contrast optics at 40xand assessed from 0 to 100%. A weak eosin-formalin (10% formalin) slain solution was used to evaluate the sperm concentration (Elkomy, 2003), using the improved Neubauer haemocytometer slide (Smith and Mayer, 1955). Total sperm output was calculated by multiplying semen ejaculate volume and semen concentration. Seminal plasma initial fructose concentration was determined immediately after semen collection according to Mann (1948). Assessments of live and normal spermatozoa were performed using an eosin-nigrosine blue staining mixture (Blom, 1950). Total number of motile sperm was calculated by multiplying the percentage of motile sperm and total sperm output and Total Motile Normal Sperm (TMNS) {(the product of total sperm outputxmotility (%))xnormal morphology sperm (%)} (Correa and Zavos, 1996).
Seminal plasma was obtained by centrifugation of semen samples at 3500 rpm for 20 min at 4°C and was stored at -20°C until later analysis. Heparinized tubes were used to collect blood samples which obtained from the ear vein of each buck every 2 weeks. Blood samples were centrifuged at 3500 rpm for 20 min to obtain plasma which was stored at -20°C.
Seminal and blood plasma samples were analyzed for Total Protein (TP) by the Biuret method according to Henry et al. (1974). Albumin (Al) concentration was determined by the method of Doumas et al. (1977). Total lipids (TLp) were measured according to Frings et al. (1972). The enzymes activities of Aspartate aminotransferase (AST) and Alanine aminotransferase (ALT) were assayed by the method of Reitman and Frankel (1957). Seminal plasma Alkaline phosphatase (AlP) activity was measured at 405 nm by the formation of para-nitrophenol from para-nitrophenylphosphate as a substrate (Principato et al., 1985). Blood triglycerides and Total cholesterol were determined according to Fossati and Prencipe (1982) and Richmond (1973). Lipid peroxidation biomarkers such as Malondialdehyde (MDA) were assayed in the blood and seminal plasma according to Conti et al. (1991). Total antioxidant capacity was measured according to Erel (2004). Blood thyroxin hormone concentration was determined using immunoassay Elisa kit. Blood testosterone concentration was measured using immunoassay commercial Elisa kit (DRG international, Inc., USA) (Tietz, 1986).
Statistical analysis: Data were analyzed as a complete randomized design (Steel and Torrie, 1980) using the general linear model procedures of SAS (2001). Significance of the effects was tested at level p≤0.05 and p≤0.01 with the appropriate F statistic. Duncan’s multiple range tests was used to detect any significant differences among the experimental means (Duncan, 1955).
RESULTS AND DISCUSSION
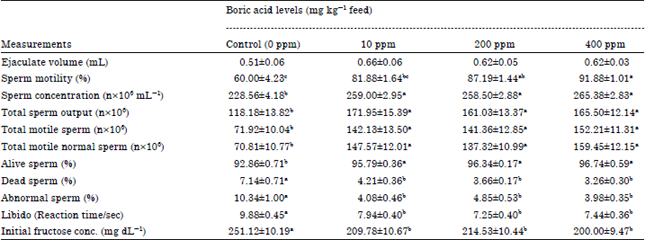
Semen characteristics: Supplemented bucks' rations with Boron was conducted to improve libido for bucks (measured by the reaction time/ seconds) which had less time before ejaculation compared to the control group and this effect was significant (p≤0.01) (Table 1). On the other hand, no significant differences were found between Boron groups in reaction time. The previous effect may be due to increased testosterone level in the treated rabbits’ groups, as they had higher level of testosterone hormone than the control group (Table 2).
| Table 1: | Dietary Boron supplementation and its impact on libido, semen characteristics, sperm output and Initial fructose concentration of adult male rabbits |
 | |
| a,b,cMeans in same rows with different superscripts are significantly different (p≤0.05) | |
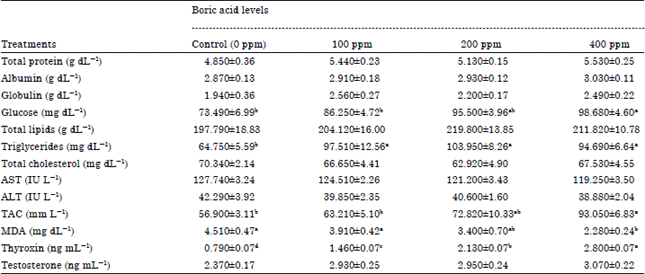
| Table 2: | Dietary Boron supplementation and its impact on blood biochemical constituents and testosterone concentration of adult male rabbits |
 | |
| a,b,cMeans in same rows with different superscripts are significantly different (p≤0.05) | |
The previous finding was in agreement with that found by Elkomy et al. (2008) and Kamel et al. (2009), who reported a good relation between increased testosterone hormone concentration and increase libido in male rabbits.
Data of semen quality characteristics (Ejaculate volume, Sperm motility, sperm concentration, live and dead sperm, abnormal sperm, total sperm output, total motile sperm and total motile normal sperm) are illustrated in Table 1. Ejaculate volume of bucks which put on rations supplemented with Boron was increased by 29.4, 21.5 and 21.5% for 100, 200 and 400 ppm boric acids groups respectively, the control group mean but this increase was not significant. Adding Boron in rabbit’ bucks rations resulted in a significant increase (p = 0.01) in sperm concentration at any studied levels to reach 13.3, 13.0 and 16.2% for 100, 200 and 400 ppm boric acid levels respectively, over the control group. Also, fed rabbits’ bucks on diet supplemented with different levels of Boron resulted in a significant increase (p≤0.01) Total Sperm Output (TSO), Total Motile Sperm (TMS) and Total Motile Normal Sperm (TMNS) per ejaculate compared to the control group. Moreover, the increased semen ejaculate volume in the Boron groups was correlated with a significant increase in sperm concentration (SCon.) and TSO in all Boron treated groups. According to Kamel et al. (2009), increase in ejaculate volume was necessary for sperms to provide them by nutritious elements that are needed for their live.
Adding Boron at any inclusion level resulted in a significant increase (p≤0.01) in live sperm to reach 3.1, 3.7 and 4.1% above the control group mean. Supplemented bucks’ rations with Boron at any studied level conducted to produce more normal shape sperm than the control group and this effect was significant (p≤0.01). The opposite trend was shown in the dead and abnormal sperm which showed a significant decrease (p≤0.01) in all Boron levels. The percentage of abnormal sperm was significantly decreased in the all Boron groups to reach -60.5, -53.1 and -61.5 less than the control group. On the other hand, no significant differences were found between Boron groups on the previous semen quality measurements. Data revealed that improvement in percentage of live and motile sperm that found in the Boron groups were conducted to a significant increase (p≤0.01) in TMS and TMNS per ejaculate compared to the control group. The positive effect of Boron on the previous measurements may be attributed to the Boron affects several enzymes that enhance and activate the spermatogenesis in seminiferous tubules to produce a complete sperm. Whereas, at the molecular level, Boron influences the activities of at least 26 enzymes (Hunt, 1998) and many of these enzymes are essential in energy substrate metabolism. In addition, it has been shown that Boron has many regulative roles in macromineral metabolism (Brown et al., 1989; Hegsted et al., 1991), energy metabolism (Hunt et al., 1997) and on the immune system (Hunt and Idso, 1999). Data of fructose concentration in seminal plasma (Table 1) revealed that there was a significant decrease in fructose concentration (p≤0.05) after feeding bucks on different levels of boric acid compared to the control group. Decreased semen fructose concentration in the DTP treated groups may be due to increase sperm concentration per ejaculate (Table 1) that consumed more fructose to meet the needs of the energy required for metabolism.
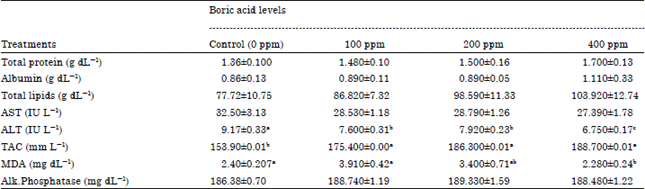
Seminal plasma characteristics: Data of seminal plasma Total Protein (TP), Albumin (Alb), Total Lipids (TLp), activities of aspartate amino transferase (AST) and alanine amino transferase (ALT), alkaline phosphatase enzyme activity (AlP), Total Antioxidant Capacity (TAC) and Malondialdehyde (MDA) are presented in Table 3.
| Table 3: | Dietary Boron supplementation and its impact on seminal plasma biochemical constituents of adult male rabbits |
 | |
| a,b,cMeans in same rows with different superscripts are significantly different (p≤0.05) | |
There was a non-significant increase in seminal plasma TP and Alb by 8.8, 10.2 and 25% of TP and 3.4, 3.4 and 29% of Alb (for 100, 200 and 400 mg boric aid/kg feed, respectively) due to feeding bucks the rations supplemented with different Boron levels compared to the control group and the highest treated Boron group had the highest TP and Alb values. Treated bucks with Boron at any studied level resulted in increasing seminal plasma total protein and albumin concentration which was associated with an improvement in semen quality and this finding was corresponding with Elkomy et al. (2008), they reported an increase in seminal plasma total protein and albumin concentrations were showed in high fertile male rabbits compared to low fertile rabbits and this increase was associated with increase in their seminal quality measurements. Many studies have shown that low content of seminal plasma proteins is associated with poor semen quality (White et al., 1987; Ashworth et al., 1994). Taha et al. (2000) revealed that there was a positive relationship between semen quality and level of seminal plasma total proteins. Similar results were found by Osama and El-Sahn (2006), they found a positive relationship between increasing seminal plasma total proteins and albumin and increasing total number of sperm output. Kulkarni et al. (1996) showed that, seminal plasma total protein is mainly composed of albumin and globulin, in addition of small quantities of non-protein nitrogen, amino acids and peptides. These compounds make up the amphoteric property of seminal plasma proteins, thus, low protein content in seminal plasma reduce its buffering capacity and in turn semen quality (Dhami et al., 1994).
It can be noticed that there was a decrease in seminal plasma activities of AST and ALT due to feeding bucks on Boron rations at any inclusion level compared to untreated bucks and this effect was significant (p≤0.01) with ALT only. additionally, the high Boron level had the lowest transaminases enzymes activities (AST and ALT). The transaminases activities in semen are a good indicator of semen quality because they measures sperm membrane stability (Kelso et al., 1997; Zedda et al., 1996). Also, the increments of the activities of AST and ALT in seminal plasma are mainly due to the leakage of these enzymes (Navarro et al., 1993). Yousef and Zeitoun (1998) found that there were negative correlation coefficients between decreased sperm motility on one side and AST and ALT release on the other side. They reported that the activities of these enzymes could be used as an indicator of sperm integrity. Yousef et al. (2003) reported that there was a negative correlation between increased ALT and AST activities and decrease ejaculate volume, sperm concentration, total sperm output, sperm motility index, total motile sperm, therefore, the decrease in the activities of these enzymes coincided with the increase of semen quality. Similar results were found by Elkomy et al. (2008), Kamel et al. (2009) and El-Sbeiy et al. (2008), they reported that the AST and ALT enzymes activities were significantly decreased in bucks that produced high motile sperm.
Supplementing bucks’ rations with Boron at any studied level resulted in a gradual and non-significant increase in seminal plasma TLp concentration by 11.7, 26.8 and 33.7% above the control group. In addition, there was a significant effect of adding Boron to bucks' rations on TAC and MDA. Whereas, feeding bucks on rations supplemented with different Boron levels (100, 200 and 400 mg boric aid/kg feed) significantly increased (p≤0.01) TAC by 14, 21 and 22.6% above the control value and this effect was in a dose-dependent manner. In contrary, the influence of dietary Boron supplementation on the measured biomarkers of oxidative stress (MDA) showed that treading bucks with Boron at any tested doses had significantly reduced (p≤0.01) MDA by 5.8, 14.1 and 40.4.6% compared to the control group. Boron treatment resulted in the increase in TAC levels in seminal plasma that protect its lipids content and sperm membrane against lipid peroxidation. According to Niedernhofer et al. (2003), MDA is an endogenous genotoxic product of enzymatic and oxygen radical-induced lipid peroxidation. Oxidative stress causes damage to biologic macromolecules (such as nucleic acids, membrane lipids and proteins) and disorder of normal metabolism and physiology (Roberts and Sindhu, 2009; Pohanka et al., 2009).
Adding Boron to bucks rations with different studied levels did not had effect on seminal plasma ALP enzyme activity in all groups treated compared to the control group. The phosphatases enzymes in semen play an important role in transamination and phosphorylation processes in sperm metabolism and thus explain the differences observed in the semen quality (Dhami et al., 1994).
From the results of semen quality and seminal plasma characteristics it could be noticed that adding Boron to rabbit bucks' rations had a beneficial effects on spermatogenesis in seminiferous tubules to produce a complete and motile sperm, In addition, it stimulated the secondary sexual gland to secrete the seminal plasma components which is necessary for sperm life. Whereas, Boron influences the activities of at least 26 enzymes (Hunt, 1998) and many of these enzymes are essential in energy substrate metabolism.
Blood biochemical constituents: Blood plasma protein constituents (total protein, albumin and globulin) of bucks’ groups that were fed Boron rations showed an increase in these measurements than the control group and these increases were not significant (Table 2). Thus, these increases were 12.1, 5.7 and 14% of total protein, 1.4, 2.1 and 5.5% of albumin and 31.9, 13.4 and 28.3% of globulin concentration above the control value for 10, 200 and 400 mg boric acid/kg feed, respectively. The present data revealed that Boron administration had a beneficial effect on rabbits’ health and immune status, whereas, the protein profile (Total protein, Albumin and globulin concentrations) was increased in Boron bucks groups and this increase was necessary to meet the vital physiological processes.
Treated bucks with different levels of Boron conducted to a gradual and significant increase in blood plasma glucose concentration (p≤0.01) and this increase was 17.3, 29.9 and 34.2% above the control value. Increasing plasma glucose concentration may be due to the increase thyroid gland activity, whereas, the thyroxin hormone secretion from thyroid gland was gradually and significantly increased with increase Boron level.
From Table 2 it could be noticed that there was an increase in blood plasma total lipids and triglycerides concentrations due to feeding bucks on rations supplemented with different Boron levels (10, 200 and 400 mg boric acid/kg feed) compared to the control group that fed the basal diet and this increase was significant for triglycerides only (p≤0.01). Moreover, data revealed that the medium Boron level (200 mg boric acid/kg feed) had the highest values of these two parameters than the low and high Boron levels. On the other side, blood plasma cholesterol concentration showed a non-significant decrease in all Boron groups than the control group and this decrease was 5.2, 10.5 and 4% less than the control value. The previous study of Bakken and Hunt (2003) supposed that the dietary Boron influences energy substrate metabolism in a wide variety of biological species including humans. In addition, Scorei and Cimpoiasu (2006) demonstrated that Boron in animals are still not well unknown but it might be involved in metabolism and utilization of various elements, glucose, triglycerides and reactive oxygen. The improved antioxidant status protect against lipid oxidation in animals (Cai and Wei, 1996).
Liver aminotransaminase enzymes activities (AST) and ALT) showed a gradual decrease when bucks were fed on rations supplemented with different Boron levels compared to the basal diet group and this decrease was non-significant. From the previous studies, serum AST activity was decreased by 100, 200 and 400 mg boron/kg feed supplementation in layer hens (Eren and Uyanik, 2007). The AST in rats (Hunt and Herbel, 1991) and both AST activities in mice (Ince et al., 2012) were also decreased by Boron. According to Hunt and Herbel (1991) and Ince et al. (2012), the decreases in AST and ALT enzymes activities by Boron supplementation may result from the protective effects of the Boron on normal liver metabolism. The same finding was found by Eren et al. (2012) when they fed broiler chicks on 500, 750 and 1000 mg boron/kg feed. Data for plasma (TAC and MAD are shown in Table 2. There was a significant effect of adding Boron to bucks’ rations on Total Antioxidant Capacity (TAC) and malondialdehyde (MDA) (p≤0.01). Whereas, adding Boron to rations at any inclusion level (100, 200 and 400 mg boric acid/kg feed) gradually boosted TAC by 11.9, 27.9 and 63.5% over the control group and this effect was highly significant with 400 mg boron/kg feed only. The increase TAC level in Boron groups was offset by a gradually decreased in blood MDA level which reached 13.3, 24.6 and 49.4% less than the control group and this effect was dose-dependent. Also, it could be concluded that the highest Boron level had significantly lowest MDA level than the control group or the other two Boron treatment. Increased TAC in Boron groups is an indicator for protection against oxidative statuses which reflected on and decrease MDA level in treated groups and this decrease was an indicator for protection against lipids peroxidation. According to Nielsen et al. (1997), MDA concentration in plasma is one of the most frequently biomarkers providing an indication of the overall lipid peroxidation level and the MDA is one of several byproducts of lipid peroxidation processes.
Thyroid hormone secretion (T4) showed a gradually significant increase (p≤0.01) due to treated bucks with Boron at any studied levels compared to the control group. The increase thyroid gland activity and its thyroxin hormone secretion in the Boron groups may be due to the effect of Boron on stimulate energy metabolism. According to Hunt et al. (1997), Boron has many regulative roles in energy metabolism (Hunt et al., 1997), moreover, Boron influenced the activities of at least 26 enzymes (Hunt, 1998) and many of these enzymes are essential in energy substrate metabolism.
Treating male rabbits with 100, 200 and 400 mg boric acid/kg feed boosted blood testosterone concentration to reach 23.6, 24.4 and 29.5%, respectively above the untreated group values which was not significant. Naghii and Samman (1993) noted that the mechanism of Boron action may be mediated by increasing the concentration of steroid hormones such as testosterone and beta-estradiol.
CONCLUSION
It is clear from the present results that supplementing bucks’ rations with boric acid as a source of Boron at any studied level improved semen characteristics and had positive effect on semen quality. Whereas, inclusion of Boron to bucks' rations resulted in a significant increase in sperm concentration, total sperm output, sperm motility, live sperm and normal sperm compared to the control group and the high Boron dose had the highest effect on the previous measurements compared to the low and medium Boron doses. Moreover, treating bucks by different Boron doses had a positive effect on their physiological status. Blood testosterone hormone had a tendency to increase in the Boron groups compared to the control group but this increase was not significant.
REFERENCES
- Armstrong, T.A., W.L. Flowers, J.W. Spears and F.H. Nielsent, 2002. Long-term effects of boron supplementation on reproductive characteristics and bone mechanical properties in gilts. J. Anim. Sci., 80: 154-161.
CrossRefDirect Link - Ashworth, P.J., R.A. Harrison, N.G. Miller, J.M. Plummer and P.F. Watson, 1994. Survival of ram spermatozoa at high dilution: Protective effect of simple constituents of culture media as compared with seminal plasma. Reprod. Fertil. Dev., 6: 173-180.
CrossRefDirect Link - Bakken, N.A. and C.D. Hunt, 2003. Dietary boron decreases peak pancreatic in situ insulin release in chicks and plasma insulin concentrations in rats regardless of vitamin D or magnesium status. J. Nutr., 133: 3577-3583.
Direct Link - Blom, E., 1950. A one-minute live-dead sperm stain by means of eosin-nigrosin. Fertil. Steril., 1: 176-177.
Direct Link - Brown, T.F., M.E. McCormick, D.R. Morris and L.K. Zeringue, 1989. Effects of dietary boron on mineral balance in sheep. Nutr. Res., 9: 503-512.
CrossRefDirect Link - Cai, Q. and H. Wei, 1996. Effect of dietary genistein on antioxidant enzyme activities in SENCAR mice. Nutr. Cancer, 25: 1-7.
CrossRefDirect Link - Conti, M., P.C. Morand, P. Levillain and A. Lemonnier, 1991. Improved fluorometric determination of malonaldehyde. Clin. Chem., 37: 1273-1275.
Direct Link - Correa, J.R. and P.M. Zavos, 1996. Preparation and recovery of frozen-thawed bovine spermatozoa via various sperm selection techniques employed in assisted reproductive technologies. Theriogenology, 46: 1225-1232.
CrossRefDirect Link - Dhami, A.J., K.L. Sahni, G. Mohan and R.P. Tripathi, 1994. Comparative evaluation of initially static and motile semen ejaculates from Friesian and Murrah buffalo bulls for physico-morphological, biochemical, enzymatic and mineral constituents of seminal plasma. Indian J. Anim. Sci., 64: 926-932.
Direct Link - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - Erel, O., 2004. A novel automated method to measure total antioxidant response against potent free radical reactions. Clin. Biochem., 37: 112-119.
CrossRefPubMedDirect Link - Fassani, E.J., A.G. Bertechini, J.A.K. Brito, R.K. Kato, E.T. Fialho and A. Geraldo, 2004. Boron supplementation in broiler diets. Revista Brasileira Ciencia Avicola, 6: 231-237.
CrossRef - Nielsen, F., B.B. Mikkelsen, J.B. Nielsen, H.R. Andersen and P. Grandjean, 1997. Plasma malondialdehyde as biomarker for oxidative stress: Reference interval and effects of life-style factors. Clin. Chem., 43: 1209-1214.
PubMedDirect Link - Fossati, P. and L. Prencipe, 1982. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem., 28: 2077-2080.
CrossRefPubMedDirect Link - Frings, C.S., T.W. Fendley, R.T. Dunn and C.A. Queen, 1972. Improved determination of total serum lipids by the sulfo-phospho-vanillin reaction. Clin. Chem., 18: 673-674.
CrossRefPubMedDirect Link - Hegsted, M., M.J. Keenan, F. Siver and P. Wozniak, 1991. Effect of boron on vitamin D deficient rats. Biol. Trace Elem. Res., 28: 243-255.
CrossRefDirect Link - Henry, R.J., D.C. Cannon and W. Winkelman, 1974. Clinical Chemistry Principles and Techniques. 11th Edn., Harper and Row Publishers, New York, Pages: 1629.
Direct Link - Hunt, C.D. and J.L. Herbel, 1991. Boron affects energy metabolism in the streptozotocin-injected, vitamin D3-deprived rat. Magnes. Trace Elem., 10: 374-386.
PubMed - Hunt, C.D., 1998. Regulation of enzymatic activity: One possible role of dietary boron in higher animals and humans. Biol. Trace Elem. Res., 66: 205-225.
PubMed - Hunt, C.D. and F.H. Nielsen, 1981. Interaction between Boron and Cholecalciferol in the Chick. In: Trace Element Metabolism in Man and Animals: Proceedings of the Fourth International Symposium on Trace Element Metabolism in Man and Animals (TEMA-4), held in Perth, Western Australia, 11-15 May 1981, Gawthorne, J., J.M. Howell and C.L. White (Eds.). Australian Academy of Science, Canberra, Australia, ISBN: 978-3642682711, pp: 597-600.
Direct Link - Hunt, C.D., J.L. Herbel and F.H. Nielsen, 1997. Metabolic responses of postmenopausal women to supplemental dietary boron and aluminum during usual and low magnesium intake: Boron, calcium and magnesium absorption and retention and blood mineral concentrations. Am. J. Clin. Nutr., 65: 803-813.
Direct Link - Ince, S., H. Keles, M. Erdogan, O. Hazman and I. Kucukkurt, 2012. Protective effect of boric acid against carbon tetrachloride-induced hepatotoxicity in mice. Drug Chem. Toxicol., 35: 285-292.
CrossRefDirect Link - Kamel, K.I., A.E. Elkomy and M.S. El-Sbeiy, 2009. The Androgenic action of Gibberellic Acid (GA3) on reproductive performance of New Zealand white rabbit bucks World J. Agric. Sci., 5: 40-48.
Direct Link - Kelso, K.A., A. Redpath, R.C. Noble and B.K. Speake, 1997. Lipid and antioxidant changes in spermatozoa and seminal plasma throughout the reproductive period of bulls. J. Reprod. Fert., 109: 1-6.
CrossRefPubMedDirect Link - Korkmaz, M., U. Sayli, B.S. Sayli, S. Bakirdere, S. Titretir, O.Y. Ataman and S. Keskin, 2007. Estimation of human daily boron exposure in a boron-rich area. Br. J. Nutr., 98: 571-575.
CrossRefDirect Link - Ku, W.W., R.E. Chapin, R.N. Wine and B.C. Gladen, 1993. Testicular toxicity of Boric Acid (BA): Relationship of dose to lesion development and recovery in the F344 rat. Reprod. Toxicol., 7: 305-319.
CrossRefDirect Link - Niedernhofer, L.J., J.S. Daniels, C.A. Rouzer, R.E. Greene and L.J. Marnett, 2003. Malondialdehyde, a product of lipid peroxidation, is mutagenic in human cells. J. Biol. Chem., 278: 31426-31433.
CrossRefDirect Link - Mann, T., 1948. Fructose content and fructolysis in semen. Practical application in the evaluation of semen quality. J. Agric. Sci., 38: 323-331.
CrossRefDirect Link - Eren, M., F. Uyanik, B.K. Guclu and A. Atasever, 2012. The influence of dietary boron supplementation on performance, some biochemical parameters and organs in broilers. Asian J. Anim. Vet. Adv., 7: 1079-1089.
CrossRef - Naghii, M.R. and S. Samman, 1993. The role of boron in nutrition and metabolism. Prog. Food Nutr. Sci., 17: 331-349.
PubMed - Navarro, C.M., P.M. Montilla, A. Martin, J. Jimenez and P.M. Utrilla, 1993. Free radical scavenger and antihepatotoxic activity of Rosmarinus tomentosus. Planta Med., 59: 312-314.
CrossRefPubMedDirect Link - Oliva, A., A. Spira and L. Multigner, 2001. Contribution of environmental factors to the risk of male infertility. Hum. Reprod., 16: 1768-1776.
CrossRefDirect Link - Pohanka, M., H. Bandouchova, J. Sobotka, J. Sedlackova, I. Soukupova and J. Pikula, 2009. Ferric reducing antioxidant power and square wave voltammetry for assay of low molecular weight antioxidants in blood plasma: Performance and comparison of methods. Sensors, 9: 9094-9103.
CrossRefDirect Link - Principato, G.B., M.C. Aisa, V. Talesa, G. Rosi and E. Giovannini, 1985. Characterization of the soluble alkaline phosphatase from hepatopancreas of Squilla mantis L. Comp. Biochem. Physiol. Part B: Comp. Biochem., 80: 801-804.
CrossRefDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Roberts, C.K. and K.K. Sindhu, 2009. Oxidative stress and metabolic syndrome. Life Sci., 84: 705-712.
CrossRefDirect Link - Scorei, R. and V. Cimpoiasu, 2006. Boron enhances the thermostability of carbohydrates. Origins Life Evol. Biospheres, 36: 1-11.
CrossRefDirect Link - Smith, J.T. and D.T. Mayer, 1955. Evaluation of sperm concentration by the hemacytometer method; comparison of four counting fluids. Fertil. Steril., 6: 271-275.
PubMed - Taha, T.A., E.l. Abdel-Gawad and M.A. Ayoub, 2000. Monthly variations in some reproductive parameters of Barki and Awassi rams throughout 1 year under subtropical conditions. 1. Semen characteristics and hormonal levels. Anim. Sci., 71: 317-324.
Direct Link - Tietz, N.W., 1986. Textbook of Clinical Chemistry. W.B. Saunders, Philadelphia, USA., ISBN-13: 9780721688862, pp: 1271.
CrossRefDirect Link - White, I.G., P. Goh and J.K. Voglmayr, 1987. Effect of male reproductive tract fluids and proteins on the metabolism and motility of ram spermatozoa. Syst. Biol. Reprod. Med., 19: 115-125.
CrossRefDirect Link - Wilson, J.H. and P.L. Ruszler, 1997. Effects of boron on growing pullets. Biomed. Trace Elem. Res., 56: 287-294.
CrossRefPubMedDirect Link - Woods, W.G., 1994. An introduction to boron: History, sources, uses and chemistry. Environ. Health Perspect., 102: 5-11.
PubMed - Yousef, M.I., G.A. Abdallah and K.I. Kamel, 2003. Effect of ascorbic acid and Vitamin E supplementation on semen quality and biochemical parameters of male rabbits. Anim. Reprod. Sci., 76: 99-111.
CrossRefPubMedDirect Link - Zedda, M.T., P.P. Bini, S. Pau and U. Sbernardori, 1996. Constituents of seminal plasma and blood serum of the ram. Bull. Soc. Italy Biol. Sper., 72: 227-230.
PubMed