
A.M. Hanafy
Department of Animal Production, Faculty of Agriculture, Suez Canal University, Ismailia, 41522, Egypt
H.A. Khalil
Department of Animal Production, Faculty of Agriculture, Suez Canal University, Ismailia, 41522, Egypt
Asian Journal of Poultry Science
Year: 2015 | Volume: 9 | Issue: 4 | Page No.: 223-232







ABSTRACT
The present study was conducted to determine the effects of chronic Dexamethasone (DEX) administration on physiological traits and semen characteristics in male Japanese quail breeders. Forty five 20 week old male quail were distributed into 3 experimental groups. All groups were fed on the same basal diet and administered with different levels of DEX, at 0 (as a control group), 0.25 and 0.50 mg/bird/day orally for 14 consecutive days. Body weight, feed intake, cloacal gland area and fertility percentage were recorded. Also, male genital organs were investigated and semen characteristics were estimated at the end and after 28 days of administration ended. Serum testosterone and corticosterone levels were analyzed by ELISA. The results revealed that, there were no significant effects of ingested DEX at different levels on body weight, feed intake and testes weight of the male Japanese quail breeders. However, the administration of DEX at 0.25 and 0.50 mg/bird decreased significantly cloacal gland area, foam production, sperm motility and viability and testosterone level. Also, treatments with DEX increased Time of Sexual Libido (TSL), Methylene Blue Reaction Time (MBRT) and serum corticosterone level compared with control group. Also, significant decrease in fertility percentage was observed after male exposure to DEX. Even after 4 weeks from administration ended, libido and sperm motility were significantly impaired. Also, significant negative correlations were found between serum corticosterone level and most studied traits. Our results indicated that chronic DEX administration had markedly effects on the reproductive performance of male quail for several weeks even after exposure to a stressors ends.
PDF Abstract XML References Citation
Received: June 15, 2015;
Accepted: July 27, 2015;
Published: September 15, 2015
How to cite this article
A.M. Hanafy and H.A. Khalil, 2015. Influence of Chronic Dexamethasone Administration on Reproductive Parameters and Semen Traits in Male of Japanese Quail. Asian Journal of Poultry Science, 9: 223-232.
DOI: 10.3923/ajpsaj.2015.223.232
URL: https://scialert.net/abstract/?doi=ajpsaj.2015.223.232
DOI: 10.3923/ajpsaj.2015.223.232
URL: https://scialert.net/abstract/?doi=ajpsaj.2015.223.232
INTRODUCTION
Climate change, human disturbance and poor housing quality are some of environmental factors that can lead to chronic stress in birds (Scheuerlein et al., 2001; Schoech et al., 2009). Chronic stress that lasts for days or even weeks can have deleterious effects such as inhibiting reproductive functions and reducing body weight (Breuner et al., 2008). One ways to study the effects of stress on the performance of the birds is treated birds with exogenous glucocorticoids to stimulate its rise in plasma that occur during stress (Tasker and Herman, 2011). Glucocorticoid concentrations can then be related to physiological or behavioural changes that occur to restore homeostasis (Sapolsky et al., 2000). The main out come of this pathway is elevated blood corticosterone, the primary glucocorticoid in birds (Silverin, 1998; Romero and Reed, 2008).
Dexamethasone (DEX) is a potent synthetic analogue of hydrocortisone and its administration is mimics the adverse effects of increased corticosterone. Using DEX as an artificial stress caused deterioration in most reproductive and physiological performance of birds such as male domestic fowls (Eid et al., 2006), laying quails (Aengwanich and Chinrasri, 2003) and pigeons (Westerhof et al., 1996). As shown recently, DEX administration produces large amounts of reactive species and is suggested to induce oxidative stress after treatment of broiler (Gao et al., 2010) and laying hens (El-Habbak et al., 2005; Eid et al., 2008). Many researches have been provided important information about how birds are affected by the elevated plasma corticosterone levels that occur during chronic stress (Calandreau et al., 2011; Gao et al., 2010; Lin et al., 2004). However, it is unclear that can reproductive and physiological function of male birds be permanently affected by chronic stress, or male birds can quickly recover from the deleterious consequences of sustained elevation in blood corticosterone concentrations.
Therefore, the aims of the present study were to determine the effects of chronic DEX treatment on physiological traits and semen characteristics of male Japanese quail both at end of treatment and up to 28 days after treatment ended.
MATERIALS AND METHODS
This experiment was carried out at the Poultry Farm, Department of Animal Production, Faculty of Agriculture, Suez Canal University, Ismailia, Egypt during spring season 2014.
Experimental design: At 20 weeks of age, 45 healthy male Japanese quails were individually weighed and randomly assigned to three experimental groups (15 male each). All males were housed in individually battery cages 20×20×25 cm. During the experimental period, male quails were provided with water and quail breeder ration ad libitum (20% CP, 3000 Kcal kg–1 ME). Diet composition was formulated to meet the recommended nutrient requirements (NRC., 1994). Ambient temperature averaged 24±2°C with relative humidity 50-60% and a 16 h light: 8 h dark-cycle was applied. All experiments were performed in accordance with institutional guidelines concerning animal use. Dexamethasone administration began after 7 days from housing of males in individual cages. Dexamethasone was mixed with mash diet and then packed in 00-capsule to give concentrations of 0 (control group), 0.25 and 0.50 mg/bird/day. Once the treatment regime was initiated (day 1), each male was orally received one capsule per day for 14 consecutive days. The duration of experiment was 6 weeks, the 1st and 2nd weeks were treatment period and another 4 weeks were recorded as a recovery period.
Measurements and laboratory analysis: Live body weight, cloacal gland area and foam production were recorded at the beginning, the end of administration period and after recovery period. Feed consumption was measured weekly. The cloacal gland areas (height and width per mm) were measured by using a caliper for all the males (Biswas et al., 2007). The foam produced by each male was collected for three times with time interval 15 min. Quantitative measurements on cloacal gland foam production were conducted immediately using an electronic analytical balance.
Semen collection: Semen samples were collected at the end of administration period and after recovery period according to Bakst and Cecil (1997). Briefly, the female birds were bring to the male in the same cage. When the male mount the female to mating, the male taking immediately before ejaculation of semen. Each male was gently restrained on the palm of the left hand and foam was squeezed out before semen collection. The lumber region towards tail was massaged 3-4 times smoothly and applied gentle pressure on either side of the vent by using thumb and fore finger. The semen was collected by micropipette. To prevent contamination with foam, feaces and watery fluid, cloacal gland was cleaned gently with tissue paper.
Semen evaluation: Time of sexual libido was measured in terms of reaction time in seconds and was estimated from the time of female was placed inside the male’s cage up to the point when the male started to mount the female. Semen characteristics were estimated immediately after collection in each ejaculate such as; semen ejaculates volume, initial motility (%), sperm concentration, live and dead sperm percentage. Ejaculate volume was measured to the nearest 1 μL using a graduated collection micropipette. Sperm motility (%) was estimated microscopically at 400X by using a drop of diluted semen with a drop of warm 0.9% saline solution (NaCl) and covered with cover slip on a warm slide. Sperm concentration per ejaculate was determined using the hemocytometer (Golzar Adabi et al., 2007). Metabolic activity of quail spermatozoa was confirmed by conducting modified Methylene Blue Reaction Time (MBRT) test as described by Shit et al. (2010). Briefly, after semen collection, 5 μL of semen was diluted with isotonic solution at 1:10 ratios in Eppendorf tube. Total volume of 5 μL methylene blue solution (50 mg % w/v) was added to the diluted semen. Immediately after gently mixing, the upper layer was covered with 0.2 mL mineral oil and incubated at 37°C in water bath. Time was estimated from addition of methylene blue till its blue colour turned into white colour. The percentage of dead spermatozoa was estimated using eosin-nigrosin stain. Percentage of dead spermatozoa was estimated by counting the number present in 100 sperm in different fields on the slide.
Blood sample and testes weight: At the end of administration period, five male quails from each group were slaughtered by cutting the jugular vein. The blood samples were centrifuged at 3000 rpm for 15 min and serum obtained was stored at -20oC for further analysis. Serum testosterone and corticosterone levels were determined by ELISA kits manufactured by DiaMetra, Spello-Perugia, Italy and IBL, Hamburg, Germany, respectively. The sensitivity of the assay was 1.631 ng mL–1 and 70 pg mL–1 and the percentage of recovery was 95-103 and 100-105% for corticosterone and testosterone, respectively. The intra- and interassay coefficients of variation were 4.08 and 5.54% and 5.8 and 10.5% for corticosterone and testosterone, respectively. Immediately after blood collection, the abdominal cavity was opened and the testes were removed and weighed using an electronic analytical scale.
Fertility test: At the end of administration period, the female birds were housed with equal number of male (1:1) for natural matting. After a week of adaptation, hatching eggs were collected for sequential 6 days. Hatching eggs were collected daily and stored at 18°C and 65% relative humidity up to 5 days before they were placed in the artificial incubator. Hatching eggs were set in incubator for 7 days on 37.5°C and 60% relative humidity. The fertility was assured by broken open all eggs and fertility percentage was calculated by the ratios of number of fertile eggs to the number of total hatching eggs set in the incubator.
Statistical analysis: Data were analyzed using the General Linear Model (GLM) procedure of SPSS (SPSS., 2009). Differences among means were detected using Duncan’s new multiple test (Duncan, 1955). Correlation coefficients among traits were estimated.
RESULTS
The results showed that administration of DEX for 2 consecutive weeks at levels of 0, 0.25,or 0.50 mg/bird/day did not statistically significant affected in mean Body Weight (BW), feed intake or testes weight of mature male quail. Although lose BW and feed intake were numerically higher in control group compared with treated groups, difference between treatments was not significant (Table 1). Similar results were observed in BW after 4 weeks of recovery period.
Also, significant difference (p≤0.05) was found in cloacal gland area after DEX administration and the lowest value was observed in 0.50 mg group. In same pattern, significant difference in foam production could be detected between 0.50 mg DEX exposure male and control male after administration period. Allowing stressed males for 4 weeks recovery period enhanced the negative effect of DEX since cloacal gland area as well as cloacal foam production increased again (Table 1). There were significant differences (p≤0.05) in time of sexual libido, ejaculate volume, initial motility, sperm concentration, MBRT and live spermatozoa after administration among treatments (Table 2). The worst sexual libido was detected in the 0.50 mg treatment group after the end of treatment. This harmful effect of DEX administration did not reduced even after allowing males for recovery period of 4 weeks. Contrary, the highest ejaculate volume of semen was collected from 0.25 mg treatment birds. Although, a numerical increase in ejaculate volume has been observed due to DEX administration, the difference was not significant compared to control. Similarly, no significant difference in semen volume was observed after recovery period among treatments.
On the other hand, the results indicated a significant reduction in motility percentage by increasing of DEX dose. The lowest motility percentage was observed with 0.50 mg treated group (79.43%). Although, a relative improve in motility percentage was observed after recover period of treated male, the difference was still significant compared to control birds. The same trend was observed in MBRT, except that administration 0.25 mg did not increase MBRT to a time more than that of control group. The concentration of spermatozoa per ejaculate increased significantly while percentage of live sperm decreased significantly in DEX compared with control group. These significant differences were disappeared after recovery period.
| Table 1: | Body weight, feed intake, total testes weight, cloacal gland area and foam production of male quails as affected by different levels of DEX treatments |
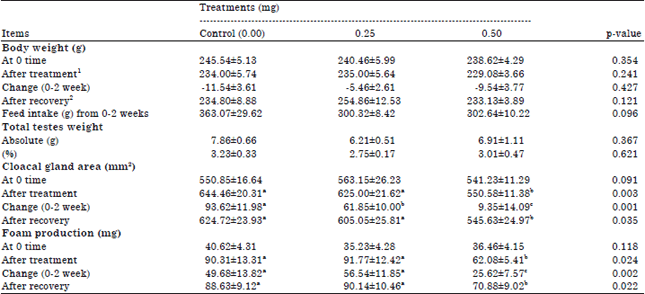 | |
| a,bMeans in any row with no common superscript differ (p≤0.05), 1After 2 weeks from DEX administration. 2After 4 weeks from DEX administration ended, values are given in Mean±SE | |
| Table 2: | Sexual libido, semen characteristics and fertility percentage of male quails as affected by different levels of DEX treatments |
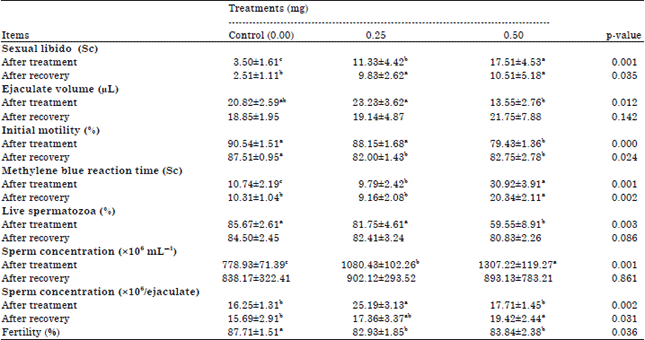 | |
| a,bMeans in any row with no common superscript differ (p≤0.05), values are given in Mean±SE | |
| Table 3: | Serum corticosterone and testosterone concentrations of male quails as affected by different levels of DEX treatments |
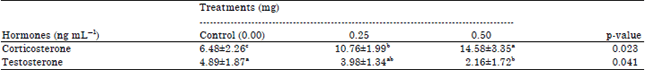 | |
| a,bMeans in any row with no common superscript differ (p≤0.05), values are given in Mean±SE | |
Moreover, serum corticosterone responses to DEX treatment were significantly (p≤0.05) induced in birds receiving the low and high DEX doses compared to control group (Table 3). Contrary with this result, testosterone decreased significantly (p≤0.05) in 0.50 mg administrated group but no significant difference was observed between 0.25 mg and control birds.
In our results, DEX administration influenced significantly the fertility percentage of male after one week of treatment ended (Table 2). The unstressed male quail has been a numerical higher percentage (p≤0.05) of fertilization in comparison to treated groups. This finding followed the same trend as the results of all the evaluations performed in the present study.
Correlation coefficients among studied traits: Results revealed that there were prevalent significant (p≤0.01) positive correlation between testosterone level and each of cloacal gland area, foam production, ejaculate volume, initial motility, live spermatozoa and fertility percentage. Also, between corticosterone level and each of TSL, MBRT, sperm concentration/mL. Same pattern was found between cloacal gland area and foam production, ejaculate volume, initial motility and live spermatozoa (Table 4). In contrast, significant negative correlation was found between concentration of testosterone and each of corticosterone level, TSL, MBRT and sperm concentration/mL. Also, between corticosterone level and cloacal gland area, foam production, initial motility (p≤0.05) and live spermatozoa. The same trend was observed between cloacal gland area and each of TSL (p≤0.05), MBRT (p≤0.05) and sperm concentration/mL (p≤0.01).
| Table 4: | Correlation coefficients among some studied traits |
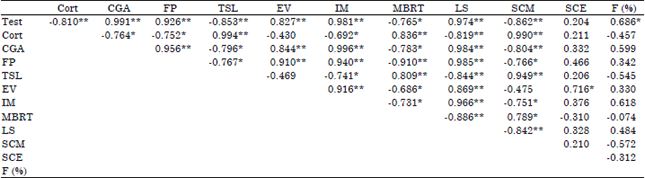 | |
| Test: Testosterone (ng mL–1), Cort: Corticosterone (ng mL–1), CGA: Cloacal gland area (mm2), FP: Foam production (mg), TSL: Time of sexual libido (Sc), EV: Ejaculate volume (µL), IM: Initial motility (%), MBRT: Methylene blue reaction time (Sc), LS: Live spermatozoa (%), SCM: Sperm concentration/mL (×106 mL–1), SCE: Sperm concentration/ejaculate (×106/ejaculate), F (%): Fertility (%), *: p≤0.05, **: p≤0.01 | |
DISCUSSION
Our study investigated the impact of DEX ingested for 14 days on reproductive parameters and semen traits in mature male Japanese quail. Also, we tested if the reproductive function of these birds can recover after oxidative stress condition induced via DEX administration.
The feed intake of mature quail fed DEX at 0.25 and 0.50 mg/bird/day was markedly lower than that of control. This result was in accordance with the report of Sapolsky et al. (2000) who found that DEX administration caused a decreased in feeding and appetite of chickens. Also, De La Cruz et al. (1981) recorded that food intake was less in adult quail given over 7 days corticosterone orally compared to control group. The effect of corticosterone on food intake in birds may be dose-dependent and hence, supports the notation that chronic stress can have deleterious consequences in birds. Corticosterone is generally considered to have a stimulatory effect on food intake and feeding behaviour in birds (Petitte and Etches, 1991; Kitaysky et al., 2003; Lohmus et al., 2006). On the other hand, treatment with corticosterone may decrease or have no effect on food intake in broiler chicks (Hayashi et al., 1994). However, food intake increased in chickens delivered higher doses of corticosterone for long period but not in chickens given lower doses for short time (Covasa and Forbes, 1995).
However, the body weight of mature quail received DEX at 0.25 or 0.50 mg/bird/day was not significantly different compare with control. These results are in agreement with the report of Hull et al. (2007) who found that the body weight of immature quail (3-5 weeks of age) which received DEX up to 0.94 mg day–1 in drinking water was unaffected despite continued treatment for two weeks. Conversely, Aengwanich (2007) and Li et al. (2009) reported that the body weight of broiler which received DEX up to 6 mg kg–1 in diets or injection into abdomen was significantly lower than control group and this decrease was consistence with levels of DEX increased. Their explanation is that DEX caused muscular dystrophy and reduced growth through restrained glucose absorption, increased plasma T3 levels and decreased protein metabolism. Although, the levels of DEX and studied birds were different in previous experiments, the status of growth period was also different between present study and others. However, this apparent DEX insensitivity may have been associated with the age of birds, older birds may simply be less sensitive to the administration effects of DEX (Davison et al., 1985; Breuner et al., 2008).
The current results showed that the growth of reproductive organs and testes were not uniformly reduced by two weeks of DEX treatment. While cloacal gland area and foam production were the most sensitive to DEX inhibition, testis was only slightly sensitive. Shit et al. (2010) reported that the direct relationship between cloacal gland and testicular activity in quail and added that the area of cloacal gland as a selection marker for sexually active male during breeding program. Also a high degree of correlation was established between the area of cloacal gland with testicular size (Follett and Maung, 1978), testosterone level (Mohan et al., 2002) and fertility (Biswas et al., 2007). Hence, it provides an external marker of the circulating androgen (Ball and Balthazart, 2010). However, after discontinuing DEX administration on day 14 of the experimental period it was found that both body weight and cloacal gland area of male quail recovered to the normal levels within 4 weeks.
It is widely accepted that, corticosterone directly inhibit GnRH and LH secretion (Etches et al., 1984) and testosterone synthesis (Buchanan et al., 2004; Van Hout et al., 2010) and this is thought to be the primary route for reproductive suppression during stressful events (Rivier and Rivest, 1991). For example, a previous study of adult male rufous-winged sparrows (Aimophilia carpalis) showed that, as corticosterone rises following handling stress, circulating testosterone levels are reduced by up to 50 percent via the direct action of corticosterone on testicular hormone production (Deviche et al., 2010). Similarly, our results have showed negative correlation between corticosterone levels and testosterone levels after chronic exposure to DEX. Consequently, a negative effect on sexual parameters and sperm quality would be expected. Induced stress by daily ingesting of DEX for 14 successive days caused a significant (p≤0.05) delayed in TSL and reduced in spermatozoa motility, viability, concentration and ejaculate value. This difference in inhibition on the libido of male and quality of semen coincide with the differences of testosterone concentrations between the DEX and non-DEX groups. Also, there are negative correlation between testosterone level and TSL and most semen quality traits. On the other hand, our results detected improvement of individual motility in 0.25 mg group and this enhance may be correlated with the increase of spermatozoa concentration and viability. These results are in agreement with those obtained with Taha (2011) who found a positive correlation between motility and concentration of spermatozoa in quail, due to the decrease in dead spermatozoa percentage. Thus, physiological changes inimical to testicular functions may not occur in short-term exposure to DEX. Interestingly, the 0.50 mg administrated had an enhancing effect on sperm concentration beyond those that had already been achieved by 0.25 mg, with a suppressing effect on sperm viability. We suppose the effects of DEX on testicular function are dose dependent and do not increase in a linear manner. The significant increase in spermatozoa concentration may be due to increase corticosterone level and decrease testosterone concentration after DEX treatments. However, after 4 weeks of recovery period the sperm concentrations as well as the libido of mature quail were consistently reduced by administrating DEX in this experiment. These findings confirm that DEX can affect male quail for up to several weeks after treatment has ended.
Oxidative stress has been shown to impair the proper functioning of Leydig cells leading to disturbed testes functions, particularly steroidogenesis and spermatogenesis, resulting in higher levels of apoptotic spermatozoa in the semen of rats and rabbits (Brecchia et al., 2010; Metukuri et al., 2010). Sperm of birds are highly susceptible to oxidative stress, which can negatively impact sperm function and ultimately male fertilizing efficiency (Losdat et al., 2011). Indeed, lipid content of the seminal plasma and spermatozoa make it a good target for free radical attacks under oxidative stress. This hypothesis was supported by Eid et al. (2006) who observed that the decline in sperm motility was associated with accumulation of thiobarbituric acid reactive substance in the blood and seminal plasma of male chicken administrated DEX for 7 successive days. By sampling males at the end and up to 28 days after the end of treatment, monitoring sperm quality over the entire spermatogenesis cycle, we might have been able to monitor the bulk of reduction in sperm quality due to DEX-induced oxidative stress. The percentage of motile sperm in the present study was significantly reduced (79.43%) by the chronic administration of 0.50 mg DEX. Hence, the possibility that germ cells suffered insults by local or systemic oxidative stress can not be discarded. Although semen quality was slightly improved after treatment ended, sperm motility may still have dramatic consequences for successful sperm-egg interactions and fertilization (Tremellen, 2008).
Average fertility results obtained in the present work after treatment ended varied from 82.93-87.71%. The fertility of eggs produced by females housed with treated male was lower 82.93 and 83.84% in 0.25 and 0.50, respectively, to the eggs of females housed with control male (87.71%). On the other hands, a numerical decrease has been observed in fertility percentage in the treated birds as compared to that of control with significant differences.
In summary, these data demonstrate that chronic DEX administration provides a non-invasive, effective method to investigate the effects of chronic stress in mature male Japanese quail. The effectiveness of the treatment was especially evident in the reduction in reproductive organ size and semen characteristics in treated males compared with untreated males. Also, libido is strongly impaired after oxidative stress, presumably through alteration in secretion of sex hormones. Importantly, DEX had marked effects on the reproductive performance of male quail for several weeks after treatment ended, suggesting that chronic elevations in plasma corticosterone resulting from climate change or human disturbance could have negative affects in birds even after exposure to a stressors ends.
REFERENCES
- Aengwanich, W. and O. Chinrasri, 2003. Effect of dexamethasone on differential white blood cell counts and heterophil/lymphocyte ratio in Japanese quails (Coturnix coturnix Japonica). Songklanakarin J. Sci. Technol., 25: 183-189.
Direct Link - Aengwanich, W., 2007. Effects of dexamethasone on physiological changes and productive performance in broilers. AJAVA., 2: 157-161.
CrossRefDirect Link - Ball, G.F. and J. Balthazart, 2010. Japanese quail as a model system for studying the neuroendocrine control of reproductive and social behaviors. ILAR J., 51: 310-325.
Direct Link - Biswas, A., J. Mohan, K.V.H. Sastry and J.S. Tyagi, 2007. Effect of dietary Vitamin E on the cloacal gland, foam and semen characteristics of male Japanese quail. Theriogenology, 67: 259-263.
CrossRefPubMedDirect Link - Brecchia, G., R. Cardinali, E. Mourvaki, G. Collodel, E. Moretti, A. Dal Bosco and C. Castellini, 2010. Short- and long-term effects of lipopolysaccharide-induced inflammation on rabbit sperm quality. Anim. Reprod. Sci., 118: 310-316.
CrossRefDirect Link - Breuner, C.W., S.H. Patterson and T.P. Hahn, 2008. In search of relationships between the acute adrenocortical response and fitness. Gen. Comp. Endocrinol., 157: 288-295.
CrossRefDirect Link - Buchanan, K.L., S. Leitner, K.A. Spencer, A.R. Goldsmith and C.K. Catchpole, 2004. Developmental stress selectively affects the song control nucleus HVC in the zebra finch. Proc. R. Soc. London B: Biol. Sci., 271: 2381-2386.
CrossRefDirect Link - Calandreau, L., A. Favreau-Peigne, A. Bertin, P. Constantin and C. Arnould et al., 2011. Higher inherent fearfulness potentiates the effects of chronic stress in the Japanese quail. Behav. Brain Res., 225: 505-510.
CrossRefDirect Link - Covasa, M. and J.M. Forbes, 1995. Selection of foods by broiler chickens following corticosterone administration. Br. Poult. Sci., 36: 489-501.
CrossRefDirect Link - Davison, T.F., B.M. Freeman and J. Rea, 1985. Effects of continuous treatment with synthetic ACTH1-24 or corticosterone on immature Gallus domesticus. Gen. Comp. Endocrinol., 59: 416-423.
CrossRefDirect Link - De La Cruz, L.F., F.J. Mataix and G. Illera, 1981. Effects of glucocorticoids on protein metabolism in laying quails (Coturnix coturnix Japonica). Compa. Biochem. Physiol. Part A: Physiol., 70: 649-652.
CrossRefDirect Link - Deviche, P.J., L.L. Hurley, H.B. Fokidis, B. Lerbour and B. Silverin et al., 2010. Acute stress rapidly decreases plasma testosterone in a free-ranging male songbird: Potential site of action and mechanism. Gen. Comp. Endocrinol., 169: 82-90.
CrossRefDirect Link - Eid, Y., T. Ebeid, M. Moawad and M. El-Habbak, 2008. Reduction of dexamethasone-induced oxidative stress and lipid peroxidation in laying hens by dietary vitamin E supplementation. Emir. J. Food Agric., 20: 28-40.
Direct Link - Eid, Y., T. Ebeid and H. Younis, 2006. Vitamin E supplementation reduces dexamethasone-induced oxidative stress in chicken semen. Br. Poult. Sci., 47: 350-356.
CrossRefDirect Link - Etches, R.J., J.B. Williams and J. Rzasa, 1984. Effects of corticosterone and dietary changes in the hen on ovarian function, plasma LH and steroids and the response to exogenous LH-RH. J. Reprod. Fertil., 70: 121-130.
CrossRefDirect Link - Follett, B.K. and S.L. Maung, 1978. Rate of testicular maturation, in relation to gonadotrophin and testosterone levels, in quail exposed to various artificial photoperiods and to natural day lengths. J. Endocrinol., 78: 267-280.
CrossRefPubMedDirect Link - Gao, J., H. Lin, X.J. Wang, Z.G. Song and H.C. Jiao, 2010. Vitamin E supplementation alleviates the oxidative stress induced by dexamethasone treatment and improves meat quality in broiler chickens. Poult. Sci., 89: 318-327.
CrossRefDirect Link - Golzar Adabi, S.H., S.H. Rahimi, M.A. Karimi and M.A. Kamali, 2007. The effects of two dietary levels of L-carnitine and vegetable fat powder on the quality of cockerels' sperm and fertility and hatchability in broiler breeders. J. Vet. Res., 3: 107-114.
Direct Link - Hayashi, K., Y. Nagai, A. Ohtsuka and Y. Tomita, 1994. Effects of dietary corticosterone and trilostane on growth and skeletal muscle protein turnover in broiler cockerels. Br. Poult. Sci., 35: 789-798.
CrossRefDirect Link - Hull, K.L., J.F. Cockrem, J.P. Bridges, E.J. Candy and C.M. Davidson, 2007. Effects of corticosterone treatment on growth, development, and the corticosterone response to handling in young Japanese quail (Coturnix coturnix Japonica). Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 148: 531-543.
CrossRefDirect Link - Kitaysky, A.S., E.V. Kitaiskaia, J.F. Piatt and J.C. Wingfield, 2003. Benefits and costs of increased levels of corticosterone in seabird chicks. Hormone Behav., 43: 140-149.
CrossRefDirect Link - Li, Y., H.Y. Cai, G.H. Liu, X.L. Dong and W.H. Chang et al., 2009. Effects of stress simulated by dexamethasone on jejunal glucose transport in broilers. Poult. Sci., 88: 330-337.
CrossRefDirect Link - Lin, H., E. Decuypere and J. Buyse, 2004. Oxidative stress induced by corticosterone administration in broiler chickens (Gallus gallus domesticus): 1. Chronic exposure. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 139: 737-744.
CrossRefDirect Link - Lohmus, M., L.F. Sundstrom and F.R. Moore, 2006. Non-invasive corticosterone treatment changes foraging intensity in red-eyed vireos Vireo olivaceus. J. Avian Biol., 37: 523-526.
CrossRefDirect Link - Losdat, S., H. Richner, J.D. Blount and F. Helfenstein, 2011. Immune activation reduces sperm quality in the great tit. PLoS One, Vol. 6.
CrossRefDirect Link - Metukuri, M.R., C.M.T. Reddy, P.R.K. Reddy and P. Reddanna, 2010. Bacterial LPS mediated acute inflammation-induced spermatogenic failure in rats: Role of stress response proteins and mitochondrial dysfunction. Inflammation, 33: 235-243.
CrossRefDirect Link - Mohan, J., R.B. Moudgal, K.V.H. Sastry, J. Tyagi and R. Singh, 2002. Effects of hemicastration and castration on foam production and its relationship with fertility in male Japanese quail. Theriogenology, 58: 29-39.
CrossRefDirect Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA., ISBN-13: 9780309048927, Pages: 176.
Direct Link - Petitte, J.N. and R.J. Etches, 1991. Daily infusion of corticosterone and reproductive function in the domestic hen (Gallus domesticus). Gen. Comp. Endocrinol., 83: 397-405.
CrossRefDirect Link - Rivier, C. and S. Rivest, 1991. Effect of stress on the activity of the hypothalamic-pituitary-gonadal axis: Peripheral and central mechanisms. Biol. Reprod., 45: 523-532.
CrossRefDirect Link - Romero, L.M. and J.M. Reed, 2008. Repeatability of baseline corticosterone concentrations. Gen. Comp. Endocrinol., 156: 27-33.
CrossRefDirect Link - Sapolsky, R.M., L.M. Romero and A.U. Munck, 2000. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory and preparative actions. Endocrinol. Rev., 21: 55-89.
CrossRefDirect Link - Scheuerlein, A., T. Van't Hof and E. Gwinner, 2001. Predators as stressors? Physiological and reproductive consequences of predation risk in tropical stonechats (Saxicola torquata Axillaris). Proc. Biol. Sci., 268: 1575-1582.
CrossRefDirect Link - Schoech, S.J., M.A. Rensel, E.S. Bridge, R.K. Boughton and T.E. Wilcoxen, 2009. Environment, glucocorticoids and the timing of reproduction. Gen. Comp. Endocrinol., 163: 201-207.
CrossRefDirect Link - Shit, N., R.P. Singh, K.V.H. Sastry, J. Mohan, N. Pandey and R.P. Moudgal, 2010. Cloacal gland size significantly alters semen production, sperm activities and fertility in different lines of Japanese quail (Coturnix coturnix Japonica). Asian J. Poult. Sci., 4: 190-197.
CrossRefDirect Link - Taha, A.T., 2011. Effect of dietary supplementation with different levels of fenugreek seeds (Trigonella foenum Graecum) on semen quality and histological testis traits of Japanese quail (Coturnix coturnix Japonica). Diyala Agric. Sci. J., 3: 1-9.
Direct Link - Tasker, J.G. and J.P. Herman, 2011. Mechanisms of rapid glucocorticoid feedback inhibition of the hypothalamic-pituitary-adrenal axis. Stress, 14: 398-406.
Direct Link - Tremellen, K., 2008. Oxidative stress and male infertility-a clinical perspective. Hum. Reprod. Update, 14: 243-258.
CrossRefDirect Link - Westerhof, I., W.E. Van den Brom and J.A. Mol, 1996. Responsiveness of the glucocorticoid-suppressed pituitary-adrenocortical system of pigeons (Columba livia domestica) to stimulation with arginine vasopressin. Avian Dis., 40: 312-320.
CrossRefDirect Link - Kitaysky, A.S., E.V. Kitaiskaia, J.F. Piatt and J.C. Wingfield, 2003. Benefits and costs of increased levels of corticosterone in seabird chicks. Hormone Behav., 43: 140-149.
CrossRefDirect Link