
D. A. Komeil
Department of Plant Pathology, Faculty of Agriculture, Alexandria University, Alexandria, Egypt
LiveDNA: 20.34451
M. Y. Abdalla
Faculty of Desert and Environmental Agriculture, Matrouh University, Egypt
A. F. El-Bebany
Department of Plant Pathology, Faculty of Agriculture, Alexandria University, Alexandria, Egypt
A. B. A. Basyony
Department of Plant Pathology, Faculty of Agriculture, Alexandria University, Alexandria, Egypt
Asian Journal of Plant Sciences
Year: 2021 | Volume: 20 | Issue: 2 | Page No.: 363-369







ABSTRACT
Background and Objective: Date palm (Phoenix dactylifera L.) is an economically important food crop in Egypt. In addition to its edible sweet fruits, it contributes in the preservation of the arid ecosystems threatened by desertification. In the current study, an isolate of Fusarium was isolated from a wilt date palm orchard in Aswan (Upper Egypt). Materials and Methods: In the present study, wilting of some orchards of date palm in Aswan (Upper Egypt), were studied. Samples were cultured in common media (PDA) and Fusarium strain was morphologically and molecularly characterized. The pathogenicity test on date palm seedlings were carried out on ten cultivars from different regions of Middle East and North Africa by stem inoculation method. A mitochondrial molecular marker to test date palm germplasm for resistance/ susceptibility to the Bayoud disease was applied in using PCR based on the two primers oli1 and oli2. Results: Results for pathogenicity test indicated that the ten studied cultivars are susceptible to Fusarium oxysporum Aswan and they differed genetically as evidenced by analysis of molecular variance using mitochondrial resistance markers to Bayoud disease. In addition, the results indicated that all germplasms of Egyptian cultivars had susceptibility marker and there is an association occurred between geographical distances of date palm populations studied. Conclusion: Screening of date palm cultivars for resistance to Bayoud disease showed that there is a genetic variation between Egyptian cultivars and other tested cultivars.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
D. A. Komeil, M. Y. Abdalla, A. F. El-Bebany and A. B. A. Basyony, 2021. Resistance Marker Detection in Ten Date Palm Cultivars to the Wilt Pathogen, Fusarium oxysporum. Asian Journal of Plant Sciences, 20: 363-369.
DOI: 10.3923/ajps.2021.363.369
URL: https://scialert.net/abstract/?doi=ajps.2021.363.369
DOI: 10.3923/ajps.2021.363.369
URL: https://scialert.net/abstract/?doi=ajps.2021.363.369
INTRODUCTION
Agrobiodiversity refers to the variability of plants, animals and microbial organisms in a cultivated area. Conservation of agrobiodiversity is central to the continuity of agricultural production systems. Date palm (Phoenix dactylifera L.), a member of the palm family Arecaceae, has been cultivated for millennia in the Middle East and North Africa1. Its time of domestication, routs of germplasm flows and the breeding history in various cultivation areas are only partially known2. Genomic data have pointed out that there are two gene pools of date palm, one in the Middle East and the second in Africa3,4. The strategic location of Egypt, being partially in North Africa and partially in South East Asia (Sinai Peninsula) may enrich the genetic pool of the various date palm varieties grown in it. Moreover, Egypt is the world largest producer of dates with annual production of 1.590 million t, equivalent to 19.7% of world production totaling 8.1 million t5.
Similar to other crops, date palm trees are attacked by various diseases that may reduce their productivity or even lead to their death. One of the most known diseases of date palm is Fusarium wilt. Several Fusarium spp., such as F. oxysporum, F. proliferatum, F. solani, F. moniliforme, F. equiseti and F. semitectum were reported to cause wilt and root rot of date palm6-8. However, F. oxysporum was the most predominant fungus isolated from most of the Egyptian governorates that they covered in their study9.
Based on the scientific and economic importance, F. oxysporum ranked fifth on the list of top ten fungal pathogens worldwide10,11. Generally, F. oxysporum strains attacking the same host plant are considered to belong to the same Forma specialis. Recently, Edel-Hermann and Lecomte12, stated that they counted 106 forma specialis for F. oxysporum, whereas, Lombard13 listed 144 different special forms for it. The proper identification of these special forms is of great importance to plant pathologists and plant breeders.
Date palm trees are subject to attack by more than one forma specialis of F. oxysporum. The most destructive one is F. oxysporum f. sp. albedinis (FOA), the causal agent of the Bayoud disease. The spread of the disease led to the extinction of several valuable varieties in the region. It was responsible for the eradication of more than 15 million date palm trees in Morocco and Algeria14. Nonetheless, in Mauritania, it destroyed several thousands of date palm trees15. Due to the success of the phytosanitary rules, Egypt and other Middle Eastern countries remained free of F. oxysporum f.sp. albedinis. Meanwhile, in the United Arab Emirates, there is another forma specialis of F. oxysporum associated with date palm wilt namely, F. oxysporum f. sp. cumini16.
Recently, farmers in upper Egypt, particularly in Aswan governorate, have been complaining about severe losses in their date palm orchards especially with more valuable varieties such as, Sakkoty, Bartmoda and Malakaby. The variety Malakaby has almost been extinct due to Fusarium wilt disease. The present study aimed at investigating the Fusarium wilt disease of date palm in Aswan governorate in upper Egypt, in order to determine whether it belongs to F. oxysporum f. sp. albedinis or not. To accomplish this goal, the reaction of certain germplasms of date palm from various origins to infection with the Aswan isolate of F. oxysporum (AFO) was studied. This was done to detect the mitochondrial molecular marker of resistance/susceptibility to Bayoud disease in the same ten germplasms from various origins and to correlate the molecular marker detection to the disease severity index observed through the variety reaction in the pathogenicity test.
MATERIALS AND METHODS
Study area: Pot experiment was carried out at the experimental green house of Plant Pathology Department, Alexandria University, Egypt, during the two winter seasons of 2017/2018 and 2018/2019.
Pathogen isolation, purification and preservation: Leaf rachis of naturally infected date palm trees (cv. Sakkoty, grown at Edfo city, Aswan governorate, Upper Egypt) was collected. At the laboratory, the leaves rachises were prepared by removing thorns and leaflets and then the midribs were cut into pieces of 10-15 cm and were washed under running tap water for 5 min to remove dust and soil particles. Leaves' rachises segments were further cut into smaller pieces (1-2 cm), then soaked for 2 min in 0.5% sodium hypochlorite solution (NaOCl), rinsed in sterile distilled water (SDW) and dried on sterilized filter papers. The surface disinfested leaves rachises sections were placed on acidified potato dextrose agar (APDA) with pH = 5 and incubated at 25±2°C for 2 weeks. Emerged fungal growth was purified using the single spore technique, transferred to potato dextrose agar (PDA) medium. Identification was performed according to the taxonomic system of Leslie and Summerell17. Stock cultures were maintained on PDA slants and refrigerated at 5°C.
Macroscopic and microscopic identification: The fungal morphology was studied macroscopically by observing the colony features (color, shape, size and hyphal morphology) and microscopically by a compound microscope using a lacto phenol cotton blue stained slide mounted with a small portion of the mycelium with the help of keys as described by Leslie and Summerell17, where asexual conidial spores were observed.
Molecular identification using ITS-PCR amplification: Extraction of genomic DNA from the Fusarium isolate from Aswan was conducted from a one-week-old PDA culture. Total genomic DNA was isolated from fresh mycelium according to the manufacturer’s instructions (i-genomic BYF DNA extraction mini kit-INTRON Biotechnology) and quantified using a spectrophotometer (MaestroNano Drop MN-913). The Internal Transcribed Spacer (ITS) region of ribosomal DNA (rDNA) of the isolated fungus was amplified using primer set ITS1 (5'-TCCGTAGGTGAACCTGCGG-3') and ITS4 (5'-TCCTCCGCTTATTGATATGC-3')18. PCR amplification was done in a 25 μL volume containing 12.5 μL PCR mix (OnePCR Ultra, GeneDirex company), 1 μL of each ITS1 and ITS4 primers. Negative controls containing the same reagents, but without DNA template, were included in all PCR reaction sets. Amplification reactions were done in a thermo cycler (Techne- Progene, UK) with an initial denaturation at 94°C for 5 min, 35 cycles of denaturation at 94°C for 40 sec, annealing at 58°C for 40 sec, extension at 72°C for 1 min, and a final extension at 72°C for 10 min. Amplification products were visualized by 1.5% polyacrylamide gel electrophoresis combined with RedSafeTM Nucleic Acid Staining Solution. The resulting amplicons of approximately 700 bp were intended for sequencing.
According to EPPO diagnostic protocol (PM7/16(1), the Fusarium isolate from Aswan was verified using the TLO3-FOA28 specific primers of F. oxysporum f.sp. albedinis19. The conventional PCR was carried out at Molecular Laboratory at Mycology Research and Disease Survey.
Sequencing and analysis: The PCR products were sent for sequencing through real biogene biotechnology services company, Alexandria, Egypt to AGCT-Advanced Genoming Computing Technologies-Germany. Sequence similarity BLAST searches were performed to compare the ITS sequence with other known related sequences using the nucleotide blast program at http://blast.ncbi.nlm.nih.gov/Blast.cgi. The sequence of the pathogenic isolate was submitted to Genbank with accession number MT337537.
Pathogenicity tests: The Fusarium isolate from Aswan was grown on APDA for 7-14 days at 25°C. The inoculum was prepared by flooding the agar surface with 10 mL of SDW, gently scraping the colony surface with a sterile spatula. The resulting spore suspension was filtered through three layers of cheese cloth. Spore suspension was adjusted to 106 conidia mL–1 using a haemocytometer.
Date fruits of the ten studied varieties, Sakkoty, Bartmoda and Siwi from Egypt, Barhi, Succary, Saguae and Ajwa from Saudi Arabia, Deglet Noor from Algeria and Deglet Noor from Tunisia and finally, Fardh from Oman were obtained from commercial producers at each country. Seeds were extracted then thoroughly were washed under tap water, surface sterilized for 5 min in 0.5% NaOCl. Seeds of each variety were then soaked in sterilized tap water for 24 hrs and then planted in seedling bags filled with sterilized mixture of equal portions (v/v) of sand and peat moss. One seedling emerged in each pot. Ten pots were used for each of the tested varieties, consequently 100 seedlings (pots) were used in the experimental run. Seedlings were allowed to grow for 6 months in the greenhouse at 25±3°C before inoculation. Five seedlings of each variety were inoculated with the fungal spores’ suspension as described by Abdalla et al.6. Each seedling was injected with 2 mL of conidial spore suspension into the crown area using a hypodermic needle and a syringe, whereas control seedlings were injected similarly but with SDW. After inoculation, all plants were watered and covered separately with dark plastic bags for 48 hrs to maintain high humidity.
Each seedling was rated on a scale from 0-4, where 0 = no visible symptoms, 1 = small lesion (5-10 mm), 2 = medium lesion (10-30 mm), 3 = large lesion (30-70 mm) and 4 = whole leaf blight. Three months after inoculation, obtained data were used to determine the disease severity index (DSI) as proposed by Abdalla et al.6.
Where:
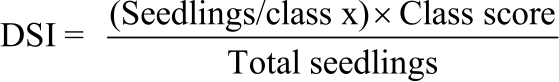 |
This experiment was repeated twice and data were combined. Obtained data were analyzed according to general linear models procedures of Statistical Analysis System (SAS version 9.3, SAS Institute Inc., Carry, NC.) The causal agent was re-isolated from the artificially inoculated seedlings and identity was confirmed by morphological and molecular characterization in order to fulfill Koch's postulates.
Detection of resistance/susceptibility of date palm varieties using molecular markers: The PCR conditions of Bayoud resistibility/susceptibility amplification reaction and the two primers: oli1 (5'-CCTTATACCAGTCGTGCTT-3') and oli2 (5'-AAGGCAGATATAATCGGA-3') were chosen according to Saleh et al.20. To detect the same molecular markers in the tested date palm varieties, PCRs were performed using 20 ng of gDNA in a total volume of 50 μL. Briefly, PCR reaction were incubated in a thermo cycler (Techne-Progene, UK) starting with 5 min of denaturation at 95°C for 5 min, followed by 35 cycles at 95°C for 30 sec and 56°C as annealing temperature for 60 sec and 72°C for 1 min and a terminal incubation at 72°C for 5 min. To verify the size and presence of the expected fragments, the amplification products were separated on a 1% agarose gel in Tris-acetic acid-disodium EDTA buffer containing RedSafeTM Nucleic Acid Staining Solution (iNtron Biotechnology) and visualized using UV transilluminator.
RESULTS AND DISCUSSION
Identification of the pathogenic fungus: Isolation from different leaves rachises tissues from various wilting date palm trees in Edfo city, Aswan governorate, Upper Egypt, revealed the presence of Fusarium oxysporum Schlecht. Emend. Snyder and Hans. According to morphological traits on APDA medium, colonies were cottony with white aerial mycelium and produced purple color into the medium. Conidiophores were simple or branched, microconidia were oval or kidney-shaped and 1-2 celled varying in size (3-5×7-12 μm). Macroconidia were less abundant, had 3-5 septa, boat-shaped, measuring 3-5×25-32 μm. Chlamydospores developed in older cultures were, generally, solitary but occasionally in pairs or short chains. Chlamydospores were smooth to rough walled, with 7-12 μm diameters. Based on sequence analysis of the ITS region of the ribosomal DNA (rDNA), the Aswan isolate was confirmed to be F. oxysporum MT337537. According to molecular verification test for F. oxysporum f. sp. albedinis21, it was proved that the Aswan isolate of F. oxysporum did not belong to the forma speciales albedinis.
Pathogenicity tests and variety reaction: All tested varieties of date palm were susceptible to infection with the Aswan isolate of F. oxysporum, however, various degrees of susceptibility were observed (Table 1). Varieties Sakkoty and Barhi were the most susceptible with DSI = 4.4. Followed by cv. Fardh with DSI = 3.5. Whereas, varieties Bartmoda and Agwa EL-Madina had DSI of 3.3, 3.0. Meanwhile, varieties Siwa, Deglet Noor from Algeria and Tunisia and Sagae were moderately resistant with DSI of 2.2, 2.5, 2.0 and 1.9, respectively. Finally, cv. Succary was the least susceptible variety with DSI of only 1.5.
 |
| Fig. 1: | PCR amplification products of the resistance/ susceptibility molecular markers generated from the ten tested date palm cultivars using oli1 and oli2 primers Date palm cultivars 1= Bartameda; 2 = Sakkoty; 3 = Siwi; 4 = Barhi; 5 = Succary; 6 = Seguae; 7 = Ajwa ; 8 = Deglet Nour; 9 = Fardh; 10 = Degelt Noor) M, DNA molecular size marker |
| Table 1: Disease severity index of the ten tested date palm cultivars | ||
| Variety | Geographic origin | *Disease severity index |
| Bartameda | Egypt | **3.3ab |
| Sakkoty | " | 4.0a |
| Siwi | " | 2.2cd |
| Barhi | Saudi Arabia | 4.0a |
| Succary | " | 1.5d |
| Seguae | " | 1.9d |
| Ajwa | " | 3.0abc |
| Deglet Nour | Tunisia | 2.0cd |
| Fardh | Oman | 3.5ab |
| Deglet Nour | Algeria | 2.5bcd |
| *Disease severity was calculated based the scale according to Abdalla et al.6 **Data are means of five replicates. Data with the same letter/s, in each column for each cultivar are not significantly different at p<0.05 | ||
Molecular characterization of tested date palm germplasm using resistance/susceptibility markers: The PCR approach, using two specific primers oli1 and oli2, was used to screen a collection of Bayoud-susceptible and Bayoud-resistant date palm varieties with respect to the identity of the plasmid structure (S or R) which they contained (Fig. 1). The PCR reaction based on the two specific primers oli1 and oli2 produced two PCR amplicons from the mitochondrial S-plasmid and R-plasmid20. Lanes from 1-10 presenting PCR amplicons from the mitochondrial S-plasmid and R-plasmid of the ten examined date palm varieties. The obtained results showed that the tested date palm germplasms were divided into two groups. The first group is characterized by a susceptibility marker with size of approximately 357 bp in lanes 1, 2 and 3, respectively. The presence of S-plasmid, contained only Egyptian varieties Bartameda, Sakkoty and Siwa. Whereas the second group is associated to the R-plasmid presumably associated with resistance to Bayoud disease. The resistance marker with molecular size of approximately 264 bp was detected with the remaining seven date palm cultivars, namely, Barhi, Succary, Seguae, Ajwa, Fardh and the two varieties of Deglet Nour. Variety Deglet Nour whether from Algeria or from Tunisia had the R-plasmid. Both varieties showed moderate susceptibility to AFO (DSI = 2.5 and 2.0). The cv. Succary from Saudi Arabia had the R- plasmid which was in agreement with the observations reported by Saleh et al.20. This economically important variety was actually the most resistant to infection with AFO. The mitochondrial molecular markers recommended b y Saleh et al.20 to test date palm germplasm for resistance/ susceptibility to Bayoud disease were utilized in this study.
Date palm is one of the main fruit crops in the arid and semiarid regions of Western Asia and North Africa. It is socio-economically and traditionally important for local populations where the culture thrives21,22. It constitutes the cornerstone of oasis agro systems in these regions. Dispersal of date palm germplasm was probably originally by seed, which gave rise to the many local cultivars that are found in the Middle East23. Depending on local soil and climate conditions, local farmers have selected various varieties to meet their needs. Currently, there are about 5000 date palm varieties distributed worldwide with different fruit color, size, shape and ripening time2. In Egypt, 52 date palm varieties are found distributed in the Nile Valley, Oasis and various desert districts9 as Egypt holds the first place among the top date producing countries worldwide.
Date palm is playing an important role in Egyptian agriculture as it represents economic wealth. It is the source of a wide range of products and services, including many necessities of life. The primary product of the date palm is fruit, which is rich in protein, vitamins and mineral salts. All secondary products of the palm resulting from annual pruning have essential uses for the cultivator. According to what was presented, palm orchards must be preserved from destructive diseases such as Bayoud disease. The destructive FOA causes date palm wilt in Algeria, Morocco and Mauritania21, was believed to be descended from a common evolutionary ancestor of F. oxysporum19. Fusarium wilt disease progression is slow and is usually spread by transport of infected plant materials24. Till now, there are five Fusarium wilt diseases infecting date palms, each with fatal consequences.
There are few reports about the declined date palm trees in Egypt, that is associated with three Fusarium spp., F. solani and F. moniliforme and F. oxysporum9,25. It was evident from the current study that decline disease found in date palm growing areas of Aswan is caused by F. oxysporum MT337537. Obtained results are in agreement with those reported by Bekheet et al.9, who found that F. oxysporum was the predominant fungus isolated from most of the Egyptian governorates and the most virulent fungus causing root rot and wilt of date palm. It was important to determine whether F. oxysporum MT337537 is not FOA, that is considered as the main problem of date palm in North Africa14,15. Date palm groves create, in this location, a favorable condition to Egyptian farmers to cultivate other crops in between date palm trees such as vegetables9,25. Some of these crops are carriers of the pathogen which may lead to a significant damage to the socio-economic status of date palm industry in this region. The fast crop rotation of vegetables may allow the pathogen to adapt for pathogenicity factors that help overcoming resistance of several crops which means high rate of new forma specials development.
Pathogenicity of F. oxysporum has only been studied on molecular basis on limited number of hosts e.g., melon, tomato, bean, chickpea and Arabidopsis thaliana26. Thus, pathogenicity of the various forma speciales of F. oxysporum on date palm has only drowned little attention of scientists especially on molecular basis. Moreover, since the degree of resistance to infection with F. oxysporum is determined by many factors, each of these factors has only limited effect27. Studies of pathogenicity of F. oxysporum on Arabidopsis sp. revealed the existence of wide variation between the levels of susceptibility among various ecotypes27. This was due to the variation of resistance mechanisms involved in limiting the infection with F. oxysporum.
These observations were, in the present study, manifested the reaction of the three tested Egyptian varieties to infection with AFO. The two varieties, Sakkoty and Partmoda grown in Aswan reacted differently to infection with AFO as compared with the Siwa variety which is grown in the Siwa oasis, located more than one thousand six hundred kilometers apart from Aswan. Whereas, variety Deglet Noor whether from Algeria or Tunisia had the R- plasmid which is in agreement with the results reported by Quenzar et al.28 and Saleh et al.20.
Since no cure exists for the Bayoud disease, genetic control remains the only solution for this problem14. Therefore, research work has aimed at developing resistance reliable markers within date palm populations existing in contaminated zones28. These efforts have resulted in finding two minicircular DNA's in date palm mitochondria: The R-plasmid (1345 bp) found in the mitochondria of Bayoud-resistant varieties and the S-plasmid (454 bp) found in date palm varieties sensitive to Bayoud. Salem et al.29 reported that strong significant correlation existed between the hosted plasmid type and resistance of a certain variety against Bayoud. In the present study, it appeared that all tested Egyptian varieties contained the S plasmid (Fig. 1). This may imply the vulnerability of the tested Egyptian varieties to the Bayoud disease. The genetic diversity is very essential for the control of certain diseases such as Bayoud. In this case, extreme quarantine precautions should be implied at the Egyptian ports in order to prevent the introduction of such devastating disease into Egypt.
More research is needed in order to obtain more information about pathogenicity of the various strains of F. oxysporum on date palm as well as the molecular basis of resistance reaction in the various varieties. This knowledge would be great asset for early prediction of disease spread, promoting programs for disease resistance and improving strategies for disease control. Combining data from classical and molecular tools will prevent losing efforts, time and costs in date palm agricultural systems.
CONCLUSION
Screening of ten Arabian date palm cultivars for resistance to Bayoud disease using mitochondrial resistance markers showed that there is a genetic variation between Egyptian cultivars and other tested cultivars. This information could be helpful for sustained preservation of the germplasm to avoid considerable damage of Bayoud.
SIGNIFICANCE STATEMENT
This study discovered that the decline disease found in date palm growing areas of Aswan is caused by F. oxysporum and this strain is not related to F. oxysporum f.sp. albedinis. This fact can be beneficial to sustained preservation of the germplasm to avoid considerable damage of Bayoud. This study will help the researcher to uncover the critical areas of genetic diversity of resistance of the different palm date that many researchers were not able to explore. Thus, a new theory on resistant/susceptibility of date palm to Bayoud disease may be arrived at.
REFERENCES
- Barrow, S.C., 1998. A monograph of Phoenix L. (Palmae:Coryphoideae). Kew Bul., 53: 513-575.
Direct Link - Pintaud, J.C., B. Ludeña, F. Aberlenc-Bertossi, S. Zehdi and M. Gros-Balthazard et al., 2013. Biogeography of the date palm (Phoenix dactylifera L., Arecaceae): Insights on the origin and on the structure of modern diversity. Acta Hortic., 994: 19-38.
CrossRefDirect Link - Hazzouri, K.M., J.M. Flowers, H.J. Visser, H.S. Khierallah and U. Rosas et al., 2015. Whole genome re-sequencing of date palms yields insights into diversification of a fruit tree crop. Nat. Commun., Vol. 6.
Direct Link - Gros-Balthazard, M., M. Galimberti, A. Kousathanas, C. Newton and S. Ivorra et al., 2017. The discovery of wild date palms in Oman reveals a complex domestication history involving centers in the Middle East and Africa. Curr. Biol., 27: 2211-2218.
CrossRefDirect Link - Ahmed, M.F.A., 2018. Management of date palm root rot diseases by using some biological control agents under organic farming system. Novel Res. Microbiol. J., 2: 37-47.
CrossRefDirect Link - Abdalla, M.Y., A. Al-Rokibah, A. Moretti and G. Mule, 2000. Pathogenicity of toxigenic Fusarium proliferatum from date palm in Saudi Arabia. Plant Dis., 84: 321-324.
Direct Link - El-Morsi, M.E.A., M.A.M. Kamhawy and M.A.A. Sallam, 2009. Effectiveness of some organic compounds in controlling pathogenic fungi associated with roots of date palm offshoots in new valley governorate, Egypt. Assiut J. Agric. Sci., 40: 137-150.
Direct Link - Saleh, A.A., A.H. Sharafaddin, M.H. El Komy, Y.A. Ibrahim, Y.K. Hamad and Y.Y. Molan, 2017. Fusarium species associated with date palm in Saudi Arabia. Eur. J. Plant Pathol., 148: 367-377.
CrossRefDirect Link - Bekheet, S.A. and S.F. El-Shrabasy, 2015. Date Palm States and Perspective in Egypt. In: Date Palm Genetic Resources and Utilization, Al-Kayri, J.M., S.M. Jain and D.V. Johnson (Eds.)., Springer, Netherlands, pp: 75-123.
CrossRefDirect Link - Geiser, D.M., M. del Mar Jimenez-Gasco, S.C. Kang, I. Makalowska and N. Veeraraghavan et al., 2004. FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol., 110: 473-479.
CrossRefDirect Link - Dean, R., J.A.L. van Kan, Z.A. Pretorius, K.E. Hammond-Kosack and A. Di Pietro et al., 2012. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol., 13: 414-430.
CrossRefDirect Link - Edel-Hermann, V. and C. Lecomte, 2019. Current states of Fusarium oxysporum formae specials and races. Phytopathology, 109: 512-530.
CrossRefDirect Link - Lombard, L., M. Sandoval-Denis, S.C. Lamprecht and P.W. Crous, 2019. Epitypification of Fusarium oxysporum–clearing the taxonomic chaos. Persoonia, 43: 1-47.
CrossRefDirect Link - Benzohra, I.E., M. Megateli and R. Berdja, 2015. Bayoud disease of date palm: History, epidemiology and integrated disease management. Afr. J. Biotechnol., 14: 542-550.
CrossRefDirect Link - Sedra, M.H., 2013. The bayoud (vascular wilt) of date palm in North Africa: Situation, research achievements and applications. Acta Hortic., 994: 59-75.
CrossRefDirect Link - Alwahshi, K.J., E.E. Saeed, A. Sham, A.A. Alblooshi and M.M. Alblooshi et al., 2019. Molecular identification and disease management of date palm sudden decline syndrome in the united Arab emirates. Int. J. Mol. Sci., Vol. 20.
CrossRefDirect Link - Leslie, J.F. and B.A. Summerell, 2006. The Fusarium Laboratory Manual. 1st Edn., Blackwell Publishing, Iowa, USA., ISBN: 0-8138-1919-9.
Direct Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Fernandez, D., M. Ouinten, A. Tantaoui, J.P. Geiger, M.J. Daboussi and T. Langin, 1998. Fot 1 insertions in the Fusarium oxysporum f. sp. Albedinis genome provide diagnostic PCR targets for detection of the date palm pathogen. Applied Environ. Microbiol., 64: 633-636.
PubMed - Saleh, A.A., M.H. El Komy, A. Eranthodi, A.S. Hamoud and Y.Y. Molan, 2015. Variation in a molecular marker for resistance of Saudi date palm germplasm to Fusarium oxysporum f. sp. albedinis the causal agent of Bayoud disease. Eur. J. Plant Pathol., 143: 507-514.
CrossRefDirect Link - EPPO (European and Mediterranean Plant Protection Organization), 2003. Fusarium oxysporum f. sp. albedinis. EPPO Bull., 33: 265-269.
CrossRefDirect Link - Jain, S.M., J.M. Al-Khayri and D.V. Johnson, 2011. Date Palm Biotechnology. Springer, Netherlands., ISBN: 978-94-007-1318-5, Pages: 743.
CrossRefDirect Link - Krueger, R.R., 2011. Date Palm Germplasm. In: Date Palm Biotechnology, Jain, S.M., J.M. Al-Khayri and D.V. Johnson (Eds.)., Springer, Netherlands, pp: 313–335.
CrossRefDirect Link - Jaradat, A.A. and A. Zaid, 2004. Quality traits of date palm fruits in a center of origin and center of diversity. Food Agric. Environ., 2: 208-217.
Direct Link - El-Deeb, H.M., S.M. Lashin and Y.A. Arab, 2007. Distribution and pathogenesis of date palm fungi in Egypt. Acta Hortic., 736: 421-429.
Direct Link - Michielse, C. and M. Rep, 2009. Pathogen profile update: Fusarium oxysporum. Molecular Plant Pathol., 10: 311-324.
CrossRefDirect Link - Czymmek, K.J., M. Fogg, D.H. Powell, J. Sweigard, S.Y. Park and S. Kang, 2007. In vivo time-lapse documentation using confocal and multi-photon microscopy reveals the mechanisms of invasion into the Arabidopsis root vascular system by Fusarium oxysporum. Fungal Genet. Biol., 44: 1011-1023.
CrossRefDirect Link - Quenzar, B., M. Trifi, B. Bouachrine, C. Hartmann, M. Marrakchi, A.A. Benslimane and A. Rode, 2001. A mitochondrial molecular marker of resistance to Bayoud disease in date palm. Theor. Appl. Genet., 103: 366-370.
CrossRefDirect Link - Salem, A., S. Rhouma, S. Zehdi, M. Marraakchi and M. Trifi, 2007. Molecular characterization of Mauritanian date palm cultivars using plasmid-like DNAs markers. Biol. Plant., 51: 169-172.
CrossRefDirect Link