
Neelam Sherwani
Department of Biology, College of Science, Sultan Qaboos University, P.O. Box 36, Al Khoud, PC123, Oman
Sardar Abulfazal Farooq
Department of Biology, College of Science, Sultan Qaboos University, P.O. Box 36, Al Khoud, PC123, Oman
LiveDNA: 91.24000
Asian Journal of Plant Sciences
Year: 2019 | Volume: 18 | Issue: 2 | Page No.: 75-84







ABSTRACT
Background and Objective: Dipterygium glaucum, a rangeland plant holds high ecological importance in desert conservation by providing a vegetation cover along with other species. This study investigated the impact of biotic and abiotic factors across different habitats, resulting in intraspecific variations in different ecotypes of Dipterygium glaucum. Materials and Methods: Morphological variations in different habitats were assessed using 2 way MANOVA and Principal Component Analysis (PCA) with SPSS 23.0 statistical software. Total phenolics were estimated using Folin and Ciocalteu reagent, chlorophyll content was determined using spectrophotometer method. Antimicrobial activity of the plant extract was assessed against 2 g positive and 2 g negative pathogenic bacterial strains. Results: Variations among populations of three habitats due to adaption to varied environmental conditions were significant. The highland variant showed robust growth compared to coastal and sand dune populations and produced 2 types of flowers white and yellow compared to only yellow flowers in other habitats. Decreased chlorophyll content and increased antimicrobial activity and total phenolics were observed under salt stress. Conclusion: Dipterygium glaucum exhibit phenotypic plasticity across different habitats resulting in intraspecific variations and exist as ecotypes.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Neelam Sherwani and Sardar Abulfazal Farooq, 2019. Impact of Habitat Heterogeneity on Growth Dynamics and Physiological Responses of Dipterygium glaucum. Asian Journal of Plant Sciences, 18: 75-84.
DOI: 10.3923/ajps.2019.75.84
URL: https://scialert.net/abstract/?doi=ajps.2019.75.84
DOI: 10.3923/ajps.2019.75.84
URL: https://scialert.net/abstract/?doi=ajps.2019.75.84
INTRODUCTION
Plant species exposed to heterogeneity in the environment in relation to abiotic and biotic factors across different habitats, encounter selective pressures1. These pressures force a single genotype to produce different morphological and physiological responses when exposed to varied environmental conditions2-4 and undergo local adaptions resulting in phenotypic plasticity5-7 and thus exhibit intraspecific divergence8. These divergences among populations, over time line produce ecotypes across different habitats9-12, representing adaptive matching through phenotypic plasticity13, which is a commonly observed feature among many species. Species, which exhibit suitable response to varied environmental conditions have wide distributions. Among habitats, variables like temperature, humidity and salinity vary greatly and are useful indicators of how a species adapts to the local climatic conditions14,15, in which local adaptations increase plant tolerance to stress16,17 and produce different morphologies13.
Oman is a subtropical dry desert country bordering the Arabian Sea and hosts diverse range of habitats. Desert prevails in the interior Oman and witness extreme high temperatures with dry and arid environmental conditions, while the coastal regions are hot and humid. Dipterygium glaucum Decne is one of the most common plants found among different habitats of northern and central Oman. Locally called ‘Alga’, this species is commonly sighted along roadsides, sandy plains, wadis (dry water course), rocky slopes and coastal areas where it is often found in stunted condition as a rangeland plant. In these diverse habitats Dipterygium glaucum exist as a xerophyte, a facultative halophyte and a highland species which suggests it is highly adaptive and has evolved suitable traits to survive the adverse arid and saline conditions. This species suffers from severe aridity, where the water scarcity and soaring temperatures affect its morphological characters18. It is a dominant species while in coexistence in a community and holds prime ecological importance in restoration and conservation of desert vegetation along with other species like Acacia tortolis, Acacia nilotica, Aerva javonica, Leptodenia pyrotecnia, Zygophyllum quaterese, Calligonum comosum, Suaeda aegyptica. The D. glaucum is used as soil erosion controller19 and in rangeland cover re-establishment under grazing pressures.
In folklore medicine, several uses are enlisted for this species. It is analeptic, blood purifier, trachea dilating agent20 used in treatment of asthma, jaundice, chronic fever, redness and irritation of skin and unhealthy patches on skin.
The medicinal value of any plant results from synergy of different secondary metabolites present in the plant. The production of these secondary compounds are influenced by environmental factors like high temperature or salinity stress that alter the metabolism in plants and show quite significant differences in their accumulation. Climate, the seasons of the year, temperature, altitude and humidity can significantly affect the quality and quantity of the bio-active compounds21-23.
Phenolic compounds play an important role in protecting the plants under a biotic stress conditions, particularly the oxidative injury24,25. Under a saline habitat enhanced salinity results in an ionic imbalance and hyperosmotic stress effecting the cellular membranes and the photosynthetic system due to harmful effect of reactive oxygen species26.
A common but ecologically and medicinally important species like Dipterygium glaucum with a wide distribution under varied habitats, adapts itself to different environmental conditions and exhibit phenotypic plasticity and thus shows physiological, morphological, genetic differences and exist as ecotypes.
There are no reports yet on effect of different habitats on the physiological and the morphological variations in Dipterygium glaucum. In this study it investigated how different habitats impact Dipterygium glaucum differently and how these variations are expressed morphologically and physiologically in the plant. This study contributes to the knowledge of various survival tactics adapted by plant species, to thrive under stressful conditions.
MATERIALS AND METHODS
The study was conducted during the period January-July, 2017.
Study site: The study area was located at 3 different habitats spread across Muscat region, the north-eastern part of Oman in Arabia Peninsula. The habitats are, Seeb/Al Hail beach, 23.6284° N, 58.2669° E, Al Sifah highlands, (400 m above sea level) 23.268° N, 58.4652° E and Bowsher sand plains-23.5821° N, 58.0271° E. For each of the 3 habitats, 3 sites were selected randomly, thus a total of 9 sites which are distributed across northeastern Oman. Each species was sampled along one transect at each of the 9 sites (Fig. 1).
Plant material: Healthy Dipterygium glaucum Decne, plants growing wild in different habitats were collected between February 15th and 15th March, 2017 (Fig. 2).
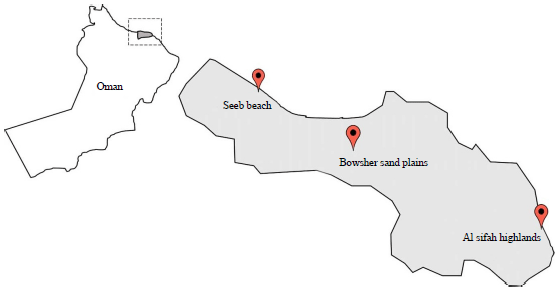 | |
| Fig. 1: | Map of Oman and the sites of sample collection |
| Al Seeb/Al Hail beach, 23.6284° N, 58.2669° E, Al Sifah highlands, (400 m above sea level) 23.268° N, 58.4652° E and Bowsher sand plains- 23.5821° N, 58.0271° E | |
 | |
| Fig. 2: | Dipterygium gluacum in natural habitat and in flowering |
Voucher specimens were compared with the herbarium sheets at Herbarium, Department of Biology, Sultan Qaboos University, Oman.
For morphometric analysis 10 plants from each habitat were assessed and morphological data was recorded on the height of the mature plant, branch length, diameter of the stem, length of root, diameter of root, total number of leaves, total number of flowers, total number of fruits, size of the flower and internode length.
Chemicals: All the chemicals that used were of analytical grade purchased from Sigma (Sigma-Aldrich, Germany) and Merck (Darmstadt, Germany).
Chlorophyll extraction: Chlorophyll was extracted from 1 g fresh leaves using 10 mL of 80% acetone27, filtered and the final volume adjusted to 100 mL. The absorbance of the solution was read at 480, 645 and 663 nm against the solvent (acetone) blank. The amounts of photo-synthetic pigments were expressed as mg g–1 fresh weight.
Calculations were done according to following equations:
| • | Total chlorophyll: 20.2 (A645)+8.02 (A663) |
| • | Chlorophyll a: 12.7 (A663)-2.69 (A645) |
| • | Chlorophyll b: 22.9 (A645)-4.68 (A663) |
![]()
Determination of total phenolics: The amount of total phenolic content was determined spectrophotometrically28 by folin-ciocalteu reagent. About 0.5 mL of methanol extract of Dipterygium glaucum was mixed 2.5 mL of 10% Folin-Ciocalteu reagent and 2.5 mL of sodium carbonate (7.5%), the mixture was allowed to stand for 15 min in parafilm covered tubes. Absorbance of the mixture was recorded at 765 nm against a blank, composed of methanol, folin-ciocalteu and sodium carbonate. Gallic acid was used as a standard. The total phenolic contents were expressed as milligrams of gallic acid equivalents per gram (mg GAE g–1 extract).
Preparation of extract: Shade dried plant material was grounded to fine powder , 100 g of the powder was extracted for 24 h in 300 mL 100% methanol in a rotatory shaker at room temperature. The residue was re-extracted under the same conditions three times. Extract was filtered and evaporated to dryness using a rotary evaporator. Finally, the residue was re-suspended in dimethyl sulfoxide (DMSO) to yield 100 mg mL–1 extract.
Micro-organisms: Bacterial strains used in the investigation were the ATCC cultures of: Staphylococcus aureus (ATCC 29213) Escherichia coli (ATCC 9637), Pseudomonas aeruginosa (ATCC 10231) and Bacillus subtilis (ATCC 6633).
Disc diffusion method: The antibacterial activity of the extract was determined by the disc diffusion method. Different strains of bacteria were cultured in a nutrient broth at 37°C for 24 h. About 6 mm sterilized discs were soaked with 100 μL of the test solution and placed on the agar plates inoculated with different bacteria. Ampicillin (1 mg disc–1) was employed as a positive control and DMSO as a negative control. Plates were incubated for 24 h at 37°C and the antibacterial activity was assessed by measuring the inhibition zones in millimeters. The experiment was run in triplicate.
Statistical analysis: Statistical analyses were done using SPSS 23.0. Two-way MANOVA was applied to test for intraspecific differences in the morphometric characters among the different habitat ecotypes. Means were compared using post hoc Tukey’s HSD at α = 0.05. Descriptive statistics were generated, listing the mean, standard deviation and coefficient of variation for each morphological character. Spearman’s ρ correlation coefficient was calculated to check correlation between mean of different growth parameters. Principal component analysis (PCA) was performed using the correlation matrix on the morphometric data in order to understand the patterns of variations within the populations from different habitats. The principal component axes were extracted according to the eigenvalues and biplot of scores of first two axes of principal component analysis was used to identify the best suited habitat and study the interrelationship among different habitats.
RESULTS
Morphological variations among growth parameters
MANOVA analysis: Morphological variations among populations were high. The MANOVA analysis for variables showed significant differences between growth parameters of plants collected from different habitats (Wilks <0.01) (Table 1).
Highly significant differences among populations were observed in plant height, branch length (Fig. 3a), stem diameter, flower size, fruit size (Fig. 3b), number of flowers and fruits (Fig. 3c). All of these characters were highly expressed under Al Sifah highland conditions followed by beach and least expressed in water stressed sand plains. The flower size in highland variants was 6 times as compared to desert morphotype. Further highland morphotypes bear two flower colors, yellow and white, while only yellow flowers were observed at other habitats. Similar trend was seen in fruit size where an average fruit in highland variant was double the size of a desert morphotype,
Roots were longer in desert morphotype, where Dipterygium glaucum was growing as a xerophyte in water limited environment as compared to beach and highland types.
| Table 1: | Results of the multi-variate analysis of variance (MANOVA) in Dipterygium glaucum using eleven morphological traits as dependent variables and three habitats as independent factors |
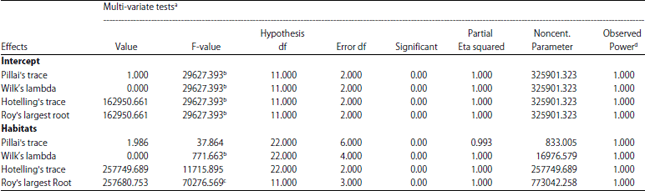 | |
a: Design: Intercept+Habitats, b: Exact statistic, c: Statistic is an upper bound on F-value that yields a lower bound on the significance level, d: Computed using alpha = 0.05 | |
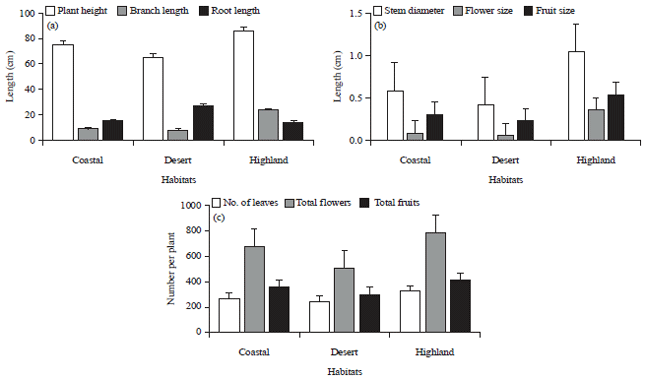 | |
| Fig. 3(a-c): | Morphological variations (mean±SD) in Dipterygium glaucum populations recorded at three different habitats, (a) Plant height, branch length and root length in coastal, desert and highland habitats, (b) Variation in flower, fruit size and (c) Variation in number of leaves, flowers and fruits per plant |
| Table 2: | Proportion of explained variance (%) of growth parameters of Dipterygium glaucum along different axis of principal component analysis |
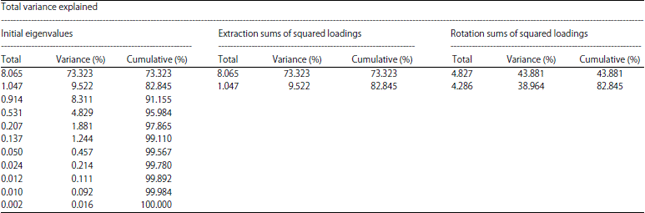 | |
| Extraction method: Principal component analysis | |
While a reverse trend was observed for root diameter where roots of coastal plants were 6 times thicker than the desert plants (Fig. 4a and b).
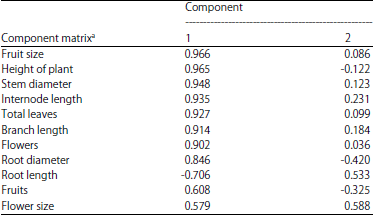
PCA analysis: The PCA after varimax rotation, extracted two PC factors with eigenvalues >1 explaining 82.845% of the variance (Table 2). The PCs plots by sampling site for the different morphological characters for the two PCs are shown in Table 3. The PCI which showed 73.323% of the total variation was highly positively correlated with traits such as total height, stem diameter, fruit size, number of leaves, except the root length which was negatively correlated with PC 1. Loadings on PC 2 which showed 9.522% (Table 2) of the total variation was positively correlated with traits like root length and flower size and negatively with root diameter.
The biplot of the PCA, clearly segregates the three distinct ecosystems (Fig. 5) as they formed non-overlapping groups and showed a considerable phenotypic variability. The high and positive loading of Al Sifah highland variants on PC1 (Fig. 5), which is positively correlated (Table 3) with most of the growth parameters suggested D. glaucum thrived best in this particular habitat and produced robust individuals.
| Table 3: | Scores of variance of morphological characters along the first two principal axes of a PCA |
 | |
| Extraction method: Principal component analysis, a: 2 components extracted | |
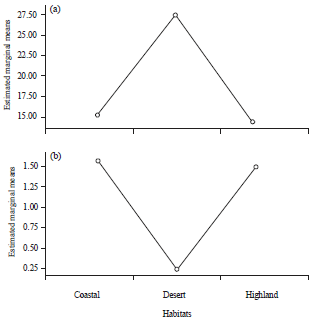 | |
| Fig. 4(a-b): | Variations in estimated marginal means of (a) Root length and (b) Root diameter from coastal, desert and highland populations |
Negative values on PC1 for the water stressed desert morphotypes makes this habitat the least favorable for most of the vegetative and reproductive traits except for root length, which was positively correlated with PC2. The loading of coastal morphotypes on the lower positive and negative sides of the PC1 suggested this habitat is not as favorable as highland conditions.
As the correlation among the growth parameters in the biplot is indicated by the directional vectors, clearly a strong correlation (Fig. 5) between the plant height, total flowers, total fruits, number of leaves, stem diameter, branch length can be observed as the angles between the directional vectors were at <90°. Root length was negatively correlated with most of other growth parameters as the angles among the directional vectors are at >90°. The PCA revealed that most of the morphological characters showed the highest value in the Al Sifah highland, except for the root diameter, which was the thickest in coastal morphotypes and root length which was longest in desert plants.
Chlorophyll content: A smaller leaf area and a seasonal leaf fall in dry conditions in this plant is compensated by occurrence of a photosynthesizing chlorenchyma tissue below the epidermis of stem. The content of total chlorophyll varied among different habitats and was 1.24 mg g–1 of the leaf fresh weight in highland plants, 1.127 mg g–1 in desert plants and 1.06 mg g–1 of the leaf fresh weight in coastal plants under increased salinity.
The content of chlorophyll-a was significantly greater than that of chlorophyll-b in all variants. Increased salinity at beach marginally decreased the chlorophyll-b content as compared to highland and desert conditions.
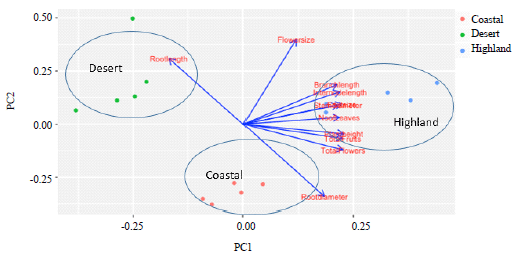 | |
| Fig. 5: | Biplot representation of the scores on the first two axes of the principal component analysis of 11 morphological characters of Dipterygium gluacum across coastal, desert and highland habitats |
| Table 4: | Antibacterial activity of Dipterygium gluacum against 4 bacterial strains |
 | |
Total phenolic content: The total phenolic content of the methanolic extract of the plants collected from desert was 65.78 and 68.35 mg g-1 GAEs for highland plants, while for coastal plants an increased total content of 73.2 mg g-1 GAEs was observed.
Antimicrobial activity: The antimicrobial activity of the extracts of Dipterygium glaucum was studied against four pathogenic bacterial strains (Table 4), 2 Gram-positive (Bacillus subtilis, Staphylococcus aureus) and 2 Gram-negative (Escherichia coli and Pseudomonas aeruginosa). Methanolic plant extracts from different habitats exhibited antibacterial activity against B. subtilis and P. aeruginosa (Table 4). Variation in inhibition zones was observed across habitats, where coastal ecotypes exhibited maximum inhibition and least was observed in Al Sifah highlands. Inhibition was more for Gram-negative P. aeruginosa as compared to Gram-positive B. subtilis. No inhibition was observed against both S. aureus and E. coli.
DISCUSSION
By exhibiting phenotypic plasticity, a plant species produces different morphotypes according to prevailing environmental conditions. The adaptive role of phenotypic plasticity in various plant species has been recognized by different studies29,30. A range of suitable habitats for the growth of Dipterygium glaucum has been revealed by this study and this species exhibited variability in its morphological characters under the influence of varied environmental conditions.
The results of MANOVA confirmed that environmental factors influenced the morphological differences between the three populations of D. glaucum across its range. According to earlier studies31-34, plant species demonstrated local adaptation along latitudinal and elevation gradients. The PCA revealed that the Al Sifah high altitude conditions were more favorable to most of the growth parameters (i.e., both vegetative and reproductive), there by indicating better growth conditions in terms of moisture availability and slightly lower temperatures. The MANOVA results further revealed that the coastal habitat conditions were relatively better for the growth of D. glaucum as compared to the inland sand dunes, where most of the characters were minimally expressed. Plasticity plays an important role in adaptive responses to the changing environment as it allows a plant to express maximally in terms of phenotype under optimal conditions and tolerate the harsh environment as suboptimal conditions35,36.
The results of principal component analysis supports the fact that the differences recorded in the multi-variate analysis (MANOVA) were real differences that did not occur simply by randomized chance. Striking phenotypic and physiological differences between two ecotypes has been recorded and documented by numerous studies37-40.
Though Dipterygium glaucum is a sand dune-forming species of high ecological importance in view of desert vegetation restoration and conservation, species suffers from severe aridity, where the lack of water and very high temperature can affects its growth performance as compared to coastal and highland variants. Different plant species under water stress recorded reduction in stem diameter41. On the contrary, root length was almost double in sand plain habitats as compared to beach and highland ecotypes. Arid desert plants develop taproots that descend to deeper soil layers to gather water that is far down, producing few lateral roots within top 1m of soil with fine small lateral roots produced in deeper soil layers42. Longer root length under water stress conditions has been observed in previous studies42,43. Root diameter increased profoundly under salinity stress due to succulence of cortex, resulting in increased volume of vacuoles which permits accumulation of larger amount of water and dissolved ions44. Similar halophytic response has been recorded by earlier studies45.
Decrease in chlorophyll content per unit of leaf area under increased salinity was observed in our study. There is an overwhelming evidence of salinity stress affecting the photosynthetic enzymes, chlorophyll and carotenoids content45-47. Accumulation of ions of different salts induces an inhibitory effect on the biosynthesis of the different fractions of chlorophyll thereby reducing the overall chlorophyll and carotenoids content48.
Higher total phenolic content was recorded in D. glaucum coastal plants as compared to the other ecotypes. Accumulation of phenolic compounds under salt stress in plants is reported by many studies49-53. In order to overcome oxidative stress due to accumulation of reactive oxygen species under salt stress, plants develop adaptive mechanisms in order to reduce the oxidative damage. The mechanism involves production of phenolic compounds to in order to scavenge the free radicals.
The plant extract from different habitats possess antibacterial activity against some of the tested organisms. Previous studies have documented antimicrobial activity in Dipterygium glaucum20,54. The diameter of inhibition zones varied among different ecotypes suggesting different phytoconstituents accumulated under different environmental conditions under different habitats. Maximum inhibition was observed for coastal morphotype suggesting more bio-active compounds being produced by the plants under salinity stress. Variation in antimicrobial activity in plant extracts across different habitats has been documented by many studies55,56.
CONCLUSION
Significant morphological plasticity among ecotypes of Dipterygium glaucum exist along different habitats. This species exist under a range of habitats but highland conditions are favorable to most of the vegetative and reproductive traits indicating better growth conditions at this habitat as compared to the salinity stressed coastal habitat and water stressed sand plains. Under the influence of various abiotic stresses, this species undergo morphological and physiological adaptations in order to survive.
SIGNIFICANCE STATEMENT
This study reinforced and justified the use of this species as a herbal remedy in traditional medicine as the methanolic extract of the plant exhibited antimicrobial activity against B. subtilis and P. aeruginosa and an adequate amount of total phenolics were observed. Being part of diet of important rangeland animals like Gazelle and an important species of community structure involved in the restoration of desert vegetation this study suggests that Dipterygium gluacum is morphologically and physiologically accustomed to variation in water and temperature and adapts itself to diverse environment conditions. This study will provide useful insight to researchers to understand the adaptations undergone by plants of arid zones to survive under harsh environmental conditions and aid in conservation of the flora of arid zones.
REFERENCES
- Valladares, F., E. Gianoli and J.M. Gomez, 2007. Ecological limits to plant phenotypic plasticity. New Phytol., 176: 749-763.
CrossRefDirect Link - Lazaro-Nogal, A., S. Matesanz, A. Godoy, F. Perez-Trautman, F. Gianoli and F. Valladares, 2015. Environmental heterogeneity leads to higher plasticity in dry‐edge populations of a semi‐arid Chilean shrub: Insights into climate change responses. J. Ecol., 103: 338-350.
CrossRefDirect Link - Sultan, S.E., 1995. Phenotypic plasticity and plant adaptation. Acta Bot. Neerl., 44: 363-383.
CrossRefDirect Link - Aranda, I., H.A. Bahamonde and D. Sanchez-Gomez, 2017. Intra-population variability in the drought response of a beech (Fagus sylvatica L.) population in the Southwest of Europe. Tree Physiol., 37: 938-949.
CrossRefDirect Link - Nicotra, A.B. and A. Davidson, 2010. Adaptive phenotypic plasticity and plant water use. Funct. Plant Biol., 37: 117-127.
CrossRefDirect Link - Voltas, J., T.A. Shestakova, T. Patsiou, G. Di Matteo and T. Klein, 2018. Ecotypic variation and stability in growth performance of the thermophilic conifer Pinus halepensis across the Mediterranean basin. For. Ecol. Manage., 424: 205-215.
CrossRefDirect Link - Olivas, N.H.D., E. Frago, M.P.M. Thoen, K.J. Kloth and F.F.M. Becker et al., 2017. Natural variation in life history strategy of Arabidopsis thaliana determines stress responses to drought and insects of different feeding guilds. Mol. Ecol., 26: 2959-2977.
CrossRefDirect Link - Linnen, C.R. and H.E. Hoekstra, 2009. Measuring natural selection on genotypes and phenotypes in the wild. Cold Spring Harb. Symp. Quant. Biol., 74: 155-168.
CrossRefDirect Link - Agren, J. and D.W. Schemske, 2012. Reciprocal transplants demonstrate strong adaptive differentiation of the model organism Arabidopsis thaliana in its native range. New Phytol., 194: 1112-1122.
CrossRefDirect Link - Lowry, D.B., K.D. Behrman, P. Grabowski, G.P. Morris, J.R. Kiniry and T.E. Juenger, 2014. Adaptations between ecotypes and along environmental gradients in Panicum virgatum. Am. Nat., 183: 682-692.
CrossRefDirect Link - Sultan, S.E., 2005. An emerging focus on plant ecological development. New Phytol., 166: 1-5.
CrossRefDirect Link - Sundqvist, M.K., N.J. Sanders and D.A. Wardle, 2013. Community and ecosystem responses to elevational gradients: Processes, mechanisms and insights for Global change. Ann. Rev. Ecol. Evol. Syst., 44: 261-280.
CrossRefDirect Link - Manzano-Piedras, E., A. Marcer, C. Alonso-Blanco and F.X. Pico, 2014. Deciphering the adjustment between environment and life history in annuals: Lessons from a geographically-explicit approach in Arabidopsis thaliana. PLoS One, Vol. 9, No. 2.
CrossRefDirect Link - Matesanz, S. and F. Valladares, 2014. Ecological and evolutionary responses of Mediterranean plants to global change. Environ. Exp. Bot., 103: 53-67.
CrossRefDirect Link - Gimeno, T.E., B. Pias, J.P. Lemos-Filho and F. Valladares, 2008. Plasticity and stress tolerance override local adaptation in the responses of Mediterranean holm oak seedlings to drought and cold. Tree Physiol., 29: 87-98.
CrossRefDirect Link - Moussa, S.A.I., W.K. Taia and F.G.M. Al-Ghamdy, 2012. Acclimation of Dipterygium glaucum Decne grown in the Western Coastal part of Saudi Arabia to different water supplies. Int. J. Res. Chem. Environ., 2: 301-309.
Direct Link - Altwaty, N.H., O.E. El-Sayed, N.A.H. Aly, M.N. Baeshen and N.A. Baeshen, 2016. Molecular and cytogenetic assessment of Dipterygium glaucum genotoxicity. An. Acad. Bras. Cienc., 88: 623-634.
CrossRefDirect Link - Shaheen, U., N.A. Shoeib, A. Temraz and M.I.S. Abdelhady, 2017. Flavonoidal constituents, antioxidant, antimicrobial and cytotoxic activities of Dipterygium glaucum grown in Kingdom of Saudi Arabia. Pharmacogn. Mag., 13: S484-S488.
CrossRefPubMedDirect Link - De Abreu, I.N. and P. Mazzafera, 2005. Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiol. Biochem., 43: 241-248.
CrossRefDirect Link - Szakiel, A., C. Paczkowski and M. Henry, 2011. Influence of environmental abiotic factors on the content of saponins in plants. Phytochem. Rev., 10: 471-491.
CrossRefDirect Link - Pavarini, D.P., S.P. Pavarini, M. Niehues and N.P. Lopes, 2012. Exogenous influences on plant secondary metabolite levels. Anim. Feed Sci. Technol., 176: 5-16.
CrossRefDirect Link - Ahmad, P., C.A. Jaleel, M.A. Salem, G. Nabi and S. Sharma, 2010. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol., 30: 161-175.
CrossRefDirect Link - Bose, J., A. Rodrigo-Moreno and S. Shabala, 2014. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot., 65: 1241-1257.
CrossRefDirect Link - Jithesh, M.N., S.R. Prashanth, K.R. Sivaprakash and A.K. Parida, 2006. Antioxidative response mechanisms in halophytes: Their role in stress defence. J. Genet., 85: 237-254.
CrossRefDirect Link - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Singleton, V.L., R. Orthofer and R.M. Lamuela-Raventos, 1999. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In: Methods in Enzymology, Burslem, G.L. (Ed.), Academic Press, Cambridge, Massachusetts, ISBN: 9780121822002, pp: 152-178.
CrossRefDirect Link - Miner, B.G., S.E. Sultan, S.G. Morgan, D.K. Padilla and R.A. Relyea, 2005. Ecological consequences of phenotypic plasticity. Trends Ecol. Evol., 20: 685-692.
CrossRefDirect Link - Mony, C., G. Thiebaut and S. Muller, 2006. An experimental study of the plastic responses of Ranunculus peltatus Schrank to four environmental parameters. Hydrobiologia, 570: 41-46.
CrossRefDirect Link - Hereford, J., 2009. A quantitative survey of local adaptation and fitness trade-offs. Am. Nat., 173: 579-588.
CrossRefDirect Link - Alberto, F.J., S.N. Aitken, R., Alia, S.C. Gonzalez-Martinez and H. Hanninen et al., 2013. Potential for evolutionary responses to climate change-evidence from tree populations. Global Change Biol., 19: 1645-1661.
CrossRefDirect Link - Debieu, M., C. Tang, B. Stich, T. Sikosek and S. Effgen et al., 2013. Co-variation between seed dormancy, growth rate and flowering time changes with latitude in Arabidopsis thaliana. PLoS One, Vol. 8, No. 5.
CrossRefDirect Link - Halbritter, A.H., R. Billeter, P.J. Edwards and J.M. Alexander, 2015. Local adaptation at range edges: Comparing elevation and latitudinal gradients. J. Evol. Biol., 28: 1849-1860.
CrossRefDirect Link - Hendry, A.P., 2016. Key questions on the role of phenotypic plasticity in eco-evolutionary dynamics. J. Hered., 1071: 25-41.
CrossRefDirect Link - Pfennigwerth, A.A., J.K. Bailey and J.A. Schweitzer, 2017. Trait variation along elevation gradients in a dominant woody shrub is population-specific and driven by plasticity. AoB Plants, Vol. 9, No. 4.
CrossRefDirect Link - Ellison, A.M., H.L. Buckley, T.E. Miller and N.J. Gotelli, 2004. Morphological variation in Sarracenia purpurea (Sarraceniaceae): Geographic, environmental and taxonomic correlates. Am. J. Bot., 91: 1930-1935.
CrossRefDirect Link - Cortese, L.M., J. Honig, C. Miller and S.A. Bonos, 2010. Genetic diversity of twelve switchgrass populations using molecular and morphological markers. Bioenergy Res., 3: 262-271.
CrossRefDirect Link - Gratani, L., 2014. Plant phenotypic plasticity in response to environmental factors. Adv. Bot., Vol. 2014.
CrossRef - Milano, E.R., D.B. Lowry and T.E. Juenger, 2016. The genetic basis of upland/lowland ecotype divergence in switchgrass (Panicum virgatum). G3: Genes Gen. Genet., 6: 3561-3570.
CrossRefDirect Link - Taia, W.K. and S.A.I. Moussa, 2011. Phenotypic variations in Calligonum comosum L’Her (Polygonaceae) grown in Al-Nefud Desert in Saudi Arabia. J. Scient. Res., 3: 421-435.
CrossRefDirect Link - Chaves, M.M., J.S. Pereira, J. Maroco, M.L. Rodrigues and C.P.P. Ricardo et al., 2002. How plants cope with water stress in the field? Photosynthesis and growth. Ann. Bot., 89: 907-916.
CrossRefDirect Link - Saidi, A., T. Ookawa, T. Motobayashi and T. Hirasawa, 2008. Effects of soil moisture conditions before heading on growth of wheat plants under drought conditions in the ripening stage: Insufficient soil moisture conditions before heading render wheat plants more resistant to drought during ripening. Plant Prod. Sci., 11: 403-414.
CrossRefDirect Link - Misra, A.N., D. Latowski and K. Strzalka, 2006. The xanthophyll cycle activity in kidney bean and cabbage leaves under salinity stress. Russian J. Plant Physiol., 53: 102-109.
CrossRefDirect Link - Taffouo, V.D., O.F. Wamba, E. Youmbi, G.V. Nono and A. Akoa, 2010. Growth, yield, water status and ionic distribution response of three bambara groundnut (Vigna subterranean (L.) Verdc.) landraces grown under saline conditions. Int. J. Bot., 6: 53-58.
CrossRefDirect Link - Heidari, A., M. Toorchi, A. Bandehagh and M.R. Shakiba, 2011. Effect of NaCl stress on growth, water relations, organic and inorganic osmolytes accumulation in Sunflower (Helianthus annuus L.) lines. Universal J. Environ. Res. Technol., 1: 351-362.
Direct Link - Ali, Y., Z. Aslam, M.Y. Ashraf and G.R. Tahir, 2004. Effect of salinity on chlorophyll concentration, leaf area, yield and yield components of rice genotypes grown under saline environment. Int. J. Environ. Sci. Technol., 1: 221-225.
CrossRefDirect Link - Valifard, M., S. Mohsenzadeh, B. Kholdebarin and V. Rowshan, 2014. Effects of salt stress on volatile compounds, total phenolic content and antioxidant activities of Salvia mirzayanii. South Afr. J. Bot., 93: 92-97.
CrossRefDirect Link - Salem, N., K. Msaada, W. Dhifi, F. Limam and B. Marzouk, 2014. Effect of salinity on plant growth and biological activities of Carthamus tinctorius L. extracts at two flowering stages. Acta Physiol. Planta., 36: 433-445.
CrossRefDirect Link - Shao, Y., F. Xu, X. Sun, J. Bao and T. Beta, 2014. Identification and quantification of phenolic acids and anthocyanins as antioxidants in bran, embryo and endosperm of white, red and black rice kernels (Oryza sativa L.). J. Cereal Sci., 59: 211-218.
CrossRefDirect Link - Zhao, X., W. Zhang, X. Yin, M. Su, C. Sun, X. Li and K. Chen, 2015. Phenolic composition and antioxidant properties of different peach [Prunus persica (L.) Batsch] cultivars in China. Int. J. Mol. Sci., 16: 5762-5778.
CrossRefDirect Link - Caliskan, O., J. Radusiene, K.E. Temizel, Z. Staunis, C. Cirak, D. Kurt and M.S. Odabas, 2017. The effects of salt and drought stress on phenolic accumulation in greenhouse-grown Hypericum pruinatum. Ital. J. Agron., 11: 271-279.
CrossRefDirect Link - Ahmed, S., M. Ashraf, A. Jabbar, K.H. Janbaz, M.S. Khan, Nisar-ur-Rahman and M. Hussain, 2006. Biological activity of Dipterygium glaucum. Pak. J. Biol. Sci., 9: 1173-1174.
CrossRefDirect Link - Amdouni, T., S.B. Abdallah, N. Msilini, F. Merck and M. Chebbi et al., 2016. Effect of salt stress on the antimicrobial activity of Ruta chalepensis essential oils. Acta Physiol. Planta., Vol. 38, No. 6.
CrossRefDirect Link - Karahan, F., C. Avsar, I.I. Ozyigit and I. Berber, 2016. Antimicrobial and antioxidant activities of medicinal plant Glycyrrhiza glabra var. glandulifera from different habitats. Biotechnol. Biotechnol. Equip., 30: 797-804.
CrossRefDirect Link