
D. Montcho
Africa Rice Center (AfricaRice), 01 BP2031Cotonou, Benin
K. Futakuchi
Africa Rice Center (AfricaRice), 01 BP2031Cotonou, Benin
C. Agbangla
University of Abomey-Calavi, BP 526 Abomey-Calavi, Benin
M. Semon
Africa Rice Center (AfricaRice), PMB 5320, Ibadan, Nigeria
I. Dieng
Africa Rice Center (AfricaRice), 01 BP2031Cotonou, Benin
M. Sie
Africa Rice Center (AfricaRice), 01 BP2031Cotonou, Benin
Asian Journal of Plant Sciences
Year: 2013 | Volume: 12 | Issue: 2 | Page No.: 79-86







ABSTRACT
Oryza glaberrima is often classified into two ecological groups: upland and floating types. To assess the phenotypic variability of O. glaberrima across hydrological conditions, 217 accessions and 5 checks (Oryza sativa) were cultivated using toposequence. The trial was conducted at the experimental field of the International Institute of Tropical Agriculture in Ibadan during the wet seasons in 2009 and 2010. An Alpha Lattice design with three replications was used. Phenotypic data including vegetative vigor, plant height and tillers number at maturity, days to 50% heading and crop duration were recorded. The results indicated high variability among all O. glaberrima tested across ecologies for plant height, tiller ability and crop duration. This variability was confirmed by low genetic correlation coefficients, which demonstrated the significance of genotype by environment interaction. Oryza glaberrima accessions tested showed best vegetative vigor from upland to lowland. In 2009, 77.5 and 82.2% of the accessions were tall in hydromorphic and lowland, respectively while they represented 87.5 and 84.8%, respectively in 2010. Tillers number was intermediate from upland to lowland in both years. Result in 2009 indicated that 54.5% in upland, 70.9% in hydromorphic and 45.3% in lowland of O. glaberrima were intermediate tillering. The second season showed 63.2% in upland, 79.1% in hydromorphic and 82.1% in lowland. High percentage of O. glaberrima accessions was observed as late and very late maturing in 2009, 89.9% in upland and 91.4% in lowland. This study support efforts to breed for phenological plasticity in any rice growing environment.
PDF Abstract XML References Citation
Received: February 28, 2013;
Accepted: May 04, 2013;
Published: July 03, 2013
How to cite this article
D. Montcho, K. Futakuchi, C. Agbangla, M. Semon, I. Dieng and M. Sie, 2013. Morphological and Phenological Response of African Rice (Oryza glaberrima Steud.) under Different Hydrological Conditions. Asian Journal of Plant Sciences, 12: 79-86.
DOI: 10.3923/ajps.2013.79.86
URL: https://scialert.net/abstract/?doi=ajps.2013.79.86
DOI: 10.3923/ajps.2013.79.86
URL: https://scialert.net/abstract/?doi=ajps.2013.79.86
INTRODUCTION
Two species of cultivated rice are known: Oryza sativa L. and Oryza glaberrima Steud. Oryza sativa or Asian rice was domesticated more than 10,000 years ago from O. rufipogon (Kovach et al., 2007) and has been adapted to a wide range of conditions including different water regimes and a wide range of soils (Takahashi, 1984). Oryza sativa includes two ecologically different subspecies. Oryza sativa japonica is cultivated in the upland ecology, while the other ecologies are occupied by O. sativa indica. Oryza glaberrima (or African Rice) was domesticated some 2,000-3,000 years ago (Porteres, 1970) from the wild ancestor O. barthii in the floodplains of the bend of the Niger River, which is the primary center of origin (Sarla and Swamy, 2005). Oryza glaberrima is increasingly replaced in Africa rice growing environment by O. sativa which has been introduced by the Portuguese (Linares, 2002). Many African farmers are shifting from O. glaberrima to O. sativa because of its low yielding caused by grain shattering and lodging susceptibility (Linares, 2002). Oryza glaberrima accessions are considered as a rich reservoir of genes and alleles suitable for the genetic improvement of rice targeting the African continent’s rice growing constraints (Jones et al., 1997). Previous research work in interspecific crossing has helped to combine weed competitiveness of O. glaberrima and high yield potential of O. sativa japonica (Jones et al., 1997). Moreover, O. glaberrima is considered to have early growth vigor, which is the most important character in weed competitiveness (Dingkuhn et al., 1998, 1999; Saito et al., 2010). Oryza glaberrima was ecologically divided into two groups (Oka, 1974): upland and deep water types. However, other studies pinpointed that there are no sub-groups in O. glaberrima (Morishima et al., 1962). In support of this opinion, some upland O. glaberrima accessions have been shown to have resistance to lowland constraints (Futakuchi and Sie, 2009), while so-called upland types and deep-water types showed the same response to submergence (Futakuchi and Jones, 2005). On the other hand, Semon et al. (2005) demonstrated that population can be divided into five different groups using genetic diversity analysis mean. Knowledge of ecological adaptation to variable water regimes is limited, especially for O. glaberrima. This study further considers the morphological and phenological response of O. glaberrima across the hydrological toposequence from upland to irrigated lowland.
MATERIALS AND METHODS
Site characteristics: The study was conducted at the experimental farm of the International Institute of Tropical Agriculture (IITA) Ibadan station, Nigeria. The station is located at latitude 7°3’N and longitude 3°45’E in the forest-savanna agro-ecology. Rainfall during the wet seasons (from May to December) was 872.25 mm in 2009 and 1680.25 mm in 2010 (Fig. 1).
Plant materials: Two hundred and seventeen accessions of O. glaberrima were selected with the objective of covering the geographic origins of African rice. Table 1 shows the frequencies distribution of the different accessions of O. glaberrima involved in this experiment across countries. Five O. sativa cultivars were included in the experiment as checks: IDSA 6, Morobérékan and WAB56-104 adapted to upland conditions and IR 64 and WITA 4 adapted to lowland conditions. The seeds were obtained from the Africa Rice Center germplasm collection.
Experimental design and crop management: Seeds of all accessions were sown at three levels in the toposequence (upland, non-flooded hydromorphic and irrigated lowland) at the IITA station experimental field in Ibadan, Nigeria. In the upland and hydromorphic plots, dry seeds were dibbled, whereas pre-germinated seeds were sown in a nearby seedbed nursery and transplanted into the main plots in lowland at 20 Days After Sowing (DAS). An Alpha Lattice design with three replications was used at each level. Before sowing or transplanting, the field was tilled manually using hoes. Plots were fertilized with 200 kg ha-1 of NPK as a basal (15-15-15) dressing at 25 DAS and with 50 kg ha-1 of urea at 45 DAS. Weeding was done by hand to avoid possible herbicide effects on crop phenology. Plots were kept weed-free by regular hand weeding.
In the lowland field, groundwater level was maintained at 5 to 10 cm depth. In upland and hydromorphic conditions, water level below the soil surface was determined using 6 cm diameter Poly Vinyl Chloride (PVC) tubes.
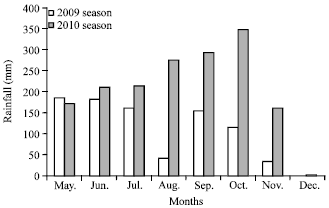 | |
| Fig. 1: | Rainfall in 2009 and 2010 (source IITA Ibadan) |
| Table 1: | Distribution of plant material across geographic origins |
 | |
Piezometers were inserted to 80 cm soil depth at three randomly selected positions in each replicate. Each 100 cm piezometer was made with 80 cm of perforations from the bottom up; 20 cm was above ground and covered by plastic sheet. The experiment was performed during the 2009 and 2010 wet seasons but differed in the sowing date, which was May 15 in 2009 and July 2 in 2010.
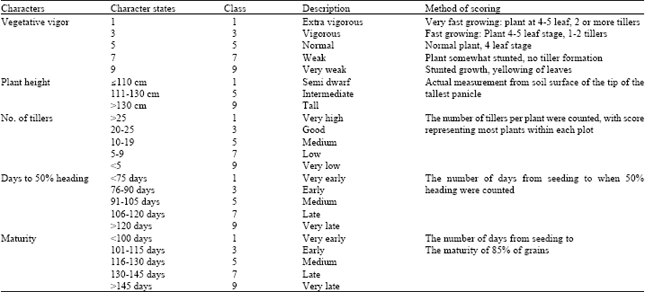
Measurements of agro-morphological characters: Water level in each PVC tube was determined daily using a long ruler. Data were collected from four plants randomly identified and labeled, selected from the middle plants in each experimental plot. Method of data measurement is shown in Table 2. Vegetative vigor was scored in each plot at 30 DAS. Plant height and tiller number were measured at maturity. Phenological response including days to 50% heading and maturity (crop duration) were visually assessed on a daily basis starting from booting stage on all plots throughout the experiment. Soil samples at different depths (0-20; 20-40 cm) were collected separately at each ecosystem before crop establishment to determine major chemical components and soil physical characteristics.
| Table 2: | List of characters and their respective classes used for the evaluation of diversity |
 | |
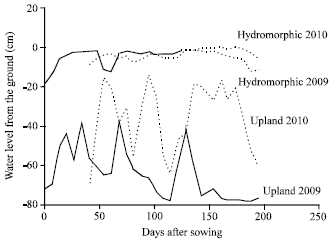 | |
| Fig. 2: | Fluctuation of water levels in 2009 and 2010 |
Statistical data analysis: The phenotypic frequencies of the five characters were analyzed by the Shannon-weaver index (H’) using the formula of Jain et al. (1975):
where, k is the number of phenotypic classes for the character and Pi is the proportion of the total number of entries in that class. H’ was estimated for each character by ecology. Each value of H’ was standardized by dividing it by its maximum value (log2k), in order to keep the values in the range of 0-1 (Abdi et al., 2002).
Analysis of variance was performed for non-transformed Shannon-weaver index.
The genetic correlation rgxy between two ecologies (x and y) was calculated according to Burdon (1977):
where, Covgxy is the covariance of both ecologies x and y, σgx is the standard deviation for ecology x and σgy is the standard deviation for ecology.
RESULTS
Groundwater fluctuations: Piezometric characterization of the experimental sites (upland and hydromorphic) was important to knowing the water availability during plant growth, with the soil surface being considered as zero level. The rainfall-dependent fluctuations in groundwater along the toposequence were between 0 and 20 cm in the hydromorphic zone in 2009 and 2010. In the upland zone, groundwater fluctuated between 37 and 78 cm in 2009 and between 14 and 68 cm in 2010. Rainfall was greater in 2010 (1680.25 mm) than in 2009 (872.25 mm). In upland 2009, drought occurred when groundwater level was closed to -80 cm. Figure 2 shows the fluctuations in groundwater in upland and hydromorphic zones in both seasons. Globally, groundwater level was obviously higher in hydromorphic condition than in upland ecology irrespective of seasons. However, this water level was similar in hydromorphic condition across seasons while it was higher in upland in 2010 than it was in 2009.
Soil characteristics: Soil samples at different depths were analyzed to determine the chemical components and the soil physical characteristics of each ecosystem (Table 3). Using Jamagne (1967) triangle of soil texture, the soils from upland and lowland were found to be sandy and similar.
| Table 3: | Physico-chemical characteristics of soil samples collected from different ecologies |
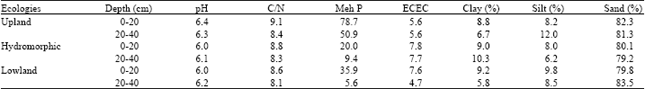 | |
| ECEC: Effective Cation exchange capacity; C: Carbon; N: Nitrogen; pH: Hydrogen potential, Meh P: Mehlich phosphorus | |
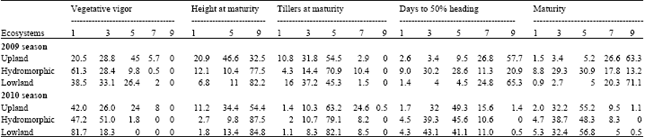
| Table 4: | Phenotypic frequencies (%) of five variables across ecologies in 2009 and 2010 |
 | |
Results showed that the Hydrogen potential (pH) of the soil was slightly acidic and ranged between 6.0 and 6.4. The ecosystems displayed a similar pattern for Carbon nitrogen ratio (C/N), which was between 8.1 and 9.1 with a low Effective Cation Exchange Capacity (ECEC) and a high concentration in phosphorus.
Morphological and phenological variability across hydrological conditions: Table 4 shows the phenotypic frequencies of five variables-vegetative vigor, plant height and number of tillers at maturity, days to 50% heading and maturity-for the 2009 and 2010 seasons. Frequencies were calculated as a percentage of the number of accessions in each class. The individuals differed from one class to another.
Monomorphic phenotypic frequency distribution was exhibited by vegetative vigor in the upland ecosystem whereas intermediate frequency was observed in hydromorphic and lowland ecosystems in 2009. In upland, 49.3% (classes 1 and 3 together) of O. glaberrima accessions used were vigorous. Nearly 90% of entries in hydromorphic ecosystem were either extra vigorous or vigorous equaling to 61.37% for class 1 and 28.4% for class 2. The lowland condition displayed 71.6% of O. glaberrima for these combined classes. Normal vigor was recorded in frequency of 45% in upland, 9.8% in the hydromorphic zone and 26.4% in lowland. The weak class bore very few individual across ecologies equaling to 5.6, 0.5 and 2%, respectively in upland, hydromorphic and lowland.
In 2010, vegetative vigor was of intermediate type in upland and lowland ecosystems. Monomorphic frequency was observed in the hydromorphic ecosystem. Forty two percent of accessions were classified as extra vigorous and 26% as vigorous in upland conditions. In hydromorphic extra vigorous and vigorous class bore 47.2 and 51% of O. glaberrima, respectively. In lowland ecosystem, extra vigorous and vigorous class indicated 81.7 and 18.3% of tested entries, respectively. The normal vigor recorded 24% in upland and only 1.8% in hydromorphic condition. The weak covered 8% of the tested O. glaberrima in the upland.
Plant height at maturity showed monomorphic phenotypic frequency in upland conditions but intermediate frequency was found in hydromorphic and lowland conditions in 2009. Most O. glaberrima accessions used were tall, except in upland conditions: 82.2% in lowland, 77.5% in hydromorphic and 32.5% upland. Under upland conditions, 46.6% of O. glaberrima tested were of intermediate height.
In 2010, intermediate phenotypic frequency was observed in all ecosystems. The majority of plants in the different ecosystems were found to be tall: 84.8% in lowland, 87.5% in hydromorphic and 54.4% in upland conditions.
Number of tillers at maturity was monomorphic in both years in all ecosystems. In 2009 the frequency of O. glaberrima having an intermediate number of tillers was: 54.5% in upland, 70.9% in hydromorphic and 45.3% in lowland. High tillering ability was observed in frequency of 37.2% of accessions in lowland, 14.4% in hydromorphic and 31.8% in upland.
In 2010, the intermediate class accounted for 63.2% in upland, 79.1% in hydromorphic and 82.1% in lowland. High tillering ability was recorded in low frequency in upland (10.3%), hydromorphic (10.7%) and lowland (8.3%) but the upland condition further displayed 24.6% of tested entries as low tillering.
In 2009, polymorphic phenotypic frequency was observed in the hydromorphic condition for days to 50% heading, whereas frequency was shown to be intermediate value in the upland and lowland ecosystems. In the upland ecosystem, the late and very late headings individuals combined at 84.5% while these of very early, early and intermediate headings were low (15.5%). In hydromorphic conditions, very early and early headings individuals were recorded for 39.2% of accessions, followed by late and very late headings (32.2%) and intermediate heading (28.6%) of O. glaberrima. Late and very late headings individuals in lowland conditions accounted for 90.1% of all accessions.
Monomorphic phenotypic frequency was given by days to 50% heading in all ecosystems in 2010. In the upland condition, the very early and early headings O. glaberrima represented were 33.7%, intermediate heading O. glaberrima accounted for 49.3% and the late and very late heading accounted for 17%. The intermediate heading entries represented 45.6% followed by the very early and early headings (43.8%) and the late heading (10.6%) in hydromorphic condition. Results obtained in lowland condition were 43.1, 41.1 and 11%, respectively for early heading, intermediate heading and late heading.
Intermediate frequency was shown for maturity in upland and lowland conditions and frequency was monomorphic in the hydromorphic condition in 2009. Late and very late maturing accessions represented together 89.9 and 91.4% in upland and lowland, respectively. In the hydromorphic condition, accessions of both class (very early and early) accounted for 38.1% followed by late and very late maturing (31%) and intermediate maturing (30.9%).
In 2010, frequency was monomorphic in all ecologies. About fifty five per cent (55.2%) of O. glaberrima were intermediate maturing in upland followed by early maturing (32.2%) and late and very late maturing (10.6%). In the hydromorphic ecosystem, intermediate maturity was recorded for 48.3% of tested entries followed by early maturing (38.7%) and late maturing (8.3%). In lowland 56.8% of O. glaberrima tested were found as intermediate maturing, 32.4% as early maturing, 5.3% as very early maturing and 5% as late maturing.
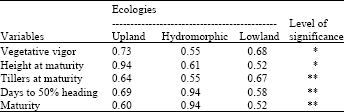
Table 5 and 6 showed the estimated Shannon-weaver diversity index (H’) values of five variables for each ecological condition in 2009 and 2010. In 2009, the analysis of variance indicated significant difference (p<0.05) across ecologies for all variables while only vegetative vigor and plant height at maturity showed significance (p<0.01) in 2010. In 2009, vegetative vigor showed H’ value of 0.73 in upland, 0.55 in hydromorphic and 0.68 in lowland conditions; plant height 0.94 in upland, 0.61 in hydromorphic and 0.52 in lowland.
| Table 5: | Estimates of Shannon-weaver diversity index in 2009 |
 | |
| *Significance at p<5% level, **Significance at p<0.01 level | |
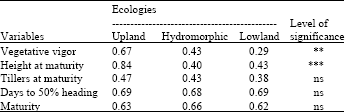
| Table 6: | Estimates of Shannon-weaver diversity index in 2010 |
 | |
| **Significance at p<0.01 level, ***Significance at p<0.001 level, ns: Non-significant | |
Number of tillers at maturity has H’ value of 0.64 in upland, 0.55 in hydromorphic and 0.67 in lowland; while days to 50% heading showed 0.69 in upland, 0.94 in hydromorphic and 0.58 in lowland. For crop duration (maturity), H’ indicated 0.60 in upland, 0.94 in hydromorphic and 0.52 in lowland conditions. In the repeated experiment in 2010, H’ values were significantly different only for vegetative vigor and plant height at maturity. Vegetative vigor indicated 0.67 in upland, 0.43 in hydromorphic and 0.29 in lowland. For plant height, Shannon-weaver index were 0.84, 0.40 and 0.43 in upland, hydromorphic and lowland, respectively. The other variables (number of tillers at maturity, days to 50% heading and crop duration) did not show significant difference from upland to lowland.
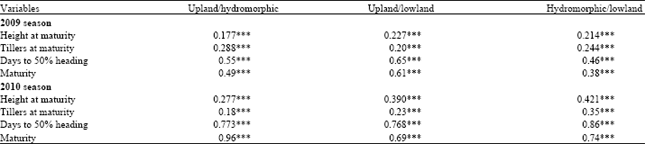
Genetic correlations rgxy between ecologies were used to estimate significant genotypexenvironment interaction (Table 7). Low values of genetic correlations were observed for plant height and tiller number at maturity in both years. When combining ecosystems, upland/hydromorphic gave 0.177, upland/lowland showed 0.227 and hydromorphic/lowland indicated 0.214 for plant height at maturity in 2009. Number of tillers at maturity revealed 0.288, 0.20 and 0.244 for the same combinations. Days to 50% heading showed 0.55 for the upland/hydromorphic combination, 0.65 for the upland/lowland combination and 0.46 for hydromorphic/lowland. Crop duration indicated 0.49, 0.61 and 0.38 for the three combinations. Positives and significant correlations were obtained for plant height, tiller number, days to 50% heading and crop duration in both seasons. The second season showed globally higher genetic correlations for all variables but little difference was observed.
| Table 7: | Genetic correlation coefficients for variables between ecosystem |
 | |
| ***Significance at p<0.001 level | |
DISCUSSION
Rice is able to grow at pH levels between 4 and 8. From upland to lowland in this study, pH values were between 6.0 and 6.4, which cannot hamper rice production. Soil physico-chemical characteristics were similar from upland to lowland. Variability of morphological and phenological response from upland to lowland was caused by the hydrological condition. Variability between ecologies was confirmed by low values of genetic correlation coefficients. Each ecosystem is considered as an environment and the genetic correlation between all pairs of ecosystem combinations can be used to access genotype by environment interaction (Hammami et al., 2009). In the absence of genotype by environment interaction, the expected genetic correlation across environments is one (Hammami et al., 2009). Previous studies by Cooper and DeLacy (1994) reported that when the genetic correlation among environments is less than one, genotype by environment is significant. In this study genetic correlation coefficients were less than one for all variables and combinations.
Phenotypic frequency of O. glaberrima was analyzed along the toposequence. This study observed that most of the accessions showed best vigor from upland to lowland; vegetative vigor scores ranged between 1 (extra vigorous) and 5 (normal vigor); with other classifications being very low. Previous studies indicated that O. glaberrima cultivars achieve superior vegetative growth vigor (Asch et al., 1999; Dingkuhn et al., 1998, 1999; Johnson et al., 1998). This study also pinpointed that most of the O. glaberrima accessions were tall in hydromorphic and lowland conditions, which can be explained by the time spent by plants in water. Long time spending of plants in water increased plant height (Sie, 1991). Number of tillers at maturity indicated monomorphic frequency in 2009 and in 2010 from upland to lowland. This result confirms Sanni et al. (2008) studies that revealed monomorphic distribution of tillers number at maturity using plant material from different ecological zones. Most accessions tested were late or very late maturing in 2009 in upland (89.9%) and lowland (91.4%). Hydromorphic showed early maturity (38.1%) and intermediate maturity (30.9%). In 2009 long crop durations are probably due, in lowland to transplanting shock received by plants and to drought in upland (Dingkuhn and Asch, 1999; Dingkuhn et al., 1990). The variation observed among O. glaberrima tested was probably related to ecosystem. Manzelli et al. (2005) reported that local environment conditions influence genotypic constitution. They suggested a close relationship between agro-ecological conditions and morphological variation of germplasm. Diversity analysis of germplasm collections of several crop species and their wild relatives revealed considerable variability (Yang et al., 1991). High variability was obtained by Sanni et al. (2008) using O. sativa landraces from different ecological zones. The variability observed in O. glaberrima accessions along the toposequence was due to the ecological heterogeneity as reported by Sanni et al. (2008). Drought in August 2009 delayed flowering and crop duration period in the upland ecosystem with high percentage of very late and late maturing accessions. In 2010, most accessions were early and intermediate maturing and the percentage of late and very late maturing accessions were very low. This result can be explained by two factors: rainfall and late planting in 2010. Rainfall was 872.25 mm in 2009 and 1680.25 mm in 2010. No drought incidence was observed in 2010. Crops planted in May and July flowered at almost the same time. The high percentage of late maturing accessions was due to the early planting, which increased crop duration in 2009 (Dingkuhn and Asch, 1999). Crop duration determines the ability of the crop to avoid abiotic stress and related to the genotype and the environment Dingkuhn and Asch (1999). The maximal crop duration was observed for the May planting.
CONCLUSION
This study shows morphological and phenological variability of O. glaberrima along the toposequence. High variability is observed from upland to lowland ecologies and confirmed by variable phenotypic frequencies, high Shannon-Weaver index and low genetic correlation coefficients. Rainfall contributes to variability between ecosystems. Transplanting in lowland and drought in upland contribute to delayed crop phenology. Crop duration depends on the date of sowing; plants sown in May and July flowered at the same time. These findings will support efforts to breed rice varieties with short duration or phenological plasticity in different rice growing environments.
ACKNOWLEDGMENT
This study was financially supported by the Stress-tolerant Rice for Africa and South Asia Project funded by the Bill and Melinda Gates Foundation and the Ministry of Foreign Affair, Japan. The authors would like to acknowledge this support with deep thanks.
REFERENCES
- Abdi, A., E. Bekele, Z. Asfaw and A. Teshome, 2002. Patterns of morphological variation of sorghum (Sorghum bicolor (L.) Moench) landraces in qualitative characters in North Shewa and South Welo, Ethiopia. Hereditas, 137: 161-172.
CrossRef - Asch, F., A. Sow and M. Dingkuhn, 1999. Reserve mobilization, dry matter partitioning and specific leaf area in seedlings of African rice cultivars differing in early vigor. Field Crops Res., 62: 191-202.
CrossRef - Burdon, R.D., 1977. Genetic correlation as a concept for studying genotype-environment interaction in forest tree breeding. Silvae Genet., 26: 168-175.
Direct Link - Cooper, M. and I.H. DeLacy, 1994. Relationships among analytical methods used to study genotypic variation and genotype-by-environment interaction in plant breeding multi-environment experiments. Theor. Applied Genet., 88: 561-572.
CrossRef - Dingkuhn, M. and F. Asch, 1999. Phenological responses of Oryza sativa, O. glaberrima and inter-specific rice cultivars on a toposquence in West Africa. Euphytica, 110: 109-126.
CrossRefDirect Link - Dingkuhn, M., M.P. Jones, D.E. Johnson and A. Sow, 1998. Growth and yield potential of Oryza sativa and O. glaberrima upland rice cultivars and their interspecific progenies. Field Crops Res., 57: 57-69.
CrossRefDirect Link - Dingkuhn, M., D.E. Johnson, A. Sow and A.Y. Audebert, 1999. Relationships between upland rice canopy characteristics and weed competitiveness. Field Crops Res., 61: 79-95.
CrossRef - Dingkuhn, M., H.F. Schnier, S.K.D. Datta, E. Wijangco and K. Dorffling, 1990. Diurnal and developmental changes in canopy gas exchange in relation to growth in transplanted and direct-seeded flooded rice. Funct. Plant Biol., 17: 119-134.
CrossRefDirect Link - Futakuchi, K. and M. Sie, 2009. Better exploitation of African rice (Oryza glaberrima Steud.) in varietal development for resource-poor farmers in West and Central Africa. Agric J., 4: 96-102.
Direct Link - Hammami, H., B. Rekik and N. Gengler, 2009. Genotype by environment interaction in dairy cattle. Biotechnol. Agron. Soc. Environ., 13: 155-164.
Direct Link - Jain, S.K., C.O. Qualset, G.M. Bhatt and K.K. Wu, 1975. Geographical patterns of phenotypical diversity in a world collection of durum wheats. Crop Sci., 15: 700-704.
Direct Link - Johnson, D.E., M. Dingkuhn, M.P. Jones and M.C. Mahamane, 1998. The influence of rice plant type on the effect of weed competition on Oryza sativa and Oryza glaberrima. Weed Res., 38: 207-216.
CrossRefDirect Link - Jones, M.P., M. Dingkuhn, G.K. Aluko and M. Semon, 1997. Interspecific Oryza sativa L. X O. Glaberrima steud. Progenies in upland rice improvement. Euphytica, 94: 237-246.
CrossRefDirect Link - Kovach, M.J., M.T. Sweeney and S.R. McCouch, 2007. New insights into the history of rice domestication. Trends Genet., 23: 578-587.
CrossRef - Linares, O.F., 2002. African rice (Oryza glaberrima): History and future potential. Proc. Natl. Acad. Sci. USA., 99: 16360-16365.
CrossRefDirect Link - Manzelli, M., S. Benedettelli and V. Vecchio, 2005. Agricultural biodiversity in Northwest Somalia-an assessment among selected Somali sorghum (Sorghum bicolor (L.) Moench) germplasm. Biodivers. Conserv., 14: 3381-3392.
CrossRef - Oka, H.I., 1974. Experimental studies on the origin of cultivated rice. Genetics, 78: 475-486.
Direct Link - Saito, K., K. Azoma and J. Rodenburg, 2010. Plant characteristics associated with weed competitiveness of rice under upland and lowland conditions in West Africa. Field Crops Res., 116: 308-317.
CrossRef - Sanni, K.A., I. Fawole, R.G. Guei, D.K. Ojo and E.A. Somado et al., 2008. Geographical patterns of phenotypic diversity in Oryza sativa landraces of Cote d' Ivoire. Euphytica, 160: 389-400.
CrossRef - Sarla, N. and B.P.M. Swamy, 2005. Oryza glaberrima: A source for the improvement of Oryza sativa. Curr. Sci., 89: 955-963.
Direct Link - Semon, M., R. Nielsen, M.P. Jones and S.R. McCouch, 2005. The population structure of African cultivated rice Oryza glaberrima (Steud.): Evidence for elevated levels of linkage disequilibrium caused by admixture with O. sativa and ecological adaptation. Genetics, 169: 1639-1647.
CrossRefDirect Link - Yang, R.C., S. Jana and J.M. Clarke, 1991. Phenotypic diversity and associations of some potentially drought-responsive characters in durum wheat. Crop Sci., 31: 1484-1491.
Direct Link