
M.S. Ruzana Adibah
Department of Forest Management, Faculty of Forestry, Universiti Putra Malaysia, 43400, Serdang Selangor, Malaysia
A.N. Ainuddin
Institute of Tropical Forestry and Forest Product, Universiti Putra Malaysia, 43400, Serdang Selangor, Malaysia
Asian Journal of Plant Sciences
Year: 2011 | Volume: 10 | Issue: 2 | Page No.: 97-107







ABSTRACT
Epiphytes are plants susceptible to the current climate change due to continuous exposure of environmental changes. In this study, we review the epiphytes responses to fluctuations in their surrounding environments. Abiotic factors such as light and water are the important contributors towards the epiphytes growth. Epiphytes might suffer from environmental stresses namely high light intensity and water deficit, affecting its growth and physiological attributes. Epiphytes use several mechanisms to counter aforementioned problems and one of it is through changes of physiological pathways. Some of the epiphytes use Crassulacean Acid Metabolism (CAM) as protection system for survival in severe environments. Future studies should include more approaches used by this plant as defense mechanisms to such stresses and more studies on leaf anatomy, leaf structure and variations in biochemical components for further understanding of the mechanisms involved.
PDF Abstract XML References Citation
Received: March 07, 2011;
Accepted: May 04, 2011;
Published: June 10, 2011
How to cite this article
M.S. Ruzana Adibah and A.N. Ainuddin, 2011. Epiphytic Plants Responses to Light and Water Stress. Asian Journal of Plant Sciences, 10: 97-107.
DOI: 10.3923/ajps.2011.97.107
URL: https://scialert.net/abstract/?doi=ajps.2011.97.107
DOI: 10.3923/ajps.2011.97.107
URL: https://scialert.net/abstract/?doi=ajps.2011.97.107
INTRODUCTION
International community is widely concerned about climate change as one of the most critical environmental issues faced by mankind (Black et al., 2011). CO2 elevation and temperature rise has its impacts on plants’ growth, development and its physiological functions (Centritto, 2002) and the earth in general. Forest growth, survival and structure are some of the major concerns from climate change as an output from the interaction between forests and the environment (Chmura et al., 2011). An understanding of forest canopy is important where it operated as a medium for the energy, mass and momentum exchanges between environment and the ecosystem of the forest (Song et al., 2010). It contains myriads of plant populations and this includes the parasites, hemi-epiphytes, vascular and non-vascular epiphytes (Benzing, 1990; Nadkarni et al., 2004). In tropical forest, vascular epiphytes are included as a vital component of the forests biological diversity (Wester et al., 2011). Existence of epiphytes at plants and atmosphere interface made them vulnerable to climate change thus, other components of the forest consequently will almost directly affected from any negative impact occur to the epiphytes (Zotz and Bader, 2009). There are also increases in awareness to the view that the survival and continuation of the epiphytes community are gradually at risk (Nadkarni, 1992). Besides, excessive logging and land use to the tropical rain forests especially in the mountain rain forest are harmful and have a dangerous effect to the epiphytes community (Barthlott et al., 2001). Plants in this natural environment have higher risks to be affected by several damages in relation to environmental stresses (Huang, 2006). Moreover, epiphytes are subjected to multiple abiotic and biotic stresses of varying intensities and durations. The location of canopy dwelling plants like epiphytes make it sensitive to environmental stress (Oberbauer et al., 1996). Where, epiphytes are mainly habituated in a complex light atmosphere (Martin et al., 2001) where the main abiotic limitation in epiphytic habitat is the insufficiency of water (Zotz and Hietz, 2001).
When the environment of epiphytes is affected by its climate, automatically the plant’s physiological responses will be directly influenced. According to Schurr et al. (2006), at any state of environmental change, photosynthesis and growth processes are likely be affected. Climate change affects the overall functions and operations of physiological processes such as growth, transpiration and respiration (Brouder and Volenec, 2008).
In addition, the growth of plants is mainly influenced by genetic traits or its surroundings (Kramer and Kozlowski, 1979). In fact, plants will respond to environmental stresses through their physiological features such as modification organs’s growth, or by using various types of ways to prevail against the stresses (Schurr et al., 2006).
Therefore, it is necessary to discover how environmental factors especially light and water affect the epiphytes as they are mostly threatened by those two types of stresses. Their responses are vital to understand the epiphytes performances in fluctuating environments. This review will focus on the nature, contribution and response of epiphytic plants to light and water stress. Even though this paper touches on the entire globe, there is a clear focus on the growth and physiological responses of epiphytes to light and water stress. These observations also supplemented with reviews on epiphytic adaptive strategies through Crassulacean Acid Metabolism (CAM) in resistance against such stresses.
EPIPHYTES
Epiphytes are plants that settle on other plants and their habitats vary from the terrestrial understory to edge of the tree canopy (Zotz and Hietz, 2001). The epiphytic plants use other plants like trees, shrubs and woody vines as host. In contrast with the parasites, they are fully autotrophic (Benzing, 1998) or are considered as true epiphytes (Reinert, 1998). Ferns, orchids and bromeliads are embraces as epiphytes where they have characteristics that include thick and waxy leaves that allow them to require small amount of water (Ainuddin, 2007). There were plenty of epiphytic plants grew in wet tropical countries (Freiberg, 2001: Benzing, 1998, 1990). Besides, the vascular epiphytes are distinct characteristic element of tropical moist forests (Hietz et al., 2002). Additionally, epiphytes are globally known as dominant elements of the tropical rainforest (Fayle et al., 2009) and most of the vascular epiphytes are tropical plants (Zotz and Hietz, 2001). In addition, Benzing (1990) also mentioned that the epiphytes rarely exist in drier places. However, he stated that there are few types of epiphytes that could grow in the arid regions such as the orchids and the bromeliads where they are supported by plants like the Mexican and Peruvian cactus.
According to Diaz et al. (2010), administrative practices in Chilean temperate rainforests prominently overlook the epiphytic plants communities. They were removed due to misperception that they are the indicators of deteriorating tree health and reducing production of timber. In fact, epiphytes do act as a source of the tropical rainforest ecology where it contributes to many forests functions such as storing water from rainfall and holding nutrients (Benzing, 2004). In the process of hydrological and nutrient cycles of a forest system, epiphytes are the vital contributor (Benzing, 1998) since they engage and hold nutrients from the surrounding (Benzing, 1990).
Epiphytes play a significant function in influencing their nearby area microclimate particularly the canopy area (Stuntz et al., 2002). Moreover, according to Rosdi and Ainuddin (2004), microclimate of the surroundings area seems to be affected by the existence of plant. For example, the surrounding microclimatic state at the forest floor, does show a balance ecological system contained by much drier and warmer condition of the atmosphere, which is as the reflection of the contribution of canopy epiphytes and their accumulated dead organic matters (Freiberg, 2001; Freiberg and Turton, 2007). The vascular and non-vascular epiphytes contribute to the forest biomass. According to Nadkarni (1984), these epiphytic plants contribute to more than a few tons per hectare of biomass. In addition, these epiphytic plants also elevated the uptake of carbon and productivity of the forest (Diaz et al., 2010). For instance, a study of an epiphyte of Asplenium nidus shows that it could accumulate its dry mass up to one tones of per hectare (Ellwood et al., 2002).
In both vascular and non vascular plants there are epiphytisms among them and which comprise of pteridophytes, flowering plants, latter bryophytes, algae and lichens (Reinert, 1998). Generally, vascular epiphytic plants varieties are mostly localized in tropical regions (Dubuisson et al., 2009). Furthermore, Otto et al. (2009) reported that plenty of epiphytes that mainly ferns are absolutely acclimatized to the environment that is airy. According to Lorenzo et al. (2010), almost 80% species of the epiphytes fit into just four families: Bromeliaceae, Orchidaceae Polypodiaceae and Araceae.
PLANT STRESS
Climate change is the present issue that manipulates the ecological features of an ecosystem. The changes in the environment have huge effects especially on plants growth and its distribution. Interaction between plants and the environment will influence plants physiological functions thus affecting some ecological processes (Hegland et al., 2009). When the environment of a plant is affected by its climate, the natural processes of this plant will be directly influenced. Plant stress is always influenced by adverse environmental conditions, such as insufficient water supply, diseases, lack of nutrients or insect damage. There are many types of stresses caused by the changes of its surrounding area either in a minor or major way. These abiotic stresses will effect growth and in the long run, it will exterminate the survival of plant.
Canopy act as a filter to plant beneath it and reduce the solar energy received to the ground (Nuruddin and Tokiman, 2005). Environmental stresses for example water deficit and high light intensity might occur to epiphytes which live beneath the forest canopy. A species of Platycerium bifurcatum developed in diverse habitat which always exposed by drought and stress of high irradiance (Rut et al., 2008). The surrounding microclimate of the epiphyte area will therefore affect their mortality and as a result, it will affect the distribution of different species (Fayle et al., 2009). Epiphytes were also reported as the first population to be declined as their ecosystem was at risk and disturbed (Dubuisson et al., 2009).
However, epiphytes also exhibit ecophysiological, morphological and anatomical adaptation to survive under these harsh canopy conditions (Lorenzo et al., 2010). According to Suzuki et al. (2005), a complex system of transcription factors and other rigid genes will control the multiple defenses of enzymes, proteins and specific pathways if any plant is acclimatize to those abiotic stresses. Each plant will use different mechanism dealing with abiotic stress. For instance in Mimosa strigillosa, under environmental stress of soil pH, more growth is distributed to shoot under optimum condition, while under stress, it allow more growth to the roots (Nuruddin and Chang, 1999). This reflects that plant use different approach as an adjustment to environmental stress.
LIGHT STRESS
Light is one of the stress factors that have a huge influence on plant. The energy source for photosynthesis in plants is light and it is the vital requirement for plant life (Long et al., 1994). Light intensity and light quality fluctuation definitely have its effects on biochemical, physiological and plants developmental processes where under the low light level, growth and development are disturbed due to insufficient energy and under high light intensity, photodamaged may occur due to overload of the plant’s system (Humby and Durnford, 2006). Epiphytes can grow under different light condition ranging from the nearly full sun which is out in the open branches to the deep shade on base of the stem (Hietz and Briones, 2001). Epiphytes environment is always exposed to such stress and the regulation of plant growth and development are affected by the changes of the environmental conditions. Epiphytes that live under plant canopy are typically exposed to the elements of harsh atmosphere where light stress may arise among the epiphytic plants.
Light stress is when an excess light is absorbed greater than what is needed during the process of photosynthesis, where the excess of light is defined as when the ratio of Photon Flux Density (PFD) and photosynthesis is high (Demmig-Adams and Adams, 1992). High light intensity will lead to high temperature which will influence the plant growth due to hotter and drier surrounding microclimate. Different mechanisms shield the photosynthetic apparatus from over-energization under the overload light but, if the mechanisms of photoprotective of plant are insufficient, photoinhibition process will be turned on (He et al., 1996). The trigger to photoinhibition occurs due to the overexposure of plant to irradiance which it is higher than the irradiance usually obtained by plant (Stancato et al., 2002). The photochemical inactivation mostly of Photosystem II (PS II) is involved in photoinhibition (Sarvikas et al., 2006). Therefore, every organism that conducts photosynthesis is probably vulnerable to injury due to radiation influences, but the level of susceptibility depends on factors such as environmental, genotype, phenotype and physiological (Alves et al., 2002).
The growth of plants is closely related to photosynthesis and without light, photosynthesis will not occur and the growth of plants will be retarded. According to Winter et al. (1983), many CAM plants are not only could survive in the area with high light intensity but also under shaded condition. Capability of some species to survive under certain light range is probably dependent on how far they can adapt to new levels of light by changing their photosynthetic response (Heschel et al., 2004).
WATER STRESS
Meteorological term for drought is implied as restriction of water for a long period of time while water stress is likely to signify the complex progression of the effect of drought which is triggered by drought (Lombardini, 2006). Drought stress also has become more severe in some area due to the global climate change (Elsheery and Cao, 2008). Living in the canopy environments, water limitation is likely to occur among epiphytes. Extensive environmental conditions of drought and salinity converge to lower water accessibility of plants lead to the limitation of photosynthesis (Flexas et al., 2004), growth and worldwide yield (Chaves et al., 2003). The rate of CO2 assimilation of plants may be reduced under the condition of water insufficiency (Stancato et al., 2001). Closure of stomata and photosynthetic rates reduction are also stimulated by the limitation of water availability (Angelopoulos et al., 1996). For instance, in Mimosa strigillosa, the stomata are closed and leaves are folded to reduce loss of water (Chang et al., 1997). In addition, both stomatal closure and inhibition of leaf growth are among the earliest reactions towards drought to defend the plants from excessive water loss where it might lead to leaf cell dehydration and runaway xylem cavitations thus, bring towards the mortality (Chaves et al., 2003).
The main limitation of plant survival and growth is water stress and during acclimatization of plants to water stress, it involves several physiological and anti-oxidative apparatus (Upadhyaya et al., 2008). Therefore, stress tolerance indicates the capability of plants to survive in adverse environment through the adaptation and acclimatization from the state of stress (Lombardini, 2006). Epiphytes must tolerate the stress to survive in the harsh environment. Therefore, there were many defenses applicable to epiphytic plants (Benzing, 1990). For instance, the adjustment of conductance can defend a plant itself to drought or water stress. Moreover, Benzing (1990) also stated that osmotic adjustment and carbon dioxide fixation are other ways that can be used by plants to overcome this situation. According to Luvaha et al. (2008), in changing of environmental climate, it is beneficial for species to apply avoidance of drought mechanisms and adaptation through active osmoregulation. In addition, plants tolerate drought by avoiding tissue dryness and at the same time sustaining water potential or enduring the low water potential (Chaves et al., 2003). Generally, thick cuticles, succulence, sunken stomata and a thicker layer of boundary on the leaf surface are some of the adaptations applied by plants to conserve their water status (Hsu et al., 2006). For C4 or CAM plants, they use other methods of to cope with stress through carbon dioxide fixation for production of sugar with a minimum water loss (Xoconostle-Cazares et al., 2010). Water insufficiency in soil and plant tissue during drought leads to the adjustment in the processes of plant photosynthesis and has its effect on plant growth, development and survival in harsh environment (Lombardini, 2006).
GROWTH RESPONSE TO LIGHT AND
WATER STRESS
Growth is a mechanism achieved by division of cell, cell enlargement and differentiation and it is associated with the physiological, ecological, genetic and morphological measures and their complex interactions (Farooq et al., 2009). One of the important elements for plant growth and development is light (Saifuddin et al., 2010). Nearly all plants use solar radiation not only source of energy for photosynthesis, but also to regulate their processes of growth and development (Lombardini, 2006). Cervantes et al. (2005) theorized that individual epiphytes under extreme light intensity microhabitats within the canopy in a tropical dry forest of Yucatan, Mexico may have a restriction in their growth and reproduction. A study by Singh and Srivastava (1985) conformed to this finding where the fern of Azolla pinnata R. Brown recorded has the lowest value of mean leaf area in both of under the shade and highest light intensity treatments (Table 1).
In terms of biomass, unlike C3 plants that grow in high light area, plants that grown under shade would distribute more biomass to their photosynthetic tissues, creating thin, horizontally oriented foliages with little intra-canopy shadings thus, they have relatively higher concentrations of chlorophyll and higher coefficients of light absorption, however, these shaded plants have smaller value in root biomass indicating low rates of transpiration and low light saturated rates of photosynthesis (Skillman et al., 2005).
Decline in the vegetative growth of plants are one of the initial consequences of drought where water deficit will stimulate changes in terms of cell structure of plants and its metabolism (Khaled, 2010). Another consequence of water stress is turgor loss which reduces the size of cells leading to reductions in leaf expansion and shoot extension plus the leaf area reduction will definitely lessen the surface area for transpiration and thus, smaller leaves will decrease the light absorption and photosynthesis (Lombardini, 2006). In a study by Ainuddin and Nur Najwa (2009), water restriction lowered length and area of Asplenium nidus.
| Table 1: | Growth of fern and epiphytes species grown under different light treatment |
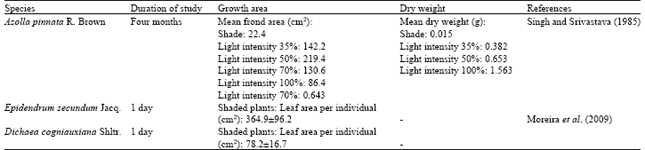 | |
According to Vurayai et al. (2011), decline in leaf area is the initial defense of plants to drought as in their study, water stress reduces area of the leaf to transpires less water. In addition, the reductions of leaf water potential and stomatal closure are the instantaneous reaction to water insufficiency, which point towards to decline in CO2 uptake and photosynthesis (Li et al., 2008). The reduction in photosynthesis results in a slower growth and lower plant biomass (Du et al., 2010) because plants need water to create biomass (Benzing, 1990). Furthermore, stomata closure and slower plant growth are desirable approaches to decrease further water loss (Sinclair and Purcell, 2005). However, only a few studies were done on the direct and long term observation as in concern of the growth of epiphytic plants (Hietz et al., 2002).
PHYSIOLOGICAL RESPONSE TO LIGHT AND WATER STRESS
Generally, the function of physiology of plant explains the growth of plants and its respond to the surrounding factors and cultural treatments (Kramer and Kozlowski, 1979). In extreme environments especially in high light intensity and water stress condition, plants will respond to changes through their physiological processes, such as the rate of photosynthesis, transpiration and stomatal conductance. In plant’s leaves, the factors from plant nutrition, light regime, leaf age, water stress and other physiological parameters can affect the photosynthetic CO2 assimilation (Von Caemmerer and Farquhar, 1981).
Although light is essential for photosynthesis, the light intensity whether it is low or high can affect plant growth (Valladares and Niinemets, 2008). Light intensity determines the degree of opening of the stomata and the guard cell, controlling water balance and influencing the photosynthesis rate of a plant via the light receptors that
drive fixation of CO2 and reduced intercellular CO2 concentration (Yu et al., 2004). In addition, the environment plays an important role in determining the photosynthesis efficiency, for example, plants that grow in a low light regime, their leaves absorb lower photon energy and they are depending on photon supplies for their photosynthesis rate (Miyake et al., 2009). Moreover, in a tropical dry forest of Yucatan, Mexico, plants that grow in the lower level of the canopy would have a lower photosynthetic rate while and plants that grow in the higher level of the canopy would experience photoinhibition (Cervantes et al., 2005). Roberts et al. (1998) found similar results where maximum rates of photosynthesis in shaded leaves are lower than the exposed ones while Schafer and Luttge (1988) found the opposite findings (Table 2). Epiphytes usually lived in a place that receives variability in light and is high in PFD since their photosynthetic apparatus will be affected, resulting in high evaporation rate which affects the plant water relations (Schafer and Luttge, 1988).
Moreover, physiological and biochemical processes of photosynthesis were affected by water stress (Ramanjulu et al., 1998). According to Chang et al. (1995), in adjustment to water stress condition, two physiological characteristics vital to plants are water transport system efficiency and regulatory system for water loss. In declining water availability, stomata closure has been noted as the earliest response of plants physiological attributes towards drought (Flexas and Medrano, 2002). Since there is known close correlation between stomatal conductance (gs) and net CO2 assimilation (Anet), it has been classified that in drought condition, stomatal closure has an influence in limiting the CO2 uptake in the leaves (Flexas et al., 2004). In the limitation of net photosynthesis rate, stomatal or non-stomatal factors were involved in this situation but the limitation was also dependable on the harshness and persistence of stress and also genetic reaction of the plant species (Ramanjulu et al., 1998). The survival capability of plantin severe drought is based on water loss limitation through minimum opening of stomata (Sanusan et al., 2010).
| Table 2: | Photosynthesis rate of epiphytes species grown under different light treatment |
 | |
Although the closure of stomata helps in sustaining high leaf water content and its water potential however is also responsible for the decline in photosynthesis (Ohashi et al., 2006). Moreover, in water deficit condition, the decline in CO2 assimilation rate is due to electron transfer in the Photosystem II was reduced, thus affecting the quantum yield in its photochemical apparatus (Stancato et al., 2001). In a study of drought tolerance in cereal species, it was found that the value of the quantum yield of PS II in water deficit condition was reduced with the increment in the stress level (Flagella et al., 1998).
Compared to other climatic elements in nature, light varies through its amplitude and radiation quantity and quality obtained by plants (Alves et al., 2002). When plants are subjected to strong light, their physiological features will respond. The declines in capability for photosynthesis were stimulated by exposing photosynthetic organisms, structures or organelles to visible light which has been denoted in various terms such as photoinhibition, photooxidation, photoinactivation, photolability, solarization and photodynamic reactions (Powles, 1984). Photoinhibition were used to explain the inhibition of photosynthetic capacity (Demmig-Adams and Adams, 1992) and the independence of gross adjustment in pigment concentration caused by excessive light (Powles, 1984). Long et al. (1994) also supported that photoinhibition was considered as light dependent and a gradual reduction of photosynthetic rate (Demmig-Adams and Adams, 1992) which was independent of any developmental change.
Furthermore, it is known that exposing green plants to excessive light has caused damages to photosynthetic apparatus (Powles, 1984). Moreover, tremendously too much light obtained by plants possibly will damage photosynthetic pigments (Powles, 1984) and the structure of plants which leading to photodamage (Larcher, 2003). In tropical region, high irradiance and high temperature arise concurrently with drought will direct the plants towards increasing their photon energy in chloroplast, which also decreases photochemical efficiency, thus leading to photosynthetic apparatus damages (Elsheery and Cao, 2008).
Chlorophyll fluorescence technique is a good indicator in determining the response from stresses. Whereby, in understanding adaptation system of plant and resistance to stress from the environment, chlorophyll fluorescence act as an assistant regards to this matter (Siam et al., 2008). This technique is handy and be used broadly by plant physiologists and ecophysiologists (Maxwell and Johnson, 2000). Chlorophyll fluorescence measuring techniques has currently improved and therefore is a vital instrument in the basic and applied physiology of plant (Krause and Weis, 1991). Using this technique, it calculates the modifications in photosystem II (PS II) action through the Chlorophyll a fluorescence changes which stimulated by the stress (Percival, 2005). There have been plenty of evidences associated with PS II acting as the main site of lesion in photoinhibition (Powles, 1984). The apparatus of photosynthesis in plants could possibly be affected provisionally by environmental stresses prior to the permanent morphological injury (Naumann et al., 2008; Percival and Sheriffs, 2002). Under various conditions and at various times, this technique of chlorophyll fluorescence measurement can estimate the parameters of the actual photosynthetic efficiency of the leaf (Maxwell and Johnson, 2000). It also measures the potential maximum of the quantum efficiency of Fv/Fm (Duraes et al., 2001).
Plant responses to stress conditions in this technique can be quantified through the measurement of fluorescence sign from dark adapting leaves of a plant for a certain period of time. As reported by Baker and Rosenqvist (2004), when the leaf was subjected to immediate light, fluorescence increases to a level of minimal fluorescence (Fo) which at this state the reaction centre of PS II.
CRASSULACEAN ACID METABOLISM: AS DEFENSE MECHANISM FROM STRESS FOR EPIPHYTES
Plant reacts to numerous stresses imposed by the climate change such as high and low concentrations of carbon dioxide, high light intensity and high temperature, which all have effects on carbon fixation and anatomical pathway (Holtum and Winter, 1999). There are three types of photosynthesis pathway, C3, C4 and Crassulacean Acid Metabolism (CAM) and many dynamic defense systems exist for epiphytics (Benzing, 1990). In fact, plants that grow in arid land modify their physiological metabolic system via CAM (Grams and Thiel, 2002). CAM is a CO2 concentrating mechanism that activates the Phosphoenolpyruvate Carboxylase (PEPC) enzyme at night for detaining the respiratory and atmospheric CO2. Physiologically, CAM preserves carbon and water in plants that grow in surroundings with limited accessibility those two resources either in short or long term basis (Borland and Taybi, 2004). In CAM photosynthesis, plant use a metabolic strategy in which during the cooler period at night, the stomata of the plants open to allow the nocturnal uptake of carbon dioxide (CO2) and during the day, stomata close to prevent the loss of water (Dodd et al., 2002; Rut et al., 2008). Thus, by this processes during day and night, it is considered that water stress could be adapted by CAM plants through the reduction of water loss (Luttge, 2004).
There have been many studies on the availability of this pathway on plants especially the epiphytes (Borland et al., 1998; Herrera et al., 2000; Hsu et al., 2006; Rut et al., 2008). A study by Holtum and Winter (1999) found that the tropical epiphytic and lithophytic ferns are the common plants that undergo CAM than it is presently investigated. In this study, eventhough the Polypodium crassifolium and Polypodium veitchii did not showed a strong CAM features, CAM activity still happened and was shown in the increase of titratable acidity. This shows that there is an occurrence of the CAM indirectly.
In a study by Wanek et al. (2002), there was an activity of CAM cycle in all stages of development of Clusia species which was the Clusia osaensis Hammel-ined., Clusia peninsulae Hammel-ined. and Clusia valerii Standl. This study shows that from the titratable protons and malic and citric acid there was a significant day-night flux cycle. Therefore, they advocate that the accessibility of water and light intensity created the appearance of CAM. Among epiphytes, the broad occasions of CAM are not expected as an outcome from short, but probably frequent phases of drought stress (Hsu et al., 2006). According to Rabas and Martin (2003), plants that are succulent will regularly uptake CO2 through CAM and additionally to the mechanism of C3.
Stresses from ecological factors could alter the isotope composition of C13 in many expected ways, ultimately via the effects on the balance among stomatal conductance and carboxylation (Robinson et al., 2000). Leavitt and Long (1982) reported that light intensity has an effect on the photosynthesis rate and in turn, it manipulates the composition of the carbon isotopic of plant and stresses from water could also influence stomata conductance and availability of water for photosynthesis. Epiphytes might survive via CAM where this mechanism allows plants to defense themselves through several stresses such as light and drought.
CONCLUSION
Climate changes have been manipulating the ecological value of an ecosystem. Light and water plays an important role in epiphytes growth and physiological performances. Many of epiphytes growth and physiological parameters are being affected, including the decline in biomass, photochemical efficiency and many more. Plants especially epiphytes have many ways in adapting to stressful conditions, which in such cases, CAM are vital for epiphytes in surviving harsh environment. The fixation of CO2 at night and the closing of stomata during day are the essential mechanisms for the epiphytes. Through these adjustments on their physiological functions, epiphytes certainly have a higher percentage in survivability by preventing themselves from dying water loss and increasing temperature. Other mechanisms encountered by plants to defense themselves from such stresses should be further investigated to get better idea of other stress responses in plants. In order to obtain a larger picture on the underlying explanation on what other parameters that are affected by light and water stress, things such as leaf anatomy, leaf structure and biochemical components variations in epiphytes need to be explored in the future.
ACKNOWLEDGMENT
We would like to thank every person involved in this project and we are also most grateful for each individual that give valuable suggestion and manuscript review. Financial support from the Research University Grant Scheme (RUGS) No. 03/01/07/0035RU are gratefully acknowledged.
REFERENCES
- Ainuddin, N.A. and D.A.N. Najwa, 2009. Growth and physiological responses of Asplenium nidus to water stress. Asian J. Plant Sci., 8: 447-450.
CrossRefDirect Link - Alves, P.L.C.A., A.C.N. Magalhaes and P.R. Barja, 2002. The phenomenon of photoinhibition of photosynthesis and its importance in reforestation. Bot. Rev., 68: 193-208.
CrossRef - Angelopoulos, K., B. Dichio and C. Xiloyannis, 1996. Inhibition of photosynthesis in olive trees (Olea europaea L.) during water stress and rewatering. J. Exp. Bot., 47: 1093-1100.
CrossRefDirect Link - Barthlott, W., V. Schmit-Neuerburg, J. Nieder and S. Engwald, 2001. Diversity and abundance of vascular epiphytes: A comparison of secondary vegetation and primary montane rain forest in the Venezuelan Andes. Plant Ecol., 152: 145-156.
Direct Link - Benzing, D.H., 1998. Vulnerabilities of tropical forests to climate change: The significance of resident epiphytes. Climatic Change, 39: 519-540.
CrossRef - Black , M.J., C. Whittaker, S.A. Hosseini, R. Diaz-Chaveza, J. Woods and R.J. Murphy, 2011. Life cycle assessment and sustainability methodologies for assessing industrial crops, processes and end products. Ind. Crops Prod.
CrossRef - Borland, A.M. and T. Taybi, 2004. Synchronization of metabolic processes in plants with Crassulacean acid metabolism. J. Exp. Bot., 55: 1255-1265.
CrossRef - Borland, A.M., L.I. Tecsi, R.C. Leegood and R.P. Walker, 1998. Inducibility of Crassulacean Acid Metabolism (CAM) in Clusia species; physiological/biochemical characterization and intercellular localization of carboxylation and decarboxylation processes in three species which exhibit different degrees of CAM. Planta, 205: 342-351.
CrossRef - Brouder, S.M. and J.J. Volenec, 2008. Impact of climate change on crop nutrient and water use efficiencies. Physiol. Plant, 133: 705-724.
CrossRef - Centritto, M., 2002. Interactive effects of climate change and water stress: implications for water-limited environments. First FAO-UCEA Technical Workshop of the Mediterranean Component of the Clim-Agri Project on Climate Change and Agriculture. Rome, Italy: FAO. http://www.fao.org/sd/climagrimed/pdf/ws01_33.pdf.
- Cervantes, S.E., E.A. Graham and J.L. Andrade, 2005. Light microhabitats, growth and photosynthesis of an epiphytic bromeliad in a tropical dry forest. Plant Ecol., 179: 107-118.
CrossRefDirect Link - Chang, M., C.M. Crowley and A.A. Nuruddin, 1995. Responses of herbaceous mimosa (Mimosa strigillosa), a new reclamation species, to cyclic moisture stress. Resources Conserv. Recycl., 13: 155-165.
CrossRefDirect Link - Chang, M., A.A. Nuruddin, C.M. Crowley and M.D. MacPeak, 1997. Evapotranspiration of herbaceous mimosa (Mimosa strigillosa), a new drought-resistant species in the southeastern United States. Resour. Conserv. Recycling, 21: 175-184.
CrossRef - Chaves, M.M., J.P. Maroco and J.S. Pereira, 2003. Understanding plant responses to drought-from genes to the whole plant. Funct. Plant Biol., 30: 239-264.
CrossRefDirect Link - Chmura, D.J., P.D. Anderson, G.T. Howe, C.A. Harrington and J.E. Halofsky et al., 2011. Forest responses to climate change in the northwestern United States: Ecophysiological foundations for adaptive management. For. Ecol. Manage., 261: 1121-1142.
CrossRef - Demmig-Adams, B. and WW. Adams III, 1992. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol., 43: 599-626.
CrossRefDirect Link - Diaz, I.A., K.E. Sieving, M.E. Pena-Foxon, J. Larrain and J.J. Armesto, 2010. Epiphyte diversity and biomass loads of canopy emergent trees in Chilean temperate rain forests: A neglected functional component. For. Ecol. Manage., 259: 1490-1501.
CrossRef - Dodd, A.N., A.M. Borland, R.P. Haslam, H. Griffiths and K. Maxwell, 2002. Crassulacean acid metabolism: Plastic fantastic. J. Exp. Bot., 53: 569-580.
CrossRef - Du, N., W. Guo, X. Zhang and R. Wang, 2010. Morphological and physiological responses of Vitex negundo L. var. heterophylla (Franch.) Rehd. to drought stress. Acta Physiol. Plant., 32: 839-848.
CrossRefDirect Link - Dubuisson, J.Y., H. Schneider and S. Hennequin, 2009. Epiphytism in ferns: Diversity and history. C R. Biol., 332: 120-128.
PubMed - Duraes, F.O.M., E.E.G. Gama, P.C. Magalhaes, I.E. Marriel and C.R. Casela et al., 2001. The usefulness of chlorophyll fluorescence in screening for disease resistance, water stress tolerance, aluminium toxicity tolerance and use efficiency in maize. Proceedings of the 17th Eastern and Southern Africa Regional Maize Conference, Feb. 11-15, CIMMYT, Mexico, DF, pp: 356-360.
- Ellwood, M.D.F., D.T. Jones and W.A. Foster, 2002. Canopy ferns in lowland dipterocarp forest support a prolific abundance of ants, termites and other invertebrates. Biotropica, 34: 575-583.
CrossRefDirect Link - Elsheery, N.I. and K.F. Cao, 2008. Gas exchange, chlorophyll fluorescence and osmotic adjustment in two mango cultivars under drought stress. Acta Physiol. Plant., 30: 769-777.
CrossRefDirect Link - Farooq, M., A. Wahid, N. Kobayashi, D. Fujita and S.M.A. Basra, 2009. Plant drought stress: Effects, mechanisms and management. Agron. Sustainable Dev., 29: 185-212.
CrossRefDirect Link - Fayle, T.M., A.Y.C. Chung, A.J. Dumbrell, P. Eggleton and W.A. Foster, 2009. The effect of rain forest canopy architecture on the distribution of epiphytic ferns (Asplenium spp.) in Sabah, Malaysia. Biotropica, 41: 676-681.
CrossRef - Flagella, Z., R.G. Campanile, M.C. Stoppelli, A. de Caro and N. di Fonzo, 1998. Drought tolerance of photosynthetic electron transport under CO2-enriched and normal air in cereal species. Physiol. Plant, 104: 753-759.
CrossRefDirect Link - Flexas, J. and H. Medrano, 2002. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot., 89: 183-189.
CrossRefPubMedDirect Link - Freiberg, M. and S.M. Turton, 2007. Importance of drought on the distribution of the birds nest fern, Asplenium nidus, in the canopy of a lowland tropical rainforest in North-Eastern Australia. Aust. Ecol., 32: 70-76.
CrossRef - Freiberg, M., 2001. The influence of epiphyte cover on branch temperature in a tropical tree. Plant Ecol., 153: 241-250.
CrossRef - Grams, T.E.E. and S. Thiel, 2002. High light-induced switch from C3-photosynthesis to Crassulacean acid metabolism is mediated by UV-A/blue light. J. Exp. Bot., 53: 1475-1483.
CrossRef - He, J., C.W. Chee and C.J. Goh, 1996. Photoinhibition of Heliconia under natural tropical conditions: The importance of leaf orientation for light interception and leaf temperature. Plant Cell. Environ., 19: 1238-1248.
CrossRef - Hegland, S.J., A. Nielsen, A. Lazaro, A.L. Bjerknes and O. Totland, 2009. How does climate warming affect plant-pollinator interactions? Ecol. Lett., 12: 184-195.
CrossRefDirect Link - Herrera, A., M.D. Fernandez and M.A. Taisma, 2000. Effects of drought on CAM and water relations in plants of Peperomia carnevalii. Ann. Bot., 86: 511-517.
CrossRef - Heschel, M.S., J.R. Stinchcombe, K.E. Holsinger and J. Schmitt, 2004. Natural selection on light response curve parameters in the herbaceous annual, Impatiens capensis. Oecologia, 139: 487-494.
CrossRef - Hietz, P. and O. Briones, 2001. Photosynthesis, chlorophyll fluorescence and within-canopy distribution of epiphytic ferns in a Mexican cloud forest. Plant Biol., 3: 279-287.
CrossRef - Hietz, P., J. Ausserer and G. Schindler, 2002. Growth, maturation and survival of epiphytic bromeliads in a Mexican humid montane forest. J. Trop. Ecol., 18: 177-191.
CrossRef - Holtum, J.A.M. and K. Winter, 1999. Degrees of crassulacean acid metabolism in tropical epiphytic and lithophytic ferns. Aust. J. Plant Physiol., 26: 749-757.
CrossRef - Hsu, C.C., T.C. Lin, W.L. Chiou, S.H. Lin, K.C. Lin and C.E. Martin, 2006. Canopy CO2 concentrations and crassulacean acid metabolism in Hoya carnosa in a subtropical rain forest in Taiwan: Consideration of CO2 availability and the evolution of CAM in epiphytes. Photosynthetica, 44: 130-135.
CrossRef - Khaled, S.A., 2010. Effect of watering regime on yield and its components of Triticum aestivum var. el-phateah L. Am. J. Plant Physiol., 5: 291-294.
Direct Link - Krause, G.H. and E. Weis, 1991. Chlorophyll fluorescence and photosynthesis: The basics. Ann. Rev. Plant Physiol. Plant Mol. Biol., 42: 313-349.
CrossRefDirect Link - Leavitt, S.W. and A. Long, 1982. Stable carbon isotopes as a potential supplemental tool in dendrochronology. Tree-Ring Bull., 42: 49-55.
Direct Link - Li, F., W. Bao, N. Wu and C. You, 2008. Growth, biomass partitioning and water-use efficiency of a leguminous shrub (Bauhinia faberi var. microphylla) in response to various water availabilities. New For., 36: 53-65.
CrossRef - Long, S.P., S. Humphries and P.G. Falkowski, 1994. Photoinhibition of photosynthesis in nature. Ann. Rev. Plant Physiol. Plant Mol. Biol., 45: 633-662.
CrossRef - Lorenzo, N., D.G. Mantuano and A. Mantovani, 2010. Comparative leaf ecophysiology and anatomy of seedlings, young and adult individuals of the epiphytic aroid Anthurium scandens (Aubl.) Engl. Environ. Exp. Bot., 68: 314-322.
CrossRefDirect Link - Luttge, U., 2004. Ecophysiology of crassulacean acid metabolism (CAM). Ann. Bot., 93: 629-652.
CrossRefDirect Link - Luvaha, E., G.W. Netondo and G. Ouma, 2008. Effect of water deficit on the physiological and morphological characteristics of mango (Mangifera indica) rootstock seedlings. Am. J. Plant Physiol., 3: 1-15.
CrossRefDirect Link - Martin, C.E., R. Hsu and T.C. Lin, 2001. Comparative photosynthetic capacity of abaxial and adaxial leaf sides as related to exposure in two epiphytic ferns in a subtropical rainforest in Northeastern Taiwan. Am. Fern J., 99: 145-154.
CrossRefDirect Link - Maxwell, K. and G.N. Johnson, 2000. Chlorophyll fluorescence-A practical guide. J. Exp. Bol., 51: 659-668.
CrossRefDirect Link - Miyake, C., K. Amako, N. Shiraishi and T. Sugimoto, 2009. Acclimation of tobacco leaves to high light intensity drives the plastoquinone oxidation system-relationship among the fraction of open PS11 centers, non-photochemical quenching of Chl fluorescence and the maximum quantum yield of PS11 in the dark. Plant Cell Physiol., 50: 730-743.
Direct Link - Moreira, A.S.F.P., J.P. de Lemos Filho, G. Zotz and R.M. dos Santos Isaias, 2009. Anatomy and photosynthetic parameters of roots and leaves of two shade-adapted orchids, Dichaea cogniauxiana Shltr. and Epidendrum secundum Jacq. Flora, 204: 604-611.
CrossRef - Nadkarni, N.M., 1984. Epiphyte biomass and nutrient capital of a neotropical elfin forest. Biotropica, 16: 249-256.
Direct Link - Nadkarni, N.M., 1992. The conservation of epiphytes and their habitats: Summary of a discussion at the international symposium on the biology and conservation of epiphytes. Selbyana, 13: 140-142.
Direct Link - Naumann, J.C., D.R. Young and J.E. Anderson, 2008. Leaf chlorophyll fluorescence, reflectance, and physiological responseto freshwater and saltwater flooding in the evergreen shrub, Myrica cerifera. Environ. Exp. Bot., 63: 402-409.
CrossRef - Nuruddin, A.A. and M. Chang, 1999. Responses of herbaceous mimosa (Mimosa strigillosa), a new reclamation species to soil pH. Conserv. Recycl., 27: 287-298.
CrossRef - Nuruddin, A.A. and L. Tokiman, 2005. Air and soil temperature characteristics of two sizes forest gap in tropical forest. Asian J. Plant Sci., 4: 144-148.
CrossRefDirect Link - Oberbauer, S.F., K. von Kleist, K.R.T. Whelan and S. Koptur, 1996. Effects of hurricane andrew on epiphyte communities within cypress domes of Everglades National Park. Ecology, 77: 964-967.
Direct Link - Ohashi, Y., N. Nakayama, H. Saneoka and K. Fujita, 2006. Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants. Biol. Plant., 50: 138-141.
CrossRefDirect Link - Otto, E.M., T. Janben, H.P. Kreier and H. Schneider, 2009. New insights into phylogeny of Pleopeltis and related Neotropical genera (Polypodiaceae, Polypodiopsida). Mol. Phylogenet. Evol., 53: 190-201.
CrossRef - Percival, G.C. and C.N. Sheriffs, 2002. Identification of drought-tolerant woody perennials using chlorophyll fluorescence. J. Arboricult., 28: 215-223.
Direct Link - Percival, G.C., 2005. The use of chlorophyll fluorescence to identify chemical and environmental stress in leaf tissue of three oak (Quercus) species. J. Arboric., 31: 215-227.
Direct Link - Powles, S.B., 1984. Photoinhibition of photosynthesis induced by visible light. Annu. Rev. Plant Physiol., 35: 15-44.
CrossRefDirect Link - Rabas, A.R. and C.E. Martin, 2003. Movement of water from old to young leaves in three species of succulents. Ann. Bot., 92: 529-536.
CrossRefDirect Link - Ramanjulu, S., N. Sreenivasalu, S.G. Kumar and C. Sudhakar, 1998. Photosinthetic characteristic in mulberry during water stress and rewatering. Photosynthetica, 35: 259-263.
CrossRefDirect Link - Roberts, A., A.M. Borland, K. Maxwell and H. Griffiths, 1998. Ecophysiology of the C3-CAM intermediate Clusia minor L. in Trinidad: Seasonal and short-term photosynthetic characteristics of sun and shade leaves. J. Exp. Bot., 49: 1563-1573.
CrossRefDirect Link - Robinson, D., L.L. Handley, C.M. Scrimgeour, D.C. Gordon, B.P. Froster and R.P. Ellis, 2000. Using stable isotope natural abundances (γ15 N and γ 13 C) to integrate the stress responses of wild barley (Hordeum spontaneum C. Koch.) genotypes. J. Exp. Biol., 51: 41-50.
CrossRefDirect Link - Rut, G., J. Krupa, Z. Miszalski, A. Rzepka and I. Slesak, 2008. Crassulacean acid metabolism in the epiphytic fern Platycerium bifurcatum. Photosynthetica, 46: 156-160.
CrossRefDirect Link - Sanusan, S., A. Polthanee, A. Audebert, S. Seripong and J.C. Mouret, 2010. Growth and yield of rice (Oryza sativa L.) as affected by cultivars, seeding depth and water deficits at vegetative stage. Asian J. Plant Sci., 9: 36-43.
CrossRefDirect Link - Saifuddin, M., A.M.B.S. Hossain and O. Normaniza, 2010. Impacts of shading on flower formation and longevity, leaf chlorophyll and growth of Bougainville glabra. Asian J. Plant Sci., 9: 20-27.
CrossRefDirect Link - Schafer, C. and U. Luttge, 1988. Effects of high irradiances on photosynthesis, growth and crassulacean acid metabolism in the epiphyte Kalanchoe unilora. Oecologia, 75: 567-574.
CrossRef - Schurr, U., A. Walter and U. Rascher, 2006. Functional dynamics of plant growth and photosynthesis-from steady-state to dynamics-from homogeneity to heterogeneity. Plant Cell Environ., 29: 340-352.
CrossRefDirect Link - Siam, A.M.J., K.M. Radoglou, B. Noitsakis and P. Smiris, 2008. Physiological and growth responses of three Mediterranean oak species to different water availability regimes. J. Arid Environ., 72: 583-592.
CrossRef - Sinclair, T.R. and L.C. Purcell, 2005. Is a physiological perspective relevant in a genocentric age. J. Exp. Bot., 56: 2777-2782.
CrossRefDirect Link - Singh, A. and O.N. Srivastava, 1985. Effect of light intensity on the growth of Azolla pinnata R. Brown at Ranchi, India. Hydrobiologia, 126: 49-52.
CrossRefDirect Link - Skillman, J.B., M. Garcia, A. Virgo and K. Winter, 2005. Growth irradiance effects on photosynthesis and growth in two co-occurring shade-tolerant neotropical perennials of contrasting photosynthetic pathways. Am. J. Bot., 92: 1811-1819.
Direct Link - Song, C., M.B. Dickinson, L. Su, S. Zhang and D. Yaussey, 2010. Estimating average tree crown size using spatial information from Ikonos and Quick Bird images: Across-sensor and across-site comparisons. Remote Sens. Environ., 114: 1099-1107.
CrossRef - Stancato, G.C., P. Mazzafera and M.S. Buckeridge, 2001. Effect of a drought period on the mobilization of non-structural carbohydrates, photosynthetic efficiency and water status in an epiphytic orchid. Plant Physiol. Biochem., 39: 1009-1016.
CrossRef - Stancato, G.C., P. Mazzafera and M.S. Buckeridge, 2002. Effects of light stress on the growth of the epiphytic orchid Cattleya forbesii lindl. X Laelia tenebrosa rolfe. Rev. Bras. Bot., 25: 229-235.
CrossRefDirect Link - Stuntz, S., U. Simon and G. Zotz, 2002. Rainforest air-conditioning: The moderating influence of epiphytes on the microclimate in tropical tree crowns. Int. J. Biometeorol., 46: 53-59.
CrossRefDirect Link - Suzuki, N., L. Rizhsky, H. Liang, J. Shuman, V. Shulaev and R. Mittler, 2005. Enhanced tolerance to environmental stress in transgenic plants expressing the transcriptional coactivator multiprotein bridging factor 1c. Plant Physiol., 139: 1313-1322.
CrossRefDirect Link - Upadhyaya, H., S.K. Panda and B.K. Dutta, 2008. Variation of physiological and antioxidative responses in tea cultivars subjected to elevated water stress followed by rehydration recovery. Acta Physiol. Plant., 30: 457-468.
CrossRefDirect Link - Valladares, F. and U. Niinemets, 2008. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst., 39: 237-257.
CrossRefDirect Link - Von Caemmerer, S. and G.D. Farquhar, 1981. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta, 153: 376-387.
CrossRefDirect Link - Vurayai, R., V. Emongor and B. Moseki, 2011. Effect of water stress imposed at different growth and development stages on morphological traits and yield of bambara groundnuts (Vigna subterranea L. Verdc). Am. J. Plant Physiol., 6: 17-27.
CrossRefDirect Link - Wanek, W., W. Huber, S.K. Arndt and M. Popp, 2002. Mode of photosynthesis during different life stages of hemiepiphytic Clusia species. Functional Plant Biol., 29: 725-732.
Direct Link - Wester, S., G. Mendieta-Leiva, L. Nauheimer, W. Wanek, H. Kreft and G. Zotz, 2011. Physiological diversity and biogeography of vascular epiphytes at Rio Changuinola, Panama. Flora, 206: 66-79.
CrossRef - Winter, K., B.J. Wallace, G.C. Stocker and Z. Roksandic, 1983. Crassulacean acid metabolism in Australian vascular epiphytes and some related species. Oecologia, 57: 129-141.
CrossRefDirect Link - Xoconostle-Cazares, B., F.A. Ramirez-Ortega, L. Flores-Elenes and R. Ruiz-Medrano, 2010. Drought tolerance in crop plants Am. J. Plant Physiol., 5: 241-256.
CrossRef - Yu, Q., Y. Zhang, Y. Liu and P. Shi, 2004. Simulation of the stomatal conductance of winter wheat in response to light, temperature and CO2 changes. Ann. Bot., 93: 435-441.
CrossRefDirect Link - Zotz, G. and P. Hietz, 2001. The physiological ecology of vascular epiphytes: Current knowledge, open questions. J. Exp. Bot., 62: 2067-2078.
CrossRefDirect Link - Zotz, G. and M.Y. Bader, 2009. Epiphytic plants in a changing world-global: Change effects on vascular and non-vascular epiphytes. Prog. Bot., 70: 147-170.
CrossRef - Baker, N.R. and E. Rosenqvist, 2004. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot., 55: 1607-1621.
CrossRefDirect Link - Flexas, J., J. Bota, F. Loreto, G. Cornic and T.D. Sharkey, 2004. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol., 6: 269-279.
CrossRefDirect Link