
Saubhik Das
Department of Botany, Taki Government College, Taki, North 24-Parganas, Pin-743429, West Bengal, India
LiveDNA: 91.3693
Asian Journal of Plant Sciences
Year: 2011 | Volume: 10 | Issue: 4 | Page No.: 263-268







ABSTRACT
Taxonomic interpretations are based on some evidences. In biosystematics along with conventional morphological parameters, now-a-days molecular markers are extensively used in solving taxonomic disputes. There may be congruence or conflict between these two approaches. In the present study with twelve species of Ipomoea, general plant morphological character states as well as RAPD fragment phenotypes were employed to reflect interrelationship and clustering pattern of the species. Species pairs showing significantly greater pairing affinity values in morphological analysis also revealed higher monomorphism in RAPD band profile. Among the species I. hispida showed least number of amplified fragments (73) whereas, I. aquatica revealed highest number of amplified fragments (213). Dendrograms computed from morphological and RAPD data showed definite clustering pattern of Ipomoea species and significantly alike relative closeness. Present study revealed a sharp congruence between morphological and molecular approach.
PDF Abstract XML References Citation
Received: June 16, 2011;
Accepted: July 26, 2011;
Published: October 31, 2011
How to cite this article
Saubhik Das, 2011. Congruence Between Morphological and Molecular Approach in Understanding Species Relationship in Ipomoea spp.: A Rare Event in Taxonomy. Asian Journal of Plant Sciences, 10: 263-268.
DOI: 10.3923/ajps.2011.263.268
URL: https://scialert.net/abstract/?doi=ajps.2011.263.268
DOI: 10.3923/ajps.2011.263.268
URL: https://scialert.net/abstract/?doi=ajps.2011.263.268
INTRODUCTION
Ipomoea is the largest genus of the family Convolvulaceae comprising 650 species distributed all over the world (Mabberley, 1997). The genus occurs throughout the tropical and subtropical regions of the globe (Miller et al., 2004) and comprises annual and perennial herbs, shrubs and even small trees. Most of the species are twining, climbing plants.
Taxonomic interpretations are based on some evidences. Morphological features are age-old parameters but molecular techniques have earned ample importance to solve the taxonomic disputes. Randomly Amplified polymorphic DNA markers (Williams et al., 1990) have been used by several workers to evaluate species relationship, also to maintain clonal cultivars. RAPD markers have been utilized to study inter and intrageneric diversity of genotypes from two Jute species (Ogunkanmi et al., 2010), for identification of mutant tomato (Kulkarni and Deshpande, 2010). RAPD molecular marker proved to be superior over morphological approach to characterize large number of landrace rice accessions (Ogunbayo et al., 2007). RAPD profile was used to analyse the genetic diversity and distance between clonal cultivars of Ipomoea batatas L. (Connolly et al., 1994; Zhang et al., 1998). Jarret and Austin (1994) proved the utility of RAPD marker for evaluating genetic diversity in sweet potato and for establishing taxonomic as well as evolutionary relationship in Ipomoea species. Phylogeny of sweet potato and its wild species was analysed involving morphological variation, crossing ability and RAPD pattern of sweet potato and its closely related species (Katsumi, 2001). Morphometrics and quantitative characteristics of mature cotyledon (Ogunwenmo, 2003) and morphological variations both qualitative and quantitative (Mondal et al., 2006) were utilised for identification of Ipomoea taxa. Morphological and molecular parameters when used simultaneously to study species relationship or infra-specific diversity analysis showed conflicting interpretation in most of the cases (El-Shazly and El-Mutairi, 2006; Paul et al., 2010).
In the present investigation, the morphological character states as well as RAPD fragment phenotype were employed. Primary goal was to study relationship between different species of Ipomoea. Secondary aim was to explore congruence if any between the interpretations based on morphological and molecular approach. The later is a rare event in taxonomy especially when different unrelated species are engaged in the study.
MATERIALS AND METHODS
Materials: Twelve species of Ipomoea collected from different parts of lower Gangetic plain of West Bengal, India, included nine wild species – I. hispida Roem and Schult., I. hederifolia L., I. pes-caprae (L.) Sweet., I. fistulosa Mart ex Choisy., I. sepiaria Koen., I. obscura (L.) Ker-Gawl., I. pes-tigridis L. I. chryseides Ker-Gawl., I. triloba (L.) Roth., two horticultural species-I. quamoclit L., I. nil (L.) Roth. and one semi cultivated species I. aquatica Forsk. Seeds of all the species were collected from different locale of West Bengal and were grown in experimental farm of Bose Research Institute, Kolkata, for consecutive three years for morphological and various bimolecular characterizations.
Morphological study: For plant morphological study 64 characters with character states including habit and habitat were taken into consideration. Measurements were taken from 10 readings for each character state. Few prominent character shared by all taxa were also considered for analysis.
RAPD analysis: Plant DNA was extracted from young seedlings by CTAB method (Rogers and Bendich, 1988). The DNA concentration in each sample was adjusted to 25 ng μL-1 for PCR.
PCR amplifications were performed according to the method of Williams et al. (1990) using DNA amplification kit from GENEI Bangalore, India, a set of 12 oligonucleotide primer, OPA 01–OPA 12 (Table 1) from Operon technologies, Inc, Alameda, Calif.) and Perkin Elmer Cetus thermal cycler. Temperature profile of each cycle was 40 sec. for denaturation at 94°C, 1 min. for Annealing at 35°C and 90 sec for extension at 72°C. Reaction continued for 40 cycles followed by 7.5 min. Hold at 72°C to ensure that extension reactions completed. PCR reaction mixture of 25 μL comprised 1X buffer, 0.2 mM of dATP, dCTP, dTTP, dGTP, 2 mM MgCl2, 0.2 μL of primer, 100 ng of template DNA and U of Taq DNA Polymerase. PCR reactions were started ‘hot’ i.e., template DNA in 20 μL reaction mixture without Taq Polymerase was denatured first at 94°C for 5 min. followed by addition of 1 U of Taq polymerase at 72°C. The amplification products were electrophoresed with φ X 174 Hae III digested DNA (GENEI) as DNA size marker.
Statistical analysis: In case of morphological analysis on the basis of presence or absence of a particular character state in all the species, a similarity matrix was prepared giving each character a definite number. Pairing Affinity (PA) values between different combinations of species pair were calculated as follows:
In case of RAPD fragment polymorphism analysis pairing affinity values were calculated as follows:
| Table 1: | Random Oligonucleotide primers used in RAPD fragment polymorphism analysis |
 | |
Using pairing affinity values, Dendrograms on morphology and RAPD fragment polymorphism were computed following Unweighted Pair Group Method with Average (UPGMA) method.
RESULTS
Morphological analysis: Most of the species were annual excepting perennial species like- I. fistulosa, I. aquatica and I. pes-caprae. Species were mostly terrestrial twining herbs excepting I. fistulosa, a terrestrial erect shrub., I. aquatica a semi-aquatic prostrate herb and I. pes-caprae, a terrestrial prostrate herb. Flower colour ranged from white to purple. Purple colour appeared to be the most prevalent colour with exception in I. chryseides (yellow), I. nil (blue).
The morphological features of uniform appearance in all the species were- simple exstipulate leaves; sepals 5, polysepalous, persistent; petals 5 gamopetalous, petals with 5 prominent bands; stamens 5, epipetalous, filaments unequal with hairy outgrowth at base; fruit valved, dehiscent capsule.
Percentage based pairing affinity values for different combination of species pair of Ipomoea were calculated from the character state distribution pattern. The values ranged from 45.3% (I. fistulosa and I. nil) to 81.25% (I. obscura and I. triloba) (Table 2). Significantly higher level of pairing affinity values (70% or above) were shown by species pair like I. quamoclit and I. hederifolia (70.68%), I. quamoclit and I. chryseides (71.87%), I. quamoclit and I. triloba (73.43%), I. hederifolia and I. obscura (73.43%). I. hederifolia and I. triloba (71.87%), I. pes-caprae and I. aquatic (73.43%), I. aquatic and I. sepiaria (75%) I. sepiaria and I. obscura (70.31%), lastly I. sepiaria and I. triloba (75%).
Dendrogram computed from pairing affinity values revealed three major clusters or groups at specific similarity level or similarity index.
| Table 2: | Pairing affinity values between different combinations of species pairs of Ipomoea on morphological features |
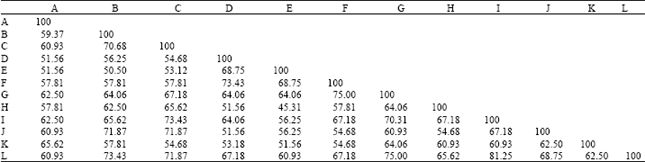 | |
| Species of Ipomoea, A: I. hispida, B: I. quamoclit, C: I. hederifolia, D: I. pes-caprae, E: I. fistulosa, F: I. aquatica, G: I. sepiaria, H: I. nil, I: I. obscura, J: I. chryseides, K: I. pes-tigridis, L: I. triloba | |
| Table 3: | RAPD fragments amplified in different Ipomoea spp. |
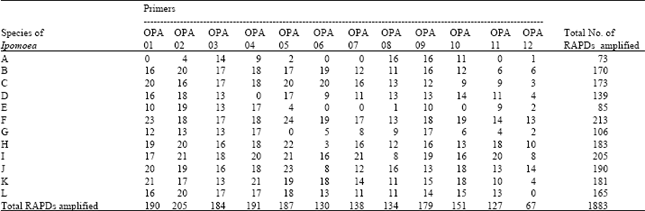 | |
| Species of Ipomoea, A: I. hispida, B: I. quamoclit, C: I. hederifolia, D: I. pes-caprae, E: I. fistulosa, F: I. aquatica, G: I. sepiaria, H: I. nil, I: I. obscura, J: I. chryseides, K: I. pes-tigridis, L: I. triloba | |
Group-A comprised I. fistulosa, I. pes-caprae and I. aquatica, Group- B included I. sepiaria, I. obscura and I. triloba and Group-C comprised I. nil, I. quamoclit, I. hederifolia, I. chryseides and I. pes-tigridis and I. hispida remained isolated from other members (Fig. 1).
RAPD polymorphism analysis: In RAPD fragment polymorphism analysis out of 12 primers used only four viz., OPA 02, 03, 08 and 09 revealed amplification for all the species. Total number of amplified fragments were 1877 in all the species from 12 primers. Only primer OPA-08 showed one monomorphic band (i.e., band having similar migrating pattern present in all the species (Fig. 2D). Other primers produced amplified fragments not universally present i.e., were polymorphic in respect of twelve species. Amplified products obtained with different primers are given in Table 3. Among the species investigated I. hispida showed least number of amplified fragments (73) whereas I. aquatica revealed highest number of amplified fragments (213). Primer OPA-02 (Fig. 2B) produced highest number of amplified products (205) whereas primer OPA -12 produced least number of amplified fragments (63) among the species (Table 3).
 | |
| Fig. 1: | Dendrogram computed from morphological features of Ipomoea spp. Showing cluster pattern |
RAPD fragment profile of a species was treated as fragment phenotype and each band was considered as potential variable.
| Table 4: | Pairing affinity values between different combinations of species pairs of Ipomoea on RAPD fragment profile |
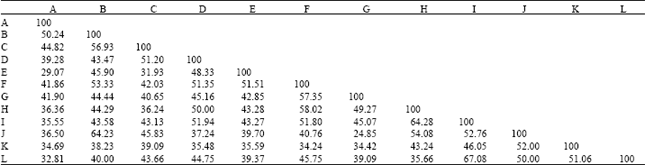 | |
| Species of Ipomoea, A: I. hispida, B: I. quamoclit, C: I. hederifolia, D: I. pes-caprae, E: I. fistulosa, F: I. aquatica, G: I. sepiaria, H: I. nil, I: I. obscura, J: I. chryseides, K: I. pes-tigridis, L: I. triloba | |
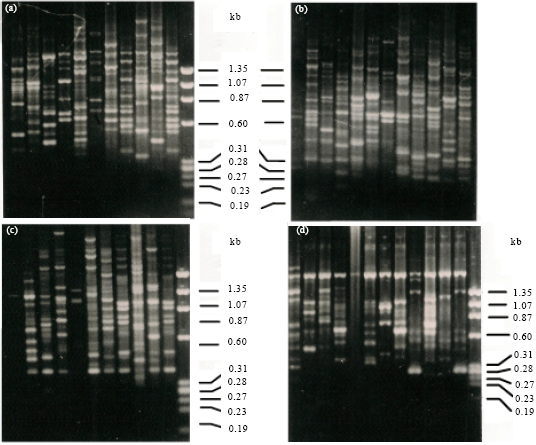 | |
| Fig. 2 (a-d): | RAPD fragment polymorphism in Ipomoea spp., (a) With primer OPA 01, Lane 1-11 1 I. quamoclit 2. I. hederifolia 3. I. pes-caprae, 4. I. fistulosa 5. I. aquatica 6. I. sepiaria, 7. I. nil 8. I. obscura 9. I. chryseides, 10. I. pes-tigridis. 11. I. triloba M. DNA size marker, (b) With primer OPA 02, Lane 1-12 1. I. hispida 2. I. quamoclit 3. I. hederifolia, 4. I. pes-caprae. 5. I. fistulosa 6. I. aquatic, 7. I. sepiaria 8. I. nil 9. I. obscura, 10. I. chryseides 11. I. pes-tigridis. 12. I. triloba, M. DNA size marker, (c) With primer OPA 05, Lane 1-11 1. I. hispida 2. I. quamoclit 3. I. hederifolia, 4. I. fistulosa 5. I. aquatica 6. I. sepiaria, 7. I. nil 8. I. obscura 9. I. chryseides, 10. I. pes-tigridis. 11. I. triloba M. DNA size marker, (d) With primer OPA 08, Lane 1-12 1. I. hispida 2. I. quamoclit 3. I. hederifolia, 4. I. pes-caprae. 5. I. fistulosa 6. I. aquatic, 7. I. sepiaria 8. I. nil 9. I. obscura, 10. I. chryseides 11. I. pes-tigridis. 12. I. triloba, M. DNA size marker |
Pairing affinity values were calculated between different combinations of species pair based on monomorphic band profile (Table 4). Pairing affinity values in one way reflected percentage based monomorphism in different combinations of species pair, in reverse way the polymorphism percentage.
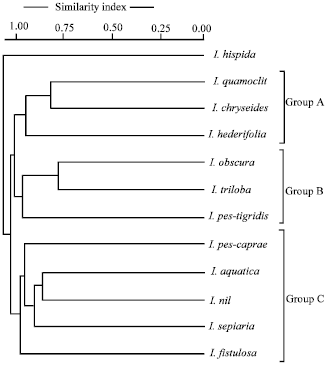 | |
| Fig. 3: | Dendrogram computed from RAPD fragment polymorphism in Ipomoea spp. |
Monomorphism percentage ranged from 24.85 to 67.08%. species pair I. sepiaria and I. chryseides showed least monomorphism of 24.85% i.e., greatest polymorphism and species pair I. obscura and I. triloba showed greatest monomorphism of 67.08% i.e., least polymorphism.
Dendrogram computed from monomorphism percentage showed distinct cluster or Groups (Fig. 3). The Dendrogram showed three major groups, Group-A comprised species- I. quamoclit, I. chryseides and I. hederifolia, Group-B comprised species – I. obscura, I. triloba and I. pes-tigridis and Group-C included species I. pes-caprae, I. aquatic, I. nil, I. sepiaria, I. fistulosa. I. hispida remained isolated from other species.
DISCUSSION
In taxonomic research along with the conventional morphological parameters, molecular markers are extensively used in solving taxonomic disputes. Same interpretations may or may not be drawn using both the parameters in respect of a particular taxonomic problem. Most of the previous taxonomic and systematic works on Ipomoea focussed the economically important cultivated hexaploid sweet potato (I. batatas L.), their origin, evolution and relationship with putative progenitor. I. trifida was appeared as a probable progenitor of I. batatas by cytogenetical and molecular studies (Srisuwan et al., 2006). More or less similar interpretation was drawn previously from combined ISSR and cpDNA dataset (Huang and Sun, 2000). There are large number of weed species interrelationship of which are least investigated.
RAPD phenotypes are inherited in a dominant fashion, therefore do not allow direct estimation of heterozygosity (Tingey and del Tufo, 1993). Prime goal of the present investigation was to group the species according to their relative closeness using morphological character states and RAPD fragment phenotype.
Numerical taxonomic techniques (Harman, 1976; Sneath and Sokal, 1973) have been proved to be a useful means to trace out genetic variability and species relationship (Elisens and Crawford, 1988). It is assumed that there exist a set of common characteristics or variables associated with each unit. The closeness between the two units is formulated in terms of a function and is called the distance.
Cluster pattern in the Dendrogram on morphology revealed clear grouping among the species. The relationship between the species of Group-A (I. fistulosa, I. pes-caprae and I. aquatica) was supported by erect or prostrate habit, perennial nature, large purple flower and big seeds. Relationship between members of Group-B (I. sepiaria, I. obscura and I. triloba) and Group-C (I. nil, I. quamoclit I. hederifolia, I. chryseides and I. pes-tigridis) is evidenced by twining annual habit, ovate-cordate leaves with acute to acuminate tip and small seeds. I. hispida is quite separable from other species in having hispid habit, very small white or purplish flower and very small seeds. The evolved members of the family Convolvulaceae are presumed to be twining plants with generally ovate-cordate leaves and showy white or purplish flower (Rendle, 1983), accordingly Group-B and C comprising comparatively evolved members. Large seeds are generally associated with woody perennials that are considered to be primitive (Salisbury, 1942) and accordingly Group-A including comparatively primitive members.
RAPD fragment obtained with different primers showed a high degree of polymorphism. Clustering pattern and relative closeness of the species was significantly alike. Dendrogram on RAPD showed complete isolation of I. hispida from other species as in morphological Dendrogram. In both the Dendrograms I. obscura and I. triloba were in close proximity. I. quamoclit I. hederifolia and I. chryseides were clustered in well defined cluster in both the occasions. Ipomoea represent one of the few cases where we can find a significant congruence between morphological and molecular approach especially when well defined separate species are considered. The minor deviations and discrepancies might have been minimized or eliminated if large number of RAPD primers were used.
ACKNOWLEDGMENT
Author expresses his sincere gratitude to Prof. Sampa Das and Dr. Debabrata Basu of Department of Plant Science, Bose institute, Kolkata, for their advice and assistance.
REFERENCES
- Connolly, A.G., I.D. Godwin, M. Cooper and I.H. DeLacy, 1994. Interpretation of randomly amplified polymorphic DNA marker data for fingerprinting sweet potato (Ipomoea batatas L.) genotypes. Theor. Applied Genet., 88: 332-336.
CrossRefDirect Link - El-Shazly, H.H. and Z. El-Mutairi, 2006. Genetic relationships of some barley cultivars, based on morphological criteria and RAPD fingerprinting. Int. J. Bot., 2: 252-260.
CrossRefDirect Link - Huang, J.C. and M. Sun, 2000. Genetic diversity and relationships of sweet potato and its wild relatives in Ipomoea series Batatas (Convolvulaceae) as revealed by Inter-Simple Sequence Repeat (ISSR) and restriction analysis of chloroplast DNA. Theor. Appl. Genet., 100: 1050-1060.
CrossRefDirect Link - Jarret, R.I. and D.F. Austin, 1994. Genetic diversity and systematic relationship in sweet potato (Ipomoea batatas (L.) Lam) and related species as revealed by RAPD analysis. Genet. Resour. Crop Evol., 41: 165-173.
CrossRefDirect Link - Katsumi, K., 2001. Phylogeny of Ipomoea species closely related to sweet potato and their breeding use. Bull. Natl. Inst. Crop Sci., 1: 1-56.
Direct Link - Kulkarni, M. and U. Deshpande, 2010. RAPD based fingerprinting of tomato genotypes for identification of mutant and wild cherry specific markers. J. Plant Sci., 5: 273-281.
Direct Link - Mondal, A., G. Kabir, G.P. Ghosh, N. Yasmin, A.M.S. Alam and H.A. Khatun, 2006. Morphological variation of ten Ipomoea species of Mangladesh. Pak. J. Biol. Sci., 9: 1714-1719.
CrossRef - Ogunbayo, S.A., D.K. Ojo, A.R. Popoola, O.J. Ariyo and M. Sie et al., 2007. Genetic comparisons of landrace rice accessions by morphological and RAPDs techniques. Asian J. Plant Sci., 6: 653-666.
Direct Link - Ogunwenmo, K.O., 2003. Cotyledon morphology: An aid in identification of Ipomoea taxa (Convolvulaceae). Feddes Repertorium, 114: 198-203.
CrossRefDirect Link - Paul, A., S. Bandyopadhyay, P. Acharyya and S.S. Raychaudhuri, 2010. Studies on genetic diversity of twelve accessions of Momordica charantia L. using morphological, RAPD and SCAR markers. Asian J. Plant Sci., 9: 471-478.
CrossRefDirect Link - Srisuwan, S., D. Sihachakr and S. Siljak-Yakovlev, 2006. The origin and evolution of sweet potato (Ipomoea batatas Lam.) and its wild relatives through the cytogenetic approaches. Plant Sci., 171: 424-433.
CrossRef - Tingey, S.V. and J.P. del Tufo, 1993. Genetic analysis with random amplified polymorphic DNA markers. Plant Physiol., 101: 349-352.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Zhang, D., M. Ghislain, Z. Huaman, A. Golmirzaie and R. Hijman, 1998. RAPD variation in sweet potato (Ipomoea batatas (L.) Lam.) cultivars from South America and Papua New Guinea. Genet. Resour. Crop Evol., 45: 271-277.
Direct Link - Ogunkanmi, L.A., W.O. Okunowo, O.O. Oyelakin, B.O. Oboh, O.O. Adesina, K.O. Adekoya and O.T. Ogundipe, 2010. Assessment of genetic relationships between two species of jute plants using phenotypic and RAPD markers. Int. J. Bot., 6: 107-111.
CrossRefDirect Link