
Ramesh Chandra
Biotechnology Lab, Central Institute for Subtropical Horticulture, Rehmankhera, Lucknow-227107, India
Madhu Kamle
Biotechnology Lab, Central Institute for Subtropical Horticulture, Rehmankhera, Lucknow-227107, India
Anju Bajpai
Biotechnology Lab, Central Institute for Subtropical Horticulture, Rehmankhera, Lucknow-227107, India
M. Muthukumar
Biotechnology Lab, Central Institute for Subtropical Horticulture, Rehmankhera, Lucknow-227107, India
Shahina Kalim
Department of Biochemistry, Bundelkhand University, Jhansi-284128, India
Asian Journal of Plant Sciences
Year: 2010 | Volume: 9 | Issue: 8 | Page No.: 437-446







ABSTRACT
In vitro selection for disease resistance in fruit crops is a biological phenomenon involving interaction between host-pathogen and additive effects of abiotic factors favoring the disease development. Among several strategies developed for disease resistance breeding, which are long term methods especially for perennials, a comparatively simpler and easier method is tissue culture based technique of selection of somaclonal variants against specific pathogen or phytotoxin. In vitro selection of plantlets regenerated in a culture media amended or supplemented with varying concentration of the culture filtrate or fractionated toxin of the specific pathogen serves as an efficient technique because of easy screening and reducing breeding cycle for developing disease resistance lines. In the past few decades, several researchers have worked out the possibility of obtaining somaclonal variants resistant to specific pathogenic race causing disease. Hence, in vitro selection strategy is a candidate alternative for conventional breeding approach for disease resistance. This review study gives an overview on prospects of in vitro selection for disease resistance with special reference to fruit crops.
PDF Abstract XML References Citation
Received: August 06, 2010;
Accepted: October 05, 2010;
Published: November 25, 2010
How to cite this article
Ramesh Chandra, Madhu Kamle, Anju Bajpai, M. Muthukumar and Shahina Kalim, 2010. In vitro Selection: A Candidate Approach for Disease Resistance Breeding in Fruit Crops. Asian Journal of Plant Sciences, 9: 437-446.
DOI: 10.3923/ajps.2010.437.446
URL: https://scialert.net/abstract/?doi=ajps.2010.437.446
DOI: 10.3923/ajps.2010.437.446
URL: https://scialert.net/abstract/?doi=ajps.2010.437.446
INTRODUCTION
Plant tissue culture is fundamental to our understanding of the control of plant function and the modification to meet the needs of modern agriculture, horticulture and forestry (Matsumoto et al., 1995; Jayasankar et al., 2000). Through tissue cultures, it is now possible to examine the selection of callus cultures, somaclonal variations (Skirvin, 1978; Larkin and Scowcroft, 1981) host-pathogen interactions, disease resistance and germplasm with enhanced disease resistance (Hammerschlag, 1988; Remotti et al., 1997). Selection for disease resistance breeding in fruit crops is an alternative to genetic engineering (Jayasankar and Gray, 2003; Chandra et al., 2004; Chandra and Mishra, 2007). It relates to the application of agents (culture filtrate, toxin fraction etc.) in regeneration system that results in lines with increased variability in resistance or susceptibility (Svabova and Lebeda, 2005). Diseases are caused by microorganisms producing toxins and metabolites which could be involved in the pathogenicity (the ability to cause disease), host-pathogen interactions and in disease expression (Wolpert et al., 1994). Phytotoxic metabolites of most of the pathogens have been reported to play a significant role in pathogenesis (Chandrasekharan and Ramakrishnan, 1973; Walker and Templeton, 1978; Amusa, 1991; Amusa et al., 1993) and these have been employed in screening crops for improved lines against disease resistance (Wheeler and Luke, 1955; Vidyasekaran et al., 1990; Amusa, 1994, 1998, 2000). Typical symptoms of most plant diseases revealed that the involvement of phytotoxic metabolites, suggesting their secretion by the pathogen in the disease development. Metabolites of many fungi may have adverse or stimulatory effects on plants (Heisey et al., 1985; Rice, 1995) such as suppression of seed germination, malformation and retardation of seedling growth (Lynch and Clark, 1984). The methods for selection of disease resistance in fruit trees have been reviewed by Hammerschlag (1983, 1984) and Hwang (1990). The principle behind this approach is that the metabolites present in the culture filtrate serve as selection agent for evolving resistant lines.
In vitro selection offers immense potential for quick and comprehensive generation of useful somaclones or mutants for resistance to various abiotic and biotic stresses and its potential applications in fruit crops. Tissue culture techniques allows pre-selection of mutagenized cells and tissues which can be regenerated into plantlets with specific resistant character. The high potentiality of selection system is based on the fact that it is possible to grow thousands of cells in petridishes or in flasks and also to achieve rapid multiplication of cell populations on the defined media (Carlson, 1973). It also helps in isolation of lines tolerant to abiotic and biotic stresses. These plants may serve as an excellent donor of the resistance gene(s) in breeding programmes. The achievement of selection technique to obtain the tolerant plant requires the availability of: (1) high variation of cell, (2) easy application of selection method, (3) regeneration method of tolerant cell and (4) the desired character to be inherited (Widodo and Sudarsono, 2005).
SELECTION AGENTS-PHYTOTOXINS
Plant pathologists have concerned themselves mainly with the chemistry and biochemistry of phytotoxins and their roles in plant pathogenesis (Huang, 2001). Pathogenic fungi and bacteria often damage their host (plants) tissues by producing toxic metabolites, which induced various symptoms such as necrosis, chlorosis, wilting, water soaking and eventually the death of plants. Phytotoxins are secondary metabolites produced by plants pathogenic microorganisms (fungi and bacteria) having low molecular weight substances and well-characterized single molecules for fungus resistance, which are easy to use in the selection experiments, which offers an alternative means of screening disease resistant cell lines. These toxic metabolites also known is one of the weapons used by pathogen inducing disease condition in susceptible host plants (Berestetskiy, 2008). However, in the past two decades, research has been extended to the use of phytotoxins as tools in screening and breeding for disease resistance (Svabova and Lebeda, 2005). The general strategy is based on considerable evidence that there is a correlation between tolerance/resistance to phytotoxins and resistance to the pathogen. Phytotoxins and culture filtrates of pathogens have been used in root growth, leaf chlorosis and lesion development assays in screening early segregating populations. They have also been used for selection of tolerant protoplasts, cells and calli, which are subsequently regenerated into putative resistant plants (Huang, 2001; Svabova and Lebeda, 2005). Phytotoxins have been recognized as a useful tool for the induction and selection of disease resistance plants using selection (Chawla and Wenzel, 1987a) and categorized as virulence or pathogenicity factor that can be used in screening and breeding for disease resistance (Vidyasekaran et al., 1990).
They are classified into two major groups, those that are non-specific and those that are host-specific toxins. Among these metabolites Fusaric Acid (FA) (5-n-butylpuridine 2-carboxyl acid) and deoxynivalenol (DON) seem to play disease development role in plants significantly. Often, the -resistant species are also FA-tolerant (Remotti and Loffler, 1996). Essential conditions for attributing the function of a disease determinant to a toxin are: (1) the demonstration that the toxin occurs in infected plants and (2) the ability of the toxin to cause at least a portion of the syndrome when placed in healthy plants (Aducci et al., 1997). Although, toxins may play important roles in some plant diseases, their possible use for selecting resistant genotypes of the host depends on two observations: (1) all reliably identified isolates of the fungus produce toxin(s) in culture and (2) the symptoms of the disease (chlorosis, necrosis, breakage of the stem, etc.) are also elicited by the toxins alone (Hamid and Strange, 2000).
Selection of the right selective agent is the critical step in establishing a successful screening system. In order to ensure that toxin-resistant plants regenerated through tissue culture will also be disease resistant; one must have some preliminary evidence for the role of the toxin in the disease (Daub, 1986). Selection is through culture filtrates as selection agent for screening of plant cells or tissues resistant to pathogens. In vitro screening has a particular purpose for pathogens that are dangerous when spread in the environment. There are several possibilities for selective agents as HSTs non host-specific toxins, living pathogens (Sacristan and Hoffman, 1979), chemicals (Carlson, 1973). Two criteria must be satisfied before a toxin is used as a screening agent: (1) the toxin produced by the pathogen must be involved in the disease development and (2) the toxin must act directly at the cellular level (Hammerschlag, 1984; Yoder, 1980). Before applying a certain toxin as selective agent, a sensitive enough bioassay system is needed to be developed which will prove the selectable toxicity of the chosen product. Then the optimum dosage for selection must be determined. Selection with host-specific toxins can result in toxin resistant plants; with complete resistance to the pathogen (qualitative) is a major result of gene mutation (Amusa, 2006). Selection for an increased level of tolerance could select for an individual with incomplete resistance, possibly determined by mutation in minor genes (quantitative resistance). Selection pressure can be applied at either the population level or on the plant regenerated from cell cultures and followed by selection in conventional field plots. While, plant and cell tissue culture techniques allow screening of very large populations of cells and regenerated plants in a small space and in a much more controlled environment than in conventional field trials.
DISEASE RESISTANCE
Disease resistance is a complex biological phenomenon involving host-pathogen and environmental interactions. Therefore, understanding the host pathogen interaction is a prime requisite for unrevealing the phenomenon of disease resistance in the host. This is a limiting factor which restricts the conventional breeding approach for disease resistance (Amusa, 2006). As an alternative, simple tissue culture based selection method is practiced in many crop plants which have potential application in fruit crops. Therefore, in vitro selection has become an indispensable tool in disease resistance breeding to increase the speed or efficiency of the breeding process, to improve the accessibility of existing germplasm and to create new variation for crop improvement. Disease conditions in plant usually result from the interaction between the host plant, the pathogen and then environment, also known as the disease triangle. These factors often limit plant disease development as well as the use of microbes directly in biological control of plant diseases. In vitro techniques used for disease resistance in fruit crops provide systems that are analogous to the prokaryotic systems, where mutations can be efficiently induced and variants selected and isolated at the cellular level. In vitro selection represents an immediate and inexpensive way of generating/selecting plant variants with tolerance to either the pathogen or its toxin from susceptible varieties, as compared to classical crossing methods. Such techniques along with others may allow introgressing only desired traits to economically important varieties, without dramatically changing other desirable agronomical and resistance characters, as reported by Evans and Sharp (1986). The direct use of selection at the cellular level is a powerful and valuable approach especially when the improvements of one or two easily identifiable characters are desired in an important variety. The main advantage is that the basic genotype of the candidate variety is usually altered slightly as opposed to procedures involving hybridization of two distinct varieties. Desired character(s) can be induced in a variety, thus reducing the time required to breed and improve a variety with the same desired character(s) through conventional hybridization methods. The most extensive studies, aimed at checking for phenotypic variation in tissue culture-propagated plants, have been conducted with among the fruit crops. Screening for somaclonal variation has a significant impact on improvement and for instance somaclonal variants had been identified for Panama or wilt disease (sp. cubense) resistance. Development of screening procedures to assess resistance and susceptibility of apple (Viseur and Tapia Figueroa, 1987) and pear (Duron et al., 1987; Brisset et al., 1988) to (fire blight) have been reported.
CULTURE FILTRATE BASED SELECTION
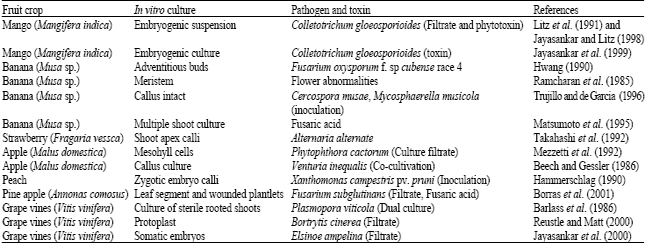
Culture filtrate of the fungal or bacterial pathogen has been used extensively for selection of resistant host species. The preliminary and basic requirement for selection of resistant lines at cellular level is to generate genetic variability in the cells. In this approach, plantlets are selected from survived explants in culture filtrate amended medium and then regenerated to get the disease resistant plantlets. In vitro selection studies on plant pathogenic bacteria are restricted to genera Clavibacter, Erwinia, Pseudomonas and Xanthomonas. While in fungi studies are reported in the genera Fusarium, Phytophthora, Alternaria, Helminthosporium in various fruit crops like mango, papaya, strawberry, banana and peach etc. For instance, in banana sp. against wilt resistance using culture filtrate as selection agent has been reported by Matsumoto et al. (1999). In grapevine, culture filtrates at cellular level were found to have resistance to and were verified (Jayasankar et al., 2000; Jayasankar and Gray, 2003). Among fruit crops, studies for selection of resistant lines have been well reported in papaya, guava, banana, mango, strawberry, peach and apple (Table 1).
TOXIN CHARACTERIZATION AND TOXIN BASED SELECTION
Toxins are compounds that are produced by the pathogens and cause part or all of the symptoms of a disease. They are of various chemical types and include peptides, glycoproteins, polysaccharides, organic acids, fatty acids and derivatives, polyketides and terpenoids (Daub, 1984; Turner, 1984). Toxic culture filtrates and purified toxins have been used for selection of disease resistant plants (Thakur et al., 2002; Liu et al., 2005; Bajpai et al., 2007). They are toxic to plants and play an important role in host-pathogen interactions and in disease. In vitro selection against disease causing phytotoxins offers an alternative means of screening disease resistant cell lines apart from using culture filtrates. During last decade there was a remarkable development in studies on the role of fungal toxins in plant pathogenesis. For instance, species has been reported to produce a variety of potent phytotoxins such as fumonisins, moniliformin, fusaric acid, 2, 5-anhydro-D-glutinol (AhG) and trichothecenes (Chawla and Wenzel, 1987 a, b; Abbas et al., 1991; Abbas and Boyette, 1992).
In toxin based in vitro selection approach, toxins from the pathogenic culture filtrate is methanol precipitated and fractionated by column chromatography using Norite-Celite column matrix. These toxin fractions are then screened for culture systems for selecting resistant lines.
| Table 1: | In vitro selection for disease resistance in fruit crops |
 | |
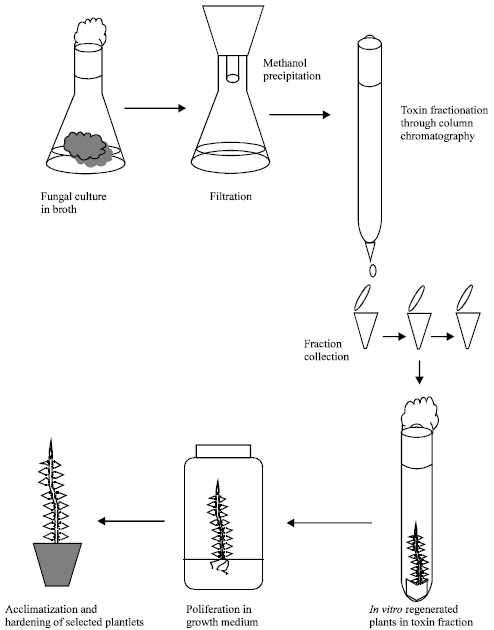 | |
| Fig. 1: | Schematic representation of steps involved during in vitro selection for fungal disease resistant lines |
The selected plants are then acclimatized and evaluated for disease resistance. The steps involved in toxin based selection for disease resistance (Fig. 1) involves toxin purification, characterization and screening for selection of toxin resistant lines. This approach is more advantageous over culture filtrate as specific toxins are used in the selection process. For example, fusaric acid isolated and purified from sp. race I was used for selecting multiple bud clumps to generate resistant lines against wilt in banana (Widodo and Sudarsono, 2005). Strategy based on the use of more or less purified culture filtrates of the pathogen as a selective agent. This type of in vitro selection has been first described by Behnke (1979), using toxins derived from, culture filtrates of the pathogen. A positive correlation has been found between toxin resistance at the cellular level and resistance to the pathogen at the plant level (Thanutong et al., 1983; Mitchell, 1984; Ling et al., 1985; Nyange et al., 1995; Rines and Luke, 1995; Jayasankar et al., 2000; Prasad and Naik, 2000; Wolpert et al., 2002; El-Hadrami et al., 2005; Sengar et al., 2009).
SOMACLONAL VARIATION AND DISEASE RESISTANCE
The genetic variability present in somatic cells, plants or plant progenies derived from cells/tissue cultured is called somaclonal variation. Larkin and Scowcroft (1983) coined the term somaclonal variation for all plant variants derived from any form of cell or tissue culture. It occurs among the population of plant resulted from culture. It is caused due to ploidy changes, nuclear fragmentation, inter and intra chromosomal translocations, somatic crossing over and sister chromatid exchanges, gene amplification and transposable genetic elements. In vitro selection is one way of somaclonal variation method. Its effectiveness and efficiency are due to its ability of changing the plant to the desired character, either by applying a selection agent on the culture media or by giving particular condition to change the somaclones with the desired character. Thus, somaclonal variants obtained by such selection process may provide a quick and useful means of obtaining agriculturally valuable traits such as disease resistance, stress (salt, low temperature) resistance, improved yield and efficient nutrient uptake (Brar and Jain, 1998). Somaclonal variation frequency is determined by a number of factors, including genotypes, explants source, duration of culture and medium composition (Brar and Jain, 1998). It can be used as an excellent tool for inducing variation in fruit improvement (Hammerschlag and Bauchan, 1984; Hwang, 1990; Brazolot et al., 1994; Karp, 1995; Jayasankar et al., 1999; Vos et al., 1998). According to the progenies of the selected somaclones may not differ significantly from the starting material. The direct use of selection at the cellular level is an alternative approach especially when the improvements of one or two easily identifiable characters are desired in an important variety. The main advantage is that the basic genotype of the candidate variety is usually altered slightly as opposed to procedures involving hybridization of two distinct varieties. Desired character(s) can be induced in a variety, thus reducing the time required to breed and improve a variety with the same desired character(s) through conventional hybridization methods (Wenzel and Foroughi-Wehr, 1990). The somaclonal variants with resistance to pathogens in fruit crops are limited only few (Table 2). For example, somaclones showing resistance to were obtained from pear (Chevreau et al., 1990). Somaclones showing resistance to were reported in mango by using partially purified phytotoxins (Jayasankar et al., 1999). In Strawberry, somaclones resistant to Toyoda et al. (1991) to Takahashi et al. (1992) and to Battistini and Rosati (1991) have also been reported. Therefore, this is an efficient approach for genetic improvement in fruit crops limited by narrow germplasm and long regeneration cycle, asexually propagated plants like bananas and for isolation of biochemical mutants like auxotroph.
| Table 2: | In vitro culture and selection of somaclonal variants in fruit crops |
 | |
Somaclonal variation approach with disease resistance character is advantageous for several reasons, including direct selection of novel phenotypes from large populations of physiologically uniform cells under defined conditions, within a limited space and within a short period of time. Ease of exposure to mutagens and the availability of haploid cells for selection of recessive traits. The cultured cells can be uniformly exposed to the selective agent, thus reducing the incidence of escapes. Culture system maintained in small spaces can potentially replace expensive greenhouse or field testing facilities. The disease causing agent remains confined to the laboratory and generation of disease resistant plants (Brazolot et al., 1994). A reliable protocol for regenerating plants from cell cultures must be available. This protocol should facilitate plant regeneration following several cycles of selection, because morphogenetic potential is often lost when cells and tissues are sub-cultured many times (Thorpe, 1982). A correlation must be established between expressions of the character being selected at the cellular level with expression at the whole plant level. Undesirable genetic changes may occur during recurrent selection. Polyploidy, aneuploidy and gross genetic changes have been observed among plants regenerated from tissue cultures (Schaeffer and Sharp, 1981) and these changes have been shown to increase with the length of time cells are in culture (Barbier and Dulieu, 1980). If possible, steps should be taken to facilitate the production of large numbers of regenerated plants which will increase the probability of recovering fertile genotypes of the desired phenotype (Tomes and Swanson, 1982). In addition, genetic stability of cell cultures should also be monitored and steps should be taken to minimize gross cytological changes. It is often difficult and time consuming to determine the ploidy of cultured cells; however, the use of cytofluorometry can simplify this task because large numbers of cells (at any stage in the cell cycle) can be measured in a short period of time (Hammerschlag, 1983). Screening for mutants at the whole plant level is a much simpler approach than selection.
In vitro cultures of tissues, organs and shoots were often used to test crop plants susceptibility to infection with various fungal pathogens. In recent years, pathotoxins (fungal toxins) have been identified as useful selection agents for induction of disease resistance through somaclonal variation in fruit crops (Table 1) in banana against causing black Sigatoka disease (Okole and Schulz, 1997), mango against causing Anthracnose (Jayasankar et al., 1999), strawberry against wilt (Orlando et al., 1997; Iwona et al., 2008), papaya against phytopthora wilt (Sharma and Skidmore, 1988), guava against wilt (Vos et al., 1998; Bajpai et al., 2007), peach against Xanthomonas (Hammerschlag, 1988) and apple against black Spot caused by Raman and Goodwin (2000) etc.
CONCLUSION AND FUTURE PROSPECTS
Traditional modes of breeding for resistance to pathogens are comprised of screening for genetic variability, searching for natural sources of resistance, pathogenicity testing and complemented with inheritance studies. Cell culture systems are well defined controlled environments, away from limitations of availability of space, time and variations due to environmental effects which are major bottlenecks in conventional breeding in perennials. New trends in resistance breeding use a combination of tissue and cell cultures with selections and classical breeding methods. Somaclonal variability alone offers new possible sources of altered characters, including susceptibility and/or resistance to pathogens. The somaclones should be monitored for the new features (Lebeda, 1995). Other methods utilize anther cultures, somatic hybridization and also genetic transformation as a way of molecular breeding of plants. An important aspect of crop improvement for disease resistance via a selection strategy is that traits selected at the cellular level must be expressed in the regenerated plants. Somaclonal variation represent useful source for introduction of valuable variations in fruit trees. These variations occur at much higher frequencies than induced mutants which are associated with undesirable features. Therefore, somaclonal variation can be combined with mutagenesis. In vitro selection of genetically stable somaclones will save time to develop traits such as disease resistance, which is a cost-effective approach over conventional breeding. Perhaps, selected putative variants should be finally field-tested to confirm the genetic stability of the disease resistant trait (Jain, 2001). It seems selection technology would be a candidate approach for disease resistance breeding in fruit crops.
The selection of somaclonal variants and induced mutants is not protected under intellectual property regulations, nor is it subject to public safety concerns that currently hamper transgenic (i.e., GMO) approaches for the development of new crop cultivars. Thus, selection is a promising, non-transgenic approach, which offers an attractive alternative method for producing improved cultivars (Jayasankar and Gray, 2003).
ACKNOWLEDGMENT
Authors are highly thankful to Director CISH for providing necessary financial research grant and preparation of manuscript is gratefully acknowledged.
REFERENCES
- Abbas, H.K. and C.D. Boyette, 1992. Phytotoxicity of fumonisin B1 on weeds and crop species. Weed Technol., 6: 548-552.
Direct Link - Abbas, H.K., C.D. Boyette, R.E. Hoagland and R.F. Vesonder, 1991. Bioherbicidal potential of Fusarium moniliforme and its phytotoxins fumonisin. Weed Sci., 39: 673-677.
Direct Link - Amusa, N.A., 2006. Microbially produced phytotoxins and plant disease management. Afr. J. Biotechnol., 5: 405-414.
Direct Link - Amusa, N.A., 1994. Production, purification and bioassay of toxic metabolites of three plant pathogenic species of Colletotrichum in Nigeria. Mycopathology, 128: 161-166.
CrossRef - Bajpai, A., R. Chandra, M. Mishra and R.K. Tiwari, 2007. Regenerating psidium spp. For screening wilt resistance rootstock under in vitro conditions. Acta Hortic., 535: 145-154.
Direct Link - Barlass, M., R.M. Miller and A.J. Antcliff, 1986. Development of methods for screening grapevines for resistance to infection by downy mildew I Dual culture in vitro. Am. J. Enol. Vitic., 37: 61-66.
Direct Link - Beech, I. and C. Gessler, 1986. Interaction between Venturia inequalis and apple callus tissue cultures: An electron microscopic study. J. Phytopathol., 115: 315-322.
CrossRef - Behnke, M., 1979. Selection of potato callus for resistance to culture filtrates of Phytophthora infestans and regeneration of resistant plants. Theor. Applied Genet., 55: 69-71.
CrossRef - Berestetskiy, A.O., 2008. A review of fungal phytotoxins: From basic studies to practical use. Applied Biochem. Microl., 44: 453-465.
CrossRef - Brar, D.S. and S.M. Jain, 1998. Somaclonal Variation: Mechanism and Applications in Crop Improvement. In: Somaclonal Variation and Induced Mutations in Crop Improvement, Jain, S.M., D.S. Brar and B.S. Ahloowalia (Eds.). Springer, Dordrecht, London, ISBN: 0792348621, Pages: 15-37.
CrossRefDirect Link - Chandra, R. and M. Mishra, 2007. Biotechnological interventions for improvement of Guava (Psidium guajava L.). Acta Hortic., 735: 117-125.
Direct Link - Chawla, H.S. and G. Wenzel, 1987. In-vitro selection for fusaric acid resistant barley plants. Plant Breed., 99: 159-163.
CrossRefDirect Link - Chawla, H.S. and G. Wenzel, 1987. In-vitro selection of barley and wheat for resistance against Helminthosporium sativum. TAG, 74: 841-845.
CrossRef - Daub, M.E., 1986. Tissue culture and the selection of resistance to pathogens. Annu. Rev. Phytopathol., 24: 159-186.
Direct Link - Dolcet-Sanjuan, R., W.S. Mok and M.C. Mok, 1990. Micropropagation of Pyrus and Cydonia and their responses to Fe-limiting conditions. Plant Cell Tissue Organ Culture, 21: 191-199.
CrossRef - El Hadrami, A., A. El Idrissi-Tourane, M. El Hassani, F. Daayf and I. El Hadrami, 2005. Toxin-based in-vitro selection and its potential application to date palm for resistance to the bayoud fusarium wilt. A review. Comptes Rendus Biol., 328: 732-744.
CrossRefDirect Link - Hamid, K. and R.N. Strange, 2000. Phytotoxicity of solanapyrones A and B produced by the chickpea pathogen Ascochyta rabiei (Pass.) Labr. and the appparent metabolism of solanapyrone A by chickpea tissues. Physiol. Mol. Plant Pathol., 56: 235-244.
CrossRef - Hammerschlag, F.A., 1988. Selection of peach cells for insensitivity to culture filtrate of Xanthomonas campestris pv. Pruni and regeneration of resistant plants. Theor. Applied Genet., 76: 865-869.
CrossRef - Hammerschlag, F.A., 1990. Resistant responses of plants regenerated from peach callus to Xanthomonas campestris pv. pruni. J. Am. Soc., 115: 1034-1037.
Direct Link - Iwona, S., M. Lech and W. Danuta, 2008. A method for in vitro testing strawberry susceptibility to Verticillum wilts. J. Fruit Ornamental Plant Res., 16: 111-121.
Direct Link - Jayasankar, S. and D.J. Gray, 2003. In vitro selection for disease resistance in plants- an alternative to genetic engineering. AgBiotechNet, 5: 1-5.
Direct Link - Jayasankar, S., R.E. Litz and D.J. Gray, 2000. In vitro selection of Vitis vinifera `Chardonnay` with Elsinoe ampelina culture filtrates is accompanied by fungal resistance and enhanced secretion of chitinase. Planta, 211: 200-208.
PubMed - Jayasankar, S., R.E. Litz, D.J. Gray and P.A. Moon, 1999. Responses of embryogenic mango cultures and seedling bioassays to a partially purified phytotoxin produced by a mango leaf isolate of Colletotrichum gloeosporioides Penz. In Vitro Cell. Dev. Biol. Plants, 35: 475-479.
CrossRefDirect Link - Karp, A., 1995. Somaclonal variation as a tool for crop improvement. Euphytica, 85: 295-302.
CrossRefDirect Link - Lane, W.D. and N.E. Looney, 1982. A selective tissue culture medium for growth of compact (dwarf) mutants of apple. Theor. Applied Genet., 61: 219-223.
CrossRefDirect Link - Larkin, P.J. and W.R. Scowcroft, 1983. Somaclonal variation and eyespot toxin tolerance in sugar cane. Plant Cell Tissue Organ Culture, 2: 111-122.
CrossRef - Ling, D.H., P. Vidyasekharan, E.S. Borromeo, F.P. Zapata and T.W. Mew, 1985. In vitro screening of rice germplasm for resistance to brown spot disease using phytotoxins. Theor. Applied Genet., 71: 133-135.
CrossRef - Liu, S., H. Wang, J. Zhang, B.D. Fitt and Z. Xu et al., 2005. In vitro mutation and selection of doubled haploid Brassica napus lines with improved resistance to sclerotinia sclerotiorum. Plant Cell Rep., 24: 133-134.
PubMed - Lynch, J.M. and S.J. Clark, 1984. Effects of microbial colonization of barley (Hordeum vulgare L.) roots on seedling growth. J. Applied Bacteriol., 56: 47-52.
CrossRef - Matsumoto, K., M.L. Barbosa and L.A.C. Souza, 1995. Race 1 Fusarium wilt tolerance on banana plants selected by fusaric acid. Euphytica, 84: 67-71.
CrossRefDirect Link - Mezzetti, B., R.H. Zimmerman, C. Mischke, P. Rosati and F.A. Hammerschlag, 1992. Merocyanine 540 as an optical probe to monitor the effects of culture filtrates of Phytophthora cactorum on apple cell membrane. Plant Sci., 83: 163-167.
CrossRef - Mitchell, R.E., 1984. The relevance of non-host-specific toxins in the expression of virulence by pathogens. Ann. Rev. Phytopathol., 22: 215-245.
CrossRef - Nyange, N.E., B. Williamson, R.J. McNicol, G.D. Lycon and C.A. Hackett, 1995. Selection of callus for resistance to partially purified phytotoxic culture filtrates from Colletotrichum kahawae. Ann. Applied Biol., 127: 425-439.
CrossRef - Orlando, R., P. Magro and E. Rugini, 1997. Pectic enzymes as a selective pressure tool for in vitro recovery of strawberry plants with fungal disease resistance. Plant Cell Rep., 16: 272-276.
CrossRef - Prasad, V. and G.R. Naik, 2000. In vitro strategies for selection of eye-spot resistant sugarcane lines using toxins of Helminthosporium sacchari. Indian J. Exp. Biol., 38: 69-73.
PubMed - Raman, H. and P.B. Goodwin, 2000. In vitro Screening of Apple Germplasm for Resistance against Black Spot caused by Venturia inaequalis. J. New Seeds, 2: 37-46.
CrossRef - Remotti, P.C. and H.J.M. Loffler, 1996. The involvement of fusaric acid in the bulb-rot of Gladiolus. J. Phytopathol., 144: 405-411.
CrossRef - Remotti, P.C., H.J.M. Loffler and L. van Vloten-Doting, 1997. Selection of cell-lines and regeneration of plants resistant to fusaric acid from Gladiolus × Grandifolorus cv. Peter Pears. Euphytica, 96: 237-245.
CrossRefDirect Link - Sacristam, M.D. and F. Hoffman, 1979. Direct infection of embyogenic tissue cultures of haploid Brassica napus with resting spores of Plasmodiophora brassica. Theor. Applied Genet., 54: 129-132.
CrossRef - Schaeffer, G.W. and F.T. Sharp, 1981. Lysine in seed protein from S-amino ethyl-L-cysteine resistant anther derived tissue cultures of rice. In vitro Cell. Dev. Biol. Plants, 17: 345-352.
Direct Link - Sengar, A.S., S. Thind, B. Kumar, P. Mittal and S.S. Gosal, 2009. In vitro selection at cellular level for red rot resistance in sugarcane (Saccharum sp.). Plant Growth Regul., 58: 201-209.
CrossRef - Sharma, N.K. and D.I. Skidmore, 1988. In vitro expression of partial resistance to Phytophthora palmivora by shoot cultures of papaya. Plant Cell Tissue Organ Culture, 14: 187-196.
CrossRef - Skirvin, R.M., 1978. Natural and induced variation in tissue culture. Euphytica, 27: 241-266.
CrossRefDirect Link - Svabova, L. and A. Lebeda, 2005. In vitro selection for improved plant resistance to toxin-producing pathogens. J. Phytopathol., 153: 52-64.
CrossRef - Takahashi, H., T. Takatsugu and M. Tsutomu, 1992. Resistant plants to Alternaria alternata strawberry pathotype selected from calli clones of strawberry cultivar Morioka-16 and their characteristics. J. Jap. Soc. Hort. Sci., 61: 323-329.
Direct Link - Thakur, M., D. Sharma and S. Sharma, 2002. In vitro selection and regeneration of carnation (Dianthus caryophyllus L.) plants resistant to culture filtrate of Fusarium oxysporum f. sp. dianthi. Plant Cell Rep., 20: 825-828.
CrossRefDirect Link - Thanutong, P., I. Furussawa and M. Yamamoto, 1983. Resistant tobacco plants from protoplast-derived calluses selected for their resistance to Pseudomonas and Alternaria toxins. Theor. Applied Genet., 66: 209-215.
CrossRefDirect Link - Thorpe, T.A., 1982. Callus Organization and de Novo Formation of Shoots Roots and Embryos in vitro. In: Application of Plant Cell and Tissue Culture to Agriculture and Industry, Tomes, D.T., B.E. Ellis, P.M. Harney, K.J. Kasha and R.L. Peterson (Eds.). University of Guelph, Guelph, Ontario, Canada, pp: 115-138.
- Tomes, D.T. and E.V. Swanson, 1982. Application of in vitro Selection to Plant Improvement. In: Application of Plant Cell and Tissue Culture to Agriculture and Industry, Tomes, D.T., B.E. Ellis, P.M. Harney, K.J. Kasha and R.L. Peterson (Eds.). University of Guelph, Guelph, Ontario, Canada, pp: 25-43.
- Toyoda, H., K. Horikoshi, Y. Yamano and S. Ouchi, 1991. Selection of Fusarium wilt disease resistance from regenerants derived from leaf callus of strawberry. Plant Cell Rep., 10: 167-170.
CrossRef - Utkhede, R.S., 1986. In vitro screening of the world apple germplasm collection for resistance to Phytophthora cactorum crown rot. Scientia Hortic., 29: 205-210.
CrossRef - Vidyasekaran, E.S., J. Birromeo and T.W. Mew, 1986. Host specific toxin production by Helminthosprium oryzae. Phytopathology, 76: 261-262.
CrossRef - Vidyasekaran, P., D.H. Ling, E.S. Borromeo, F. J. Zapata and T.W. Mew, 1990. Selection of brown resistant rice plants from Helminthosporium oryzae toxin resistant calluses. Ann. Applied Biol., 117: 515-523.
CrossRef - Vos, J.E., M.H. Schoeman, P. Berjak, M.P. Watt and A.J. Toerien, 1998. In vitro selection and commercial release of guava wilt resistant rootstocks. Acta Hortic., 513: 69-79.
Direct Link - Walker, H.L. and G.E. Templeton, 1978. In vitro production of a phytotoxic metabolite by Colletotrichum gloeosporiodes f. sp. aeschynomene. Plant Sci. Lett., 13: 91-96.
CrossRefDirect Link - Wenzel, G. and B. Foroughi-Wehr, 1990. Progeny tests of barley, wheat and potato regenerated from cell cultures after in vitro selection for disease resistance. Theor. Applied Genet., 80: 359-365.
CrossRefDirect Link - Wheeler, H. and H. Luke, 1955. Mass screening for disease resistant mutants in Oats. Science, 122: 1229-1229.
CrossRef - Wolpert, T.J., L.D. Dunkle and L.M. Ciuffetti, 2002. Host-selective toxins and avirulence determinants: What's in a name? Annu. Rev. Phytopathol., 40: 251-285.
CrossRefDirect Link - Wolpert, T.J., D.A. Navarre, D.L. Moore and V. Macko, 1994. Identification of the 100-KD victorin binding protein from oats. Plant Cell, 6: 1145-1155.
Direct Link - Larkin, P.J. and W.R. Scowcroft, 1981. Somaclonal variation-a novel source of variability from cell cultures for plant improvement. Theor. Applied Genet., 60: 197-214.
CrossRefDirect Link
pradeep kumar Reply
Very good paper with all information regarding "in vitro selection" and also related to somaclonal variation study in relation to resistant development by using toxin..