
Yasser M. Awad
Department of Agricultural Botany, Faculty of Agriculture, Suez Canal University, Ismailia, 41522, Egypt
A-G.I.O. Baz
Department of Agricultural Botany, Faculty of Agriculture, Suez Canal University, Ismailia, 41522, Egypt
Mohamed M. Sakr
Department of Agricultural Botany, Faculty of Agriculture, Suez Canal University, Ismailia, 41522, Egypt
Yong Sik OK
Department of Biological Environment, College of Agriculture and Life Sciences, Kangwon National University, Chuncheon 200-701, Korea
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 8 | Page No.: 536-543







ABSTRACT
The objective of this research was to evaluate the effects of morphactin, on the growth and anatomical features of soybean plants (Glycine max L. Merrill) cv. Crawford. Field experiments were conducted under different irrigation regimes (every 5 and 10 days) on soybeans (Glycine max L. Merril) that were planted in a newly reclaimed sandy soil at the Experimental Farm at Suez Canal University, Egypt during seasons of 1998 and 1999. Different concentrations of the morphactin (0, 50 and 200 mg L-1) were added at 40 days after sowing by foliar application. The morphactin treatments significantly decreased the plant height (14.88 and 21.44% inhibition of stem elongation under the two irrigation regimes, respectively), while they increased the number of branches. The pod number plant-1, seed number plant-1, dry weight (g), nodule number plant-1, reducing sugar content (mg g-1 DW) and total phenols (mg g-1 DW) increased significantly in response to morphactin treatment at 200 mg L-1. Similarly, the thickness of the midrib, mesophyll, vascular bundle, xylem, cambium + phloem and xylem vessel in the main vascular bundle increased in response to treatment with morphactin at 200 mg L-1. Low irrigation regime (every 10 days) had adverse effect on the growth and anatomical features of soybean. Overall, it is recommended that 200 mg L-1 morphactin can be used for the stable production of soybean (Glycine max L. Merrill) in newly reclaimed sandy soils in Egypt.
PDF Abstract XML References Citation
How to cite this article
Yasser M. Awad, A-G.I.O. Baz, Mohamed M. Sakr and Yong Sik OK, 2009. The Effect of Morphactin (Methyl 2-Chloro-9-hydroxyfluorene-9-carboxylate) on the Growth and Anatomical Features in Soybean (Glycine max (L). Merril) Cultivar. Asian Journal of Plant Sciences, 8: 536-543.
DOI: 10.3923/ajps.2009.536.543
URL: https://scialert.net/abstract/?doi=ajps.2009.536.543
DOI: 10.3923/ajps.2009.536.543
URL: https://scialert.net/abstract/?doi=ajps.2009.536.543
INTRODUCTION
Soybean (Glycine max L. Merrill), which is a member of the family Fabaceae, is an ancient leguminous crop that is to be extensively cultivated in Egypt. The wild genus of Glycine was first found in Japan and Korea (Hymowitz and Singh, 1987; Kanazawa et al., 1998). The cultivated soybean is likely to be derived from the subgenus Soja (Moench) (Hartwig, 1973). The development of soybean cropping systems has recently received a great attention due to the shortage in protein and vegetable oil in Egypt (Baz et al., 1984; El-Kheir et al., 1994; El-Batal and Abddel-Gawad, 1995). In general, soybean seeds contain 35-40% protein on a dry weight basis (Mujoo et al., 2003); therefore, soybean production will be play a major role in meeting the demands for protein in Egypt. In Egypt, approximately 16500 ha of soybeans were cultivated while the average yield was 3.303 tons ha-1 in 2007 (FAO, 2007).
Morphactins are a group of synthetic bioregulatants with a high morphogenetic potency that affect almost every stage of plant growth and development (El-Desoki et al., 1994). Morphactin has recently a great attention due to its ability to enhance plant cell production of secondary metabolites with many health benefits including antioxidant (Mathur and Ramawat, 2008: Roat and Ramawat, 2009). For example, Cantliffe et al. (1972) and Lukasik (1975) reported that morphactin treatment increased the number of pods per soybean plant, but had little effect on the development of fruit. However, Smolinski (1976) reported that foliar applications of morphactin to Vicia faba L. cv. Hangdown plants led to significantly increased numbers of lateral branches per plant due to the inhibitory effect of morphactin, which weakened the apical dominance of the plant. In addition, foliar application of 10 μM methylchloflurenol (morphactin) to Glycine max var. Anoka one week after flowering produced a 40% increase in pod number and seed number with only a 12% inhibition in stem elongation (Nooden and Nooden, 1985). Peterson et al. (1990) also pointed out that treatment of soybean plants with morphactins resulted in increases in pod set or delays in flowering and pod abscission. In addition, application of a foliar spray containing 100 mg L-1 chlorflurenol to one-month-old Phaseolus vulgaris cv. Carioca plants decreased their height at 14 and 21 days after treatment in a greenhouse trial (Castro et al., 1990). Furthermore, the application of morphactin at 10~200 mg L-1 increased the number of pods plant-1 and decreased the height of Vicia faba L. var. Giza 3 when compared to the control plants (El-Desoki et al., 1994).
Conversely, morphactin foliar applications to Vicia faba L. cv. Hngdown reduced the size of the epidermal and mesophyll cells in the secondary stems, but increased the number of vascular bundles when compared to the control (Smolinski, 1976). Furthermore, morphactin CF125 (2-chloro-9-hydroxyfluoren-9-carboxylic acid) foliar applications at 25-200 mg L-1 to Vicia faba cv. Giza 3 induced increased leaf thickness, except at 50 mg L-1. Moreover, application of morphactin at 25 and 200 mg L-1 increased the thickness of palisade tissues, while treatment with intermediate concentrations showed the opposite effect. Additionally, treatment with 100 mg L-1 CF125 decreased the midrib thickness, but this was increased in response to treatment with other concentrations (Ali et al., 1994). Additionally, treatment with morphactin at all concentrations was found to increase stem diameter, while treatment with 25 and 50 mg L-1 CF125 induced xylem differentiation, whereas xylem differentiation was inhibited by the higher concentrations that simulated the formation of thicker leaflets (Ali et al., 1994).
D’Andria et al. (1991) evaluated the response of two soybean cultivars, Glycine max L. Merr. cv. Hodgson and Nova, to various watering schedules and found that water stress reduced flower production, increased the percentage dropped flowers and decreased pod formation. Water availability during the vegetative period also affected plant development, especially the number of fertile nodes and the number of pods per node (D’Andria et al., 1991). Moreover, El-Kheir et al. (1994) observed that water stress decreased the plant height, number of leaves and branches/plant, total leaf area, shoot dry weight/plant, seed yield, chlorophyll a and b and carotenoid contents in leaves and the oil content, whereas it increased the seed protein and leaf proline content.
This study was conducted to evaluate the impacts of foliar application of morphactin as a plant growth retardant on the productivity, growth and anatomical characteristics of soybean plants cultivated under high density in a newly reclaimed sandy soil. In addition, the effects of the treatments on root nodule formation were evaluated.
MATERIALS AND METHODS
Field experiments were conducted using soybean plants (Glycine max L. Merrill) cv. Crawford planted in newly reclaimed sandy soil at the experimental farm of Suez Canal University during 1998 and 1999. The seeds were obtained from the Legume Research Division of the Field Crops Institute at the Agricultural Research Centre, Cairo, Egypt. Two different levels of morphactin (50 and 200 mg L-1) were applied by foliar application at 40 Days After Sowing (DAS) and the plants were cultivated under two different irrigation regimes (5 and 10 days). Morphactin was obtained from the BDH chemical company (Ltd. Poole England, UK). The control plants were sprayed with tap water.
The experimental split plot design consisted of three rows 4 m in length and 60 cm in width (plot area 7.2 m2) for each treatment. Calcium superphosphate (15% P2O5) at 238 kg ha-1 and potassium sulphate (48% K2O) at 119 kg ha-1 were added during seed-bed preparation. In addition, ammonium nitrate (46.5% N) at 71.4 kg ha-1 was dressed beside the plants prior to the second irrigation (at 16 DAS). Soybean cv. Crawford seeds were inoculated with the strain of Bradyrhizobium japonica provided by the Legume Research Division of the Field Crops Institute at the Agricultural Research Center. The seeds were sown in hills spaced 20 cm apart on April 20, 13th during 1998 and 1999 seasons. At 13 DAS, the stand was thinned to two plants per hill. All other practices were done as recommended to region.
Ten plants were sampled from each treatment at random from the middle of the plot during the harvest stage (110 DAS). Therefore, the morphological characteristics of soybean plants, including vegetative characteristics (average plant height (cm), average number of branches plant-1 and average plant dry weight without pods (g)) and reproductive characteristics (average number of pods plant-1 and average number of seeds plant-1) were recorded during both cultivation seasons using the ten plant samples. In addition, the average number of nodules plant-1 was recorded at 60 DAS. Chemical analysis was performed on leaf samples (taken at 60 DAS) collected from treated and untreated soybean plants. The samples were extracted using the method described by Abdel-Rahman et al. (1975), after which the total phenols were determined using the modified Folin-Ciocalteu method described by William et al. (1965). Moreover, reducing sugars were determined using Nelson’s method as described by Moore (1974). Transverse sections through the seventh internode of the main stem of the soybean plants and the blade of the terminal leaflet of the seventh leaf that developed on the main stem were evaluated histologically at 60 DAS. Permanent specimens were prepared and double stained with Safranin and Light green, according to the methods described by Willey (1971), after which the sections were examined microscopically and measured using an eyepiece micrometer.
Statistical analysis: The data were evaluated by one-way Analysis of Variance (ANOVA) using CoStat Version 6.311 (CoHort Software, Berkeley, CA 94701). The means were compared using the Least Significant Difference (LSD) (Steel and Torrie, 1980). Time trend analysis of both total production and the total quantity of imports was conducted.
RESULTS
The curve estimation model in Fig. 1 clearly shows that the total production decreased significantly by 715.8 ton every year, which represents 1.5% of the average total production from 1990 to 2003. Conversely, the total quantity of imports increased significantly by 25, 373 ton year-1, which represents 17.4% of the average total production during the same period.
The effect of various concentrations of morphactin on growth parameters, chemical components and the microscopic structure of the soybean (Glycine max L.) cv. Crawford were investigated in field experiments conducted during two growing seasons. As shown in Table 1, all concentrations of morphactin led to a significant decrease in the average length of the main stem when compared with the untreated plants. The lowest values of average length of the main stem were 34.6 and 33.5 cm in the plants treated with 50 mg L-1 morphactin under the first irrigation regime and 22.75 and 25.7 cm in plants treated with 200 mg L-1 under the second irrigation regime at 1st and 2nd seasons, respectively. Moreover, treatment with morphactin induced a significant increase in the average number of lateral branches plant-1. The highest numbers of branches plant-1 were 4.6 and 3.3 in the plants treated with 200 mg L-1 morphactin under the first irrigation regime, whilst the highest values were 3.3 and 2.2 branches plant-1 in plant treated with 50 mg L-1 under the second irrigation regime at 1st and 2nd seasons, respectively. The statistical analysis demonstrated that the differences in the average dry weight of the plants (without pods) increased significantly by 20.6 and 12.07 g when compared to the controls in response to treatment with 200 and 50 ppm morphactin under the first irrigation regime at the first and second seasons, respectively, but the opposite trend was observed under the second irrigation regime possibly due to drought stress.
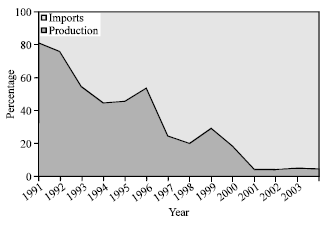 | |
| Fig. 1: | Curve estimation model for time trend analysis of the total production of soybean and total quantity of imports |
| Table 1: | The effects of morphactin treatments on the growth characteristics of soybean (Glycine max L.) cv. Crawford during the two examination seasons |
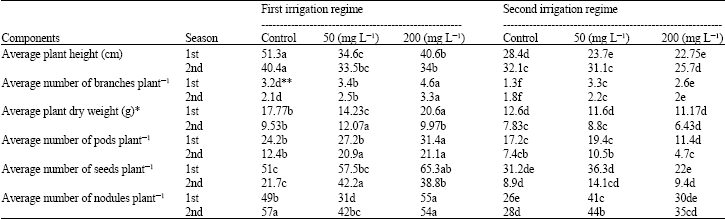 | |
| *Shoot and root dry weight without pods. **Means followed by the same letter within a row are not significantly different at the 0.05 probability level | |
As shown in Table 1, the average numbers of pods and seeds plant-1 demonstrated that the highest values were 31.4, 21.1, 19.4 and 10.5 pods plant-1 in plants treated by 200 and 50 mg L-1 under the first and the second irrigation regimes at 1st and 2nd seasons, respectively. Similarly, 65.3 and 36.3 seeds plant-1 were observed in plants treated with 200 and 50 mg L-1 morphactin under the first and the second irrigation regimes at 1st season, respectively. In the second season, there were 42.2 and 14.1 pods plant-1 observed in response to treatment with 50 mg L-1 morphactin under the first and second irrigation regimes. Furthermore, morphactin induced a significant increase in root nodules under some situations. The highest numbers of root nodules plant-1 was 55 in plants treated with 200 mg L-1 morphactin under the first irrigation regime in 1st season, while treatment with 50 mg L-1 morphactin produced the highest values under the second irrigation regime in both seasons (41 and 44 root nodules plant-1 during the first and second season, respectively).
As shown in Table 2, the levels of reducing sugars and total phenols increased in response to treatment with morphactin under both irrigation regimes when compared with the controls. The most effective treatments were 200 mg L-1 morphactin under the first irrigation regime, which resulted in 8.3 and 11.9 mg g-1 DW reducing sugars and total phenol, respectively, followed by 7.3 and 7.8 mg g-1 DW reducing sugars and total phenol in plant leaves treated with 50 mg L-1 morphactin under the second irrigation regime at 1st and 2nd seasons, respectively.
Microscopic measurements of certain characteristics in the transverse sections of the seventh internode of the main vascular bundle of soybean cv. Crawford are presented in Table 3 and Fig. 2a and b. All treatments clearly increased the average thickness of the main vascular cylinder and the average number of xylem vessels arm-1 under both irrigation regimes. The highest average thicknesses of the main vascular cylinder observed were 3.03 and 4.1 mm in response to treatment with 50 and 200 mg L-1 under the first and second irrigation regimes, respectively.
| Table 2: | Morphactin effect on chemical components of the studied soybean plants leaves (mg g-1 DW) |
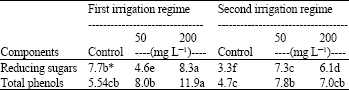 | |
| *Means followed by the same letter within a row are not significantly different at the 0.05 probability level | |
| Table 3: | The effects of morphactin treatments on the anatomical characteristics of soybean cv. Crawford stems taken at 60 DAS during the 1st season (mm) |
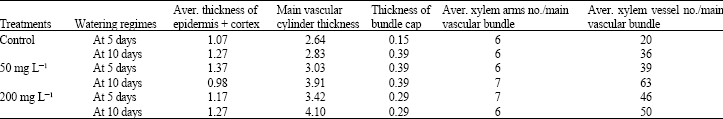 | |
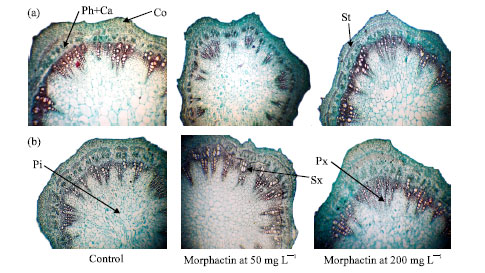 | |
| Fig. 2: | Transverse sections through the seventh internode of the soybean main stem as affected by morphactin treatments (10.24 X). Ca: Cambium; Co: Cortex; Ep: epidermis; Ph: Phloem; Pi: Pith; Px: Primary xylem; St: Sclerenchyma tissue; Sx: Secondary xylem. (a) First irrigation refime and (b) second irrigation regime |
In addition, the highest average number of xylem vessels/main vascular bundle were 46 and 63 in response to treatment with 200 and 50 mg L-1 morphactin under the first and the second irrigation regimes, respectively.
Table 4 and Fig. 3a, b and 4a, b show the data describing the internal morphological characteristics of the seventh terminal leaflet of soybean cv. Crawford. Most morphactin treatments increased the measured characteristics. Under the first irrigation regime, the highest values were recorded in response to treatment with 50 mg L-1 and then 200 mg L-1. The highest average thicknesses of the mesophyll, palisade tissue, spongy tissue, midrib, vascular bundle, xylem, cambium+phloem and xylem vessel main-1 vascular bundle were recorded under the first irrigation regime. The greatest increases in the average mesophyll thickness (0.94 mm), average thickness of the midrib (6.13 mm), average vascular bundle thickness (1.69 mm), average number of xylem arms/main vascular bundle (18) andaverage xylem vessel number/main vascular bundle (70) were observed in response to treatment with 50 mg L-1 morphactin. Conversely, treatment with morphactin at 200 mg L-1 resulted in the greatest thickness of the cambium + phloem (0.43 mm) and spongy tissue (0.51 mm). Drought stress had adverse effects on the target endomorphological parameters under the second irrigation regime (watering at 10 days), except for the average number of xylem arms/main vascular bundle, xylem vessel no./main vascular bundle and the average thickness of the midrib and xylem, which increased in response to treatment of plants under drought stress with morphactin.
| Table 4: | Soybean cv. Crawford endomorphological characteristics of the sixth terminal leaflet in response to morphactin treatments at 60 days after planting during the 1st season |
 | |
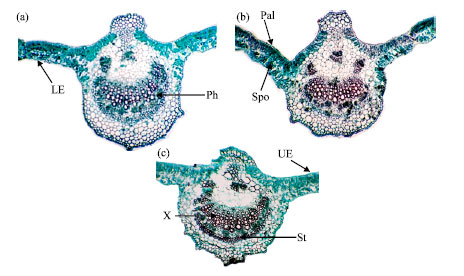 | |
| Fig. 3: | Transverse sections through the terminal leaflet blade of the seventh developed leaf on the soybean main stem in response to foliar application of morphactin under the first irrigation regime (16 X). LE: Lower epidermis; Mvb: Midvein vascular bundle, Pal: Palisade tissue, Ph: Phloem, X: Xylem, Spo: Spongy tissue, St: Sclerenchyma tissue, UE: Upper epidermis. (a) Morphactin at 50 mg L-1, (b) morphactin at 200 mg L-1 and (c) control |
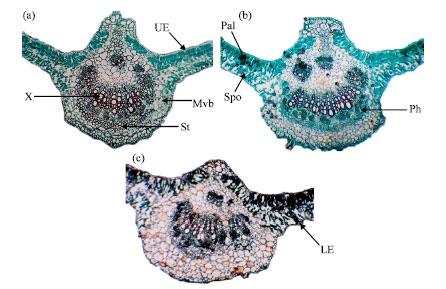 | |
| Fig. 4: | Transverse sections through the terminal leaflet blade of the developed seventh leaf on the soybean main stem after foliar morphactin application during the second irrigation regime (16 X). LE: Lower epidermis, Mvb: Midvein vascular bundle, Pal: Palisade tissue, Ph: Phloem, X: Xylem, Spo: Spongy tissue, St: Sclerenchyma tissue, UE: Upper epidermis. (a) Morphactin at 50 mg L-1, (b) morphactin at 200 mg L-1 and (c) control |
DISCUSSION
It is well known that endogenous and exogenous plant growth regulators such as auxins, gibberellins, cytokinins and growth inhibitors play an important role in modifying and regulating many physiological processes in the plants (Baz et al., 1984). Moreover, these processes are greatly influenced by environmental conditions (Leopold, 1964). Morphactin inhibited enzymes of the major metabolic pathways that affecting plant growth via hormonal regulation and morphological changes can be occurred based on the application rate (Sztajer et al., 1990). The changes in the average length of the main stem in the present study are consistent with the results of studies conducted by Nooden and Nooden (1985), Castro et al. (1990), Ali et al. (1994) and El-Desoki et al. (1994), who stated that the length of the main stem of different leguminous plants including Glycine max L., Phaseolus vulgaris and Vicia faba L. was decreased (12 and 49.23% inhibition of stem elongation of soybean and common bean plants, respectively) in response to morphactin treatments. Ziegler (1970) demonstrated that treatment with morphactin blocked gibberellin biosynthesis and reduced the content of endogenous GA-like substances, which led to decreased soybean stem elongation. Furthermore, the average number of lateral branches per plant observed in the present study was in agreement with the results of a study conducted by Wakhloo (1975), who suggested that morphactin weakens the apical dominance of the plants. The average plant dry weights observed in the present study were in accordance with the results of a study conducted by Zayed et al. (1985). Similarly, the reproductive characteristics observed in the present study were similar to those observed by Nooden and Nooden (1985) and El-Desoki et al. (1994), who treated Glycine max L. var. Anoka and Vicia faba L. var. Giza 3, respectively, with morphactin. Moreover, Baz et al. (1984) found that foliar application of growth retardants led to increased root nodule formation, numbers of root nodules and dry weight in soybean plants via a reduction in the endogenous root content of GA-like substances, but that the nodule size was similar to that of control plants, which is similar to the results of the present study. Moreover, the total sugar and phenol contents observed in the present study were similar to the levels observed by Zayed et al. (1985) and Castro et al. (1990).
In the study, the morphological characteristics were similar to the results of a study conducted by Ali et al. (1994), who found that foliar application of 25-200 mg L-1 morphactin increased stem diameter. Also, they found that xylem differentiation was induced by 25 and 50 mg L-1 CF125, which led to increase leaf thickness. Furthermore, they found that the palisade tissue thickness increased in response to treatment with 25 and 200 mg L-1, but decreased in response to intermediate concentrations. They also found that the midrib thickness was decreased by 100 mg L-1 CF125 and increased by other concentrations. Finally, they found that high concentrations of CF125 stimulated the formation of a waxy epidermal layer, swelling of cortical cells, abnormal differentiation of vascular bundles in lateral branches and the formation of thicker leaflets. Similarly, Dybing and Yarrow (1984) stated that morphactin increased the thickness of leaves and palisades, as well as the spongy mesophyll cell count.
El-Batal and Abdel Gawad (1995) stated that variations in plant response to the application of plant growth retardants among seasons may be due, at least in part, to differences in the average daily temperature, which affect the morphological and physiological response of soybean plants. This phenomenon would explain the differences that were observed between the two seasons evaluated in the present study. Similarly, Parker (2000) reported that temperature affects plant productivity and growth. Moreover, some findings differed from previous studies can be explained via the competition between plants grown at high densities (planting on two sides of a row), while in some cases referred to the effect of genotype (i.e., Vicia faba L.). There were no pronounced changes in yield and the components of the plants observed in the present study, which may have been due to the poor quality of the newly reclaimed sandy soil used in the present study.
Overall, the results of the present study revealed that treatment with morphactin had remarkable effects on the production, chemical constituents and reproductive characteristics of soybean plants, in addition to the number of root nodules per plant, even though the plants were grown in newly reclaimed sandy soils with low amounts of NPK fertilizers. However, morphactin treatment at 200 mg L-1 is recommended to induce pronounced increases in the yield of soybean (Glycine max (L). Merrill) cv. Crawford.
ACKNOWLEDGMENT
This research was supported by a grant (Code No. 2-2-3) from Sustainable Water Resources Center of 21st Century Frontier Research Program.
REFERENCES
- Abdel-Rahman, M., T. Thomas, G.J. Doss and L. Howel, 1975. Changes in endogenous plant hormones in cherry, tomato fruits during development and maturation. Physiol. Plant., 34: 39-43.
CrossRef - Castro, P.R., B. Appezzato, C.W. Lara, A. Pelissari and M. Pereira et al., 1990. Action of plant regulators on growth, nutritional aspects, anatomy and the productivity of dry beans (Phaseolus vulgaris cv. Carioca). Anais da Escola Superior de Agricultura Luiz de Queiroz, 47: 11-28.
CrossRef - El-Desoki, E.R., S.I. Omara, Z.A. Ali and R.R. El-Masry, 1994. Effect of morphactin Cf125 on the growth and development of Vicia faba L. and Orobanche crenata Forsk. J. Agric. Sci. Mansoura Univ., 19: 1691-1699.
Direct Link - Kanazawa, A., A. Tozuka, S. Akimoto, J. Abe and Y. Shimamoto, 1998. Phylogenetic relationships of the mitochondrial genomes in the genus Glycine subgenus Soja. Genes Genet. Syst., 73: 255-261.
CrossRef - Mujoo, R., D.T. Trinh and P.K.W. Ng, 2003. Characterization of storage proteins in different soybean varieties and their relationship to tofu yield and texture. Food Chem., 82: 265-273.
CrossRef - Peterson, C.M., J.C. Williams and A. Kuang, 1990. Increased pod set of determinate cultivars of soybean, Glycine max, with 6-Benzylaminopurine. Bot. Gazette, 151: 322-330.
CrossRefDirect Link - Roat, C. and K.G. Ramawat, 2009. Morphactin and 2iP markedly enhance accumulation of stilbenes in cell cultures of Cayratia trifolia (L.) Domin. Acta Physiol. Plant., 31: 411-414.
CrossRef - Sztajer, H., E. Zboinska, I. Zbyryt, J. Kieleczawa, B. Lejczak and P. Kafarski, 1990. Preliminary studies on the mechanisms of action of phosphonic analogues of morphactins on plants and bacteria. Biol. Plant., 32: 28-34.
CrossRef - Wakhloo, J.L., 1975. Studies on the growth, flowering and production of female sterile flowers as affected by different levels of foliar potassium in Solanum sisvmbrifolium Lan. III- Interaction between foliar potassium and applied daminozide, chlormequat chloride and chlorflurenol-methyl. J. Exp. Cell Res., 12: 427-433.
CrossRef - Zayed, E.A., A.I. El-Zawily and E.S. Nofal, 1985. Studies on growth, productivity and some physiological aspects of hot pepper (Capsicum annum L. var. Red Cherry) II- The effect of the interaction between morphactin and NPK fertilization. J. Agric. Sci. Mansoura Univ., 10: 191-198.
Direct Link