
R. Angoshtari
Department of Agronomy and Plant Breeding, Campus of Agriculture and Natural Resources, University of Tehran, P.O. Box 31587-77871, Karaj, Iran
R. Tavakkol Afshari
Department of Agronomy and Plant Breeding, Campus of Agriculture and Natural Resources, University of Tehran, P.O. Box 31587-77871, Karaj, Iran
S. Kalantari
Department of Horticultural Sciences, Campus of Agriculture and Natural Resources, University of Tehran, P.O. Box 31587-77871, Karaj, Iran
M. Omidi
Department of Agronomy and Plant Breeding, Campus of Agriculture and Natural Resources, University of Tehran, P.O. Box 31587-77871, Karaj, Iran
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 4 | Page No.: 276-284







ABSTRACT
The present study describes, firstly, the effect of cultivar, photoenvironment and different abscisic acid (ABA) treatments on somatic embryogenesis in Brassica napus and secondly, describes the effects of ABA and drying rates on acquisition desiccation tolerance in cultivar Opera. In embryogenesis experiment, 20 days old cotyledon-derived calli were cultured in somatic embryo induction media with different concentrations of ABA (0, 0.5, 10 and 50 μM). These were incubated either in the dark or light. The calli were transferred to the same medium with either the same ABA or different ABA concentrations every 10 days. After 30 days the numbers of mature somatic embryos were counted. The results showed that light stimulated somatic embryo formation and maturation. Furthermore, differences in ABA response related to somatic embryogenesis were observed between cultivars. Also, embryos were likely more sensitive to ABA in days 1-10 and 21-30. For desiccation experiment, three ABA treatments were chosen from cultivar Opera. Then these embryos were cultured in germination medium without dehydration or dehydrated either with a fast dehydration rate or a slow dehydration rate before culturing in germination medium. The results showed that ABA treated embryos were more desiccation tolerant than non ABA treated embryos. It also showed that as the concentration of ABA increased, embryos tolerance to desiccation also increased. Moreover, slow drying rate was more beneficial for desiccation tolerance induction than fast drying in ABA treated embryos. In addition, ABA treated embryos had higher germination rates even when they were not dehydrated.
PDF Abstract XML References Citation
How to cite this article
R. Angoshtari, R. Tavakkol Afshari, S. Kalantari and M. Omidi, 2009. Effects of Abscisic Acid on Somatic Embryogenesis and Induction of Desiccation Tolerance in Brassica napus. Asian Journal of Plant Sciences, 8: 276-284.
DOI: 10.3923/ajps.2009.276.284
URL: https://scialert.net/abstract/?doi=ajps.2009.276.284
DOI: 10.3923/ajps.2009.276.284
URL: https://scialert.net/abstract/?doi=ajps.2009.276.284
INTRODUCTION
In plant tissue culture, regeneration occurs in two main ways; organogenesis (shooting) and somatic embryogenesis (Chawla, 2002). Organogenesis is the formation of a structure which in one end, is connected to the primary vascular tissues of the mother callus (Chawla, 2002). In contrast, somatic embryos are bipolar structures which are not connected to the primary vascular tissues of the mother calli (Chawla, 2002; Jimenez, 2005). For a number of reasons, somatic embryos are more desirable. First, because they are used as a model system in embryological studies in flowering plants; second, because of their practical application for large-scale vegetative propagation by using bioreactors; third, because they can be cryopreserved, which makes it possible to establish gene banks and fourth, because they are attractive targets for gene transformation (Shiota et al., 1999; Jimenez, 2005; George et al., 2008).
Many factors, like photoenvironment and Plant Growth Regulators (PGRs), influence regeneration either via somatic embryogenesis or shooting (Sun et al., 1998; Ghnaya et al., 2008; George et al., 2008). Depending on the plant species and cultivar, light (500-10000 Lux) seems to influence somatic embryo formation and shooting (Ascencio-Cabral et al., 2008; George et al., 2008). By now three kind of photoreceptors known as phytochrome (red and far red detector), cryptochrome (blue and UV-B detector) and phototropin (blue and UV-A detector) are thought to be involved in plant development (Ascencio-Cabral et al., 2008; George et al., 2008). Studies showed that somatic embryo formation increased in Carcia papaya in the red light and in Araujia sericifera in the light (Ascencio-Cabral et al., 2008; George et al., 2008). Also, it had been shown that growth and embryogenesis of carrot suspensions was inhibited, but not prevented by white (26 μmol/m2/sec) and blue (27.5 μmol/m2/sec) light, but the highest number of embryoids were obtained when they were kept in the dark, red and green light (George et al., 2008). The same study also suggested that red and blue light spectra could be used with advantage to manipulate carrot embryogenesis and embryo growth; as embryos growth, blue light also encouraged the formation of secondary embryos; red light hastened development towards the hearth stage; while ABA synthesis at this time was most pronounced under blue light (George et al., 2008).
Abscisic acid is shown to be important in both somatic embryogenesis and embryo (somatic and zygotic) maturation (Jimenez, 2005; Braybrook and Harada, 2008; Suzuki and McCarty, 2008; Wasilewska et al., 2008). Aspects of zygotic embryo maturation such as reserve products accumulation and acquisition of desiccation tolerance are shown to be influenced by ABA (Wakui et al., 1999; Gawronska et al., 2000; Finkelstein et al., 2002; Sreedhar et al., 2002; Schmidt et al., 2005). In zygotic embryos, it was reported that ABA levels decreased during embryo development whereas ABA sensitivity increased during the same period (Finkelstein et al., 1985). However, both endogenous ABA levels and sensitivity decreased during desiccation (Finkelstein et al., 1985). Studies showed that ABA level in somatic embryos are much lower than their zygotic counterparts (Gawronska et al., 2000; George et al., 2008). ABA treatments are shown to increase normal somatic embryo production (Gawronska et al., 2000; Chawla, 2002; Feher et al., 2003). ABA treatments also shown to stimulate the production of protein and oil reserves in somatic embryos to levels comparable to those found in their zygotic counterparts (Wilen et al., 1990; Schmidt et al., 2005). The effects of wounding, high salt concentrations, heavy metal ions or osmotic stress on inducing somatic embryos which is seen in diverse plant species, may be because of the effects of all this situations on endogenous ABA levels (Wilen et al., 1990; Finkelstein et al., 2002; Feher et al., 2003). In most species low levels of exogenous ABA treatments (0.1-1 μM) increased somatic embryo formation in early stages, whereas when more developed (after late hearth stage) even more ABA levels (100 μM) seem to be beneficial (George et al., 2008).
Desiccation is the final phase of normal embryonic development in most angiosperms and appears to be important in the transition from embryogeny to the ability to germinate and form normal seedlings (Finkelstein et al., 1985; Bomal and Tremblay, 1999; Bomal et al., 2002). Desiccation tolerant cells have some unique characteristics. These include the substitution of large vacuoles by plastids containing storage substances; protection of the integrity of the DNA; orderly dismantling of cytoskeletal elements; having the capability of microtubule reassembly; changes in overall protein secondary structure; accumulation of di and oligosaccharides and synthesis of especial proteins such as Late Embryogenesis Abundant (LEA) proteins (Golovina et al., 2001; Sreedhar et al., 2002; Faria et al., 2005; Berjak and Pammenter, 2008). Furthermore, the majority of these cells appear to be arrested in the G1 phase of the cell cycle (Golovina et al., 2001; Faria et al., 2005; Schmidt et al., 2006; Berjak and Pammenter, 2008). The main difference of somatic and zygotic embryos is their difference in responding to desiccation. Zygotic embryos of orthodox seeds are desiccation tolerant whereas somatic embryos, either orthodox or recalcitrant, are desiccation sensitive.
Desiccation studies in rapeseed (Brassica napus L.) are of great importance. It had been shown that, in oilseeds such as rapeseed, at least 10% of the major storage product of developing embryos, which is triacylglycerol is lost during the desiccation phase of seed development (Chia et al., 2005). So, a better understanding of this critical phase in seed maturation is needed if increasing in overall oil production of rapeseed is favored. Apart from this, it had been shown that desiccated somatic embryos can be preserved successfully for long periods of time without using cryoprotectants pretreatments and expensive programmable freezers (Shiota et al., 1999). So this simple method may provide a new tool for preservation of somatic embryos in gene banks.
In the present study, firstly, the effects of cultivar; light and different ABA treatments on somatic embryogenesis in rapeseed (Brassica napus L.) were studied. In this experiment, our main goal was to find the proper photoenvirnoment and dose and moment of application of ABA in related to somatic embryogenesis. Then the effects of different ABA treatments and different drying rates on acquisition of desiccation tolerance in Brassica napus (cultivar Opera) were studied. Here, we tried to propose the proper dose of ABA and drying rate for inducing desiccation tolerance in these somatic embryos.
MATERIALS AND METHODS
Plant materials: Three genotypes of rapeseed (Brassica napus L.) were used in this study; these were the winter cultivars Okapi and Opera and the spring cultivar PF7045/91. These seeds were given by the Oilseeds Research Department, Seed and Plant Improvement Institute, Karaj, Iran. They were harvested in 2007.
Chemicals: The ingredients of MS (Murashige and Skoog’s medium) basal medium, BA (N6-benzyladenine), 2, 4-D (2, 4-Dichlorophenoxyacetic acid), NAA (1-Naphthylacetic acid) and L-glutamine were products from Merck, Germany. Abscisic acid (ABA) was a product from Sigma-Aldrich, Canada.
Somatic embryogenesis: Seeds were surface sterilized with ethanol 70% for 1 min followed by sterilizing with 2.5% sodium hypochlorite for 15 min. They were washed 3 times in sterile distilled water and germinated in a jar containing MS medium without vitamins, myo-inositol and plant growth regulators, with 1.5% sucrose and 0.8% agar at a destiny of 10 seeds per jar. These jars were cultured in dark at 22°C for 3 days and after that, they were transferred to fluorescent light (3000 Lux) generated by fluorescent lamps (Day light, 20 W, 511, PARS, IRAN) at 22°C and left to grow for 7 more days. The cotyledon sections were transferred to MS basal media containing 1 mg L-1 NAA; 1 mg L-1 2, 4-D; 1 mg L-1 BA, 3.4% sucrose and 0.7% agar-agar (Sun et al., 1998). The pH of the media was adjusted to 5.8 prior to autoclaving at 121°C for 20 min. Glass Petri dishes (12 cm) containing these media and explants were cultured in the dark at 22°C for 10 days. Each Petri dish contains 5 pieces of cotyledon (each cotyledon was cut transversely into two equal pieces). After 10 days, the calli formed was subcultured in the same media and spend 10 more days in the same conditions. The 20 days old calli were then transferred to MS basal media with 200 mg L-1 L-glutamine, 4.5 mg L-1 BA, 0.2 mg L-1 NAA, 3% sucrose, 0.7% agar-agar (somatic embryo induction medium) and different ABA concentrations (Table 1). The pH was adjusted to 5.8. Glass Petri dishes (12 cm) containing these media and calli were cultured in either dark or fluorescent light (3000 Lux) generated by fluorescent lamps (Day light, 20 W, 511, PARS, IRAN) at 22°C for the next 30 days. The calli were subcultured in days 11 and 21 (Table 1). After 30 days, mature somatic embryos were counted.
Desiccation: Mature somatic embryos of cultivar Opera, which were formed in ABA treatments S1, S4 and S12, were chosen for desiccation treatments. Desiccation treatment 1 (D1) was a treatment without desiccation. In D1, embryos were directly transferred to Petri dishes containing 1/2MS basal media without PGRs (Sun et al., 1998). These Petri dishes were incubated at 22°C in the light (3000 lux) generated by fluorescent lamps (Day light, 20 W, 511, PARS, IRAN). Desiccation treatment 2 (D2) was fast desiccation treatment. In D2, embryos were dried within 4 h by placing Petri dishes containing these embryos without covers in an air-flow cabinet (Wolkers et al., 1999).
| Table 1: | Abscisic acid (ABA) treatments used for somatic embryogenesis in Brassica napus L. These treatments included dose and application time of ABA treatments |
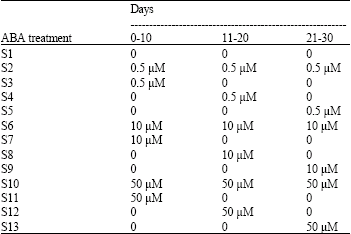 | |
| S1, S4 and S12 were also used for desiccation | |
The Relative Humidity (RH) of the air flow was approximately 40%, as monitored with a hygroscope (model Hygroskop DT, Rotronic, Zurich, Switzerland). In desiccation treatment 3 (D3) embryos were dried through a series of desiccators in which Relative Humidity (RH) was kept constant using a saturated solution of K2SO4 (97%), NaCl (75%) and K2CO3.1.5H2O (40%) (Johnson-Flanagan et al., 1992). Desiccators were kept in dark at 22°C. In this desiccation treatment, embryos were transferred from a desiccator at a higher RH to one at a lower RH, every 72 h. In order to avoid imbibitional damage, before rehydration, desiccated embryos from D2 and D3 were transferred to a desiccator in which the RH was kept 97% for 4 h. After that, these embryos were cultured in 1/2MS basal media without PGRs (Sun et al., 1998), in the light (3000 lux) generated by fluorescent lamps (Day light, 20 W, 511, PARS, IRAN) at 22°C. These embryos were assessed for desiccation recovery and normal development at 4 weeks after rehydration. Only those embryos that showed both shoot and root meristem growth were scored as surviving (Johnson-Flanagan et al., 1992).
Data analysis: In somatic embryogenesis experiment, treatments consisted of three cultivars (PF7045/91, Opera and Okapi), two photoenvironments (light and dark) and thirteen ABA treatments (Table 1). In this experiment, mature somatic embryos in each Petri dish were calculated. For desiccation survival, survival percent were calculated. Twenty five embryos from each ABA treatment and replicate were used for each desiccation treatment. Here the treatments consisted of three ABA treatments (S1, S4 and S12) and three desiccation treatments (D1, D2 and D3). These experiments (somatic embryogenesis and desiccation) were arranged in two factorial experiments as completely randomized designs (Table 2, 3).
| Table 2: | Analysis of variance for the effects of cultivar, light and ABA treatments on somatic embryogenesis in Brassica napus |
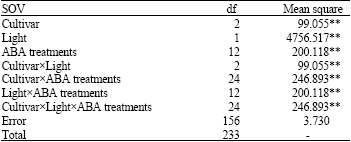 | |
| **Level of significance = 0.01. Cultivar, light and ABA treatments have significant effects on somatic embryo formation and maturation in Brassica napus | |
| Table 3: | Analysis of variance for the effects of ABA (S1, S4 and S12) and drying rate (D1, D2 and D3) treatments on germination percentage of mature somatic embryos of Brassica napus cultivar Opera |
 | |
| **Level of significance = 0.01. Both ABA and desiccation treatments significantly affected the induction of desiccation tolerance in Brassica napus cultivar Opera somatic embryos | |
In each experiment, each treatment combination was replicated three times. Statistical analyses were done using SAS (version 9.00) analysis of variance procedures (Table 2, 3). Means of main effects interactions were compared by Tukey’s Honestly Significant Difference Test of MSTATC software (Fig. 3, 4).
RESULTS AND DISCUSSION
Somatic embryogenesis: All factors (photoenvironment, cultivar and ABA treatments) had significant effects on somatic embryo formation and maturation (Table 2). In somatic embryogenesis experiment, although some embryo like structures were observed in the dark (Fig. 1a), no mature somatic embryos were produced in this photoenvironment condition. In contrast, somatic embryos were formed and matured only in the light (Fig. 2). Also, in cultivars Okapi and Opera under light condition, shooting were observed in media S7 and S13 (Fig. 1B, C). No shooting were observed in the dark in these media, therefore, regeneration (either via somatic embryogenesis or shooting) is somehow light dependent. Actually, light was shown to affect regeneration in some plant species (Ascencio-Cabral et al., 2008; George et al., 2008). There are some reports on the involvement of light on phytohormone synthesis (George et al., 2008). This may explain how light influences somatic embryo formation and organogenesis in plants.
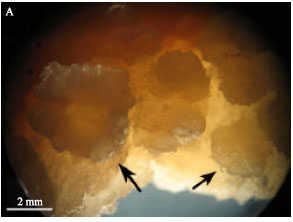 | |
| Fig. 1: | (A) Somatic embryos of Brassica napus formed in the dark, (B) shoots of Brassica napus formed from calli in the light and (C) shoots growth after spending 5 weeks in 1/2MS, hormone free media in the light |


The effects of ABA on shooting and somatic embryogenesis were shown to be cultivar dependent. As mentioned before, in cultivars Okapi and Opera shooting were observed in media S7 and S13. It is likely that 10 μM of ABA in earlier stages of development (days 1-10) and 50 μM of ABA in later stages (days 21-30) induced shoot formation in these 2 cultivars.
Earlier studies showed that ABA is beneficial for embryo formation and maturation (Finkelstein et al., 1985; Gawronska et al., 2000; Finkelstein et al., 2002; Chawla, 2002; Jimenez, 2005; George et al., 2008).
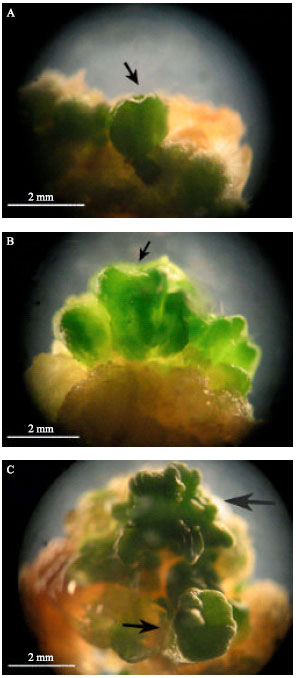 | |
| Fig. 2: | Brassica napus somatic embryo formation in the light. (A) Hearth stage somatic embryo, (B) torpedo stage somatic embryo and (C) cotyledonary stage somatic embryos with normal cotyledon number (small arrow) and abnormal cotyledon number (big arrow). Mature somatic embryos formed only in the light |
But to our knowledge this was the first time that different ABA concentrations in different embryogenesis stages were used for embryogenesis studies in Brassica napus cultivars. Although earlier studies showed that in most species low levels of exogenous ABA treatments (0.1-1 μM) used in early stages, induced somatic embryo formation, here in cultivars PF7045/91 and Opera somatic embryo formation were observed when calli were exposed to much higher concentrations of ABA [10 μM (S7) and 50 μM (S11)] in early stages of somatic embryo formation (Fig. 3). Of course, more studies on endogenous levels of ABA are needed in these cases. Furthermore, in the case of somatic embryogenesis, PF7045/91 showed the highest rate of mature embryo formation when it was exposed to 0.5 μM of ABA for the whole 30 days (S2) or 10 μM of ABA in days 21-30 (S9). In days 1-10, PF7045/91 responded to all concentrations of ABA (S3, S7 and S11 respectively), but its response was not significantly different (Fig. 3). No responses, related to somatic embryogenesis, were observed in days 11-20 in this cultivar and in days 21-30, the highest response occurred when the embryos where exposed to 10 μM of ABA (S9). This response would reduce significantly when the concentration of ABA declined to 0.5 μM (S5). Cultivar Okapi had the highest somatic embryo formation rate when exposed to 0.5 μM of ABA in days 1-10 or 21-30 and also when exposed to 50 μM of ABA in days 11-20 (S3, S5 and S12, respectively). These responses were not significantly different, so it is possible that these embryos were more sensitive to ABA in their first and last 10 days of development. On the other hand, when exposed for the whole 30 days to the low concentration (0.5 μM) of ABA (S2), Okapi calli produced a small amount of somatic embryos. It did not produce somatic embryos in the absence of ABA (S1) and produced mature somatic embryos in only 4 media of 13 media. These might show that ABA sensitivity in Okapi is higher than the other two cultivars. Cultivar Opera showed responses to all ABA concentrations in days 1-10 (S3, S7 and S11). These responses were not significantly different (Fig. 3). In days 11-20, this cultivar showed the best response, when exposed to 0.5 and 50 μM of ABA (S4 and S12, respectively). These responses were not significantly different either. In days 21-30, this cultivar produced mature somatic embryos when exposed to 0.5 and 10 μM of ABA (S5 and S6, respectively). Also, Opera did not produced any mature somatic embryos when its calli were exposed to low concentration (0.5 μM) of ABA for the whole 30 days (S2), but it was the only cultivar that produced mature somatic embryos in the absence of exogenous ABA (S1). These might show that ABA endogenous level in Opera is higher than the other two cultivars.
To make a long story short, three main points from this part should be mentioned. (1) Somatic embryos of Brassica napus (cultivars Opera, Okapi, PF7045/91) matured in the light but not in the dark. (2) The effect of ABA on somatic embryogenesis in Brassica napus is cultivar dependent; Okapi somatic embryos seemed to be more sensitive to ABA whereas Opera somatic embryos seemed to have higher levels of endogenous ABA.
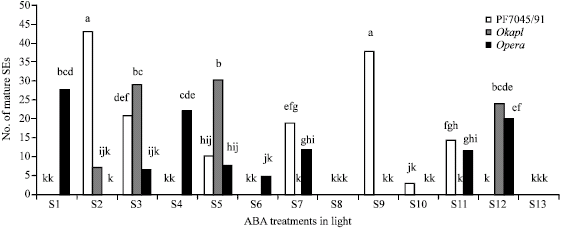 | |
| Fig. 3: | Number of mature Somatic Embryos (SEs) formed in different ABA treatments in the light, in three rapeseed cultivars PF4045/91, Okapi and Opera. Values represent the mean of three replicates. Means followed by the same letter are not significantly different (Tukey’s honestly significant difference test, probability level = 0.05). Cultivar Opera was the only cultivar which produced mature somatic embryos in the control [ABA free medium (S1)]. Interestingly, this cultivar produced the highest number of somatic embryos in the same ABA treatment (S1). The highest number of somatic embryo production between the three cultivars, belongs to cultivar FP7045/91 in ABA treatments S2 and S9. Cultivar Okapi produced the highest number of somatic embryos in S3 and S5 |
(3) It is likely that embryos of winter cultivars (like Opera and Okapi) are more sensitive or have higher endogenous levels of ABA than spring cultivars (here PF7045/91). Of course more studies on endogenous levels of ABA in different Brassica napus cultivars are needed for proving these hypotheses.
Desiccation: Since cultivar Opera was the only cultivar which produced mature somatic embryos in the control [ABA free medium (S1)], it was used for desiccation experiment (Fig. 3). Both ABA and desiccation treatments had significant effects on induction of desiccation tolerance in Opera (Table 3). Unlike previous studies in this field, here different ABA concentrations were used for desiccation tolerance induction. In this experiment it had been shown that in the presence of exogenous ABA, germination percentage of somatic embryos increased significantly, no matter they were dehydrated or not (Fig. 4, 5). For example, comparing to the control (S1), when ABA was used (either 0.5 μM or 50 μM), germination percentage increased nearly 15-25% in those embryos which were not dehydrated (Fig. 4). When mature somatic embryos were not dehydrated, no significant difference in germination percentage was observed between different ABA concentrations (S4 and S12). In contrast, when they were dehydrated (either by fast drying or slow drying), significant difference were observed between ABA concentrations (Fig. 4).
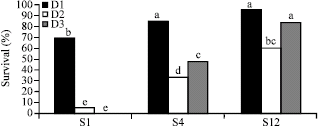 | |
| Fig. 4: | Somatic embryo survival percentage after ABA treatments, desiccation treatments and rehydration in rapeseed (Brassica napus cv. Opera). Values represent the mean of three replicates. Means followed by the same letter are not significantly different (Tukey’s honestly significant difference test, probability level = 0.05). When exogenous ABA was used (S4 and S12), germination (survival) percentage of somatic embryos increased significantly, no matter they were dehydrated or not. Also, in the presence of ABA slow drying rate (D3) resulted in higher germination percentage than fast drying rate (D2) |
Comparing to the control (S1), germination percentage of fast dried somatic embryos increased 28% when 0.5 μM ABA (S4) was used, but it increased even higher (nearly 55%) when 50 μM (S12) ABA was used. In the case of slow drying rate, germination percentage of these somatic embryos increased 48 and 84%, respectively, when 0.5 and 50 μM ABA were used. As shown in these cases, higher concentration of ABA (S12) was more beneficial on inducing desiccation tolerance than lower ABA concentration (S4).
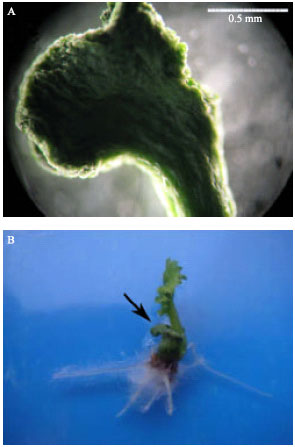 | |
| Fig. 5: | (A) Cotyledon of a desiccated somatic embryo of Brassica napus L. cultivar Opera. (B) Somatic embryo growth 5 weeks after rehydration in 1/2MS, hormone free media in the light. The arrow shows the cotyledon |
Studies shown that the external surface of the desiccation-tolerant embryos was uniformly shriveled due to severe desiccation and their internal tissue system was well preserved, whereas in desiccation-sensitive ones, dehydration caused tearing of the epidermis and collapse of the internal tissue system (Wakui et al., 1999). After rehydration, desiccation tolerant embryos recovered and continue normal growing but desiccation sensitive ones did not recover or remained shriveled (Wakui et al., 1999).
Researchers have shown that the rate of drying is an important factor in initiating the mechanism of desiccation tolerance in plant organs (Bomal and Tremblay, 1999; Wolkers et al., 1999; Wesley-Smith et al., 2001; Bomal et al., 2002; Sreedhar et al., 2002; Wang et al., 2002). Here, slow drying rate resulted in higher germination percentage in the presence of ABA. In fact, it is possible that during fast drying time is lacking for the synthesis of specific protective compounds (such as LEA proteins and certain kind of sugars) which are related to acquisition of desiccation tolerance (Wolkers et al., 1999; Wang et al., 2002). When 0.5 μM ABA (S4) was used, in slow dried somatic embryos germination percentage was increased nearly 15% comparing to fast dried ones (Fig. 4). Furthermore, in slow dried somatic embryos which were formed in S12 (50 μM ABA) medium, germination percentage increased even higher (24%).
In the absence of ABA, slow drying protocol led to complete intolerance to desiccation; whereas some survival occurred in the faster drying rate (Fig. 4). It is likely that another effect of ABA appears in such cases; if slow drying be considered as a form of accelerated ageing treatment; ABA might have reduced cellular damage by curtailing metabolic activity (Sreedhar et al., 2002). As a result, somatic embryos could be more sensitive to slow drying when they are grown in the absence of ABA (Sreedhar et al., 2002).
To summarize the main points of this part these points should be mentioned. (1) Exogenous ABA improved somatic embryo germination percentage no matter these embryos was dehydrated or not. (2) When dehydrated (no matter fast or slow), higher level of exogenous ABA was more effective on somatic embryo germination percentage than lower level of ABA. (3) When exogenous ABA was used, slow drying rate improved somatic embryo germination percentage more than fast drying rate.
In the recent years, researchers have found relationships between two novel aspects of plant development which are totipotency via somatic embryogenesis and maturation phase of zygotic embryo development in seed plants (Braybrook and Harada, 2008). Different plant growth regulators such as abscisic acid (ABA), Gibberellic Acid (GA), ethylene and auxin are involved in both of these processes (Gawronska et al., 2000; Finkelstein et al., 2002; Feher et al., 2003; Braybrook and Harada, 2008; Suzuki and McCarty, 2008; Berjak and Pammenter, 2008). In addition, the application of polyamines, jasmonate and brassinosteroids has proved to be useful for initiating somatic embryogenesis in many plant species (George et al., 2008). When it comes to desiccation tolerance, as shown here, ABA has a positive effect on this phenomenon, but the effect of other plant growth regulators are yet to be understood. All in all, although so many advances have been made in the fields of embryogenesis and desiccation tolerance, if we want to have a better understanding of these processes there are still so many questions to be answered in these fields.
ACKNOWLEDGMENT
This study was funded by the grant number of 7101027/1/002 from the University of Tehran, Vice Chancellor for Research. The authors wish to express their gratitude to the Department of Agronomy and Plant Breeding, University of Tehran, Iran, for supporting this work.
REFERENCES
- Ascencio-Cabral, A., H. Gutierrez-Pulido and B. Rodriguez-Garay, 2008. Plant regeneration of Carica papaya L. through somatic embryogenesis in response to light quality, gelling agent and phloridzin. Sci. Hort., 118: 155-160.
CrossRef - Berjak, P. and N.W. Pammenter, 2008. From Avicennia to Zizania: Seed recalcitrance in perspective. Ann. Bot., 101: 213-228.
CrossRefPubMedDirect Link - Bomal, C., L. Van Quy and F.M. Tremblay, 2002. Induction of tolerance to fast desiccation in black spruce (Picea mariana) somatic embryos: Relationship between partial water loss, sugars and dehydrins. Physiol. Plant, 115: 523-530.
CrossRef - Braybrook, S.A. and J.J. Harada, 2008. LECs go crazy in embryo development. Trends Plant Sci., 13: 624-630.
CrossRef - Chia, T.Y.P., M.J. Pike and S. Rawsthorne, 2005. Storage oil breakdown during embryo development of Brassica napus (L.). J. Exp. Bot., 56: 1285-1296.
CrossRef - Faria, J.M., J. Buitink, A.A.M. van Lammeren and H.W.M. Hilhorst, 2005. Changes in DNA and microtubules during loss and re-establishment of desiccation tolerance in germinating Medicago truncatula seeds. J. Exp. Bot., 56: 2119-2130.
CrossRef - Feher, A., T.P. Pasternak and D. Dudits, 2003. Transition of somatic plant cells to an embryogenic stage. Plant Cell Tissue Org. Cult., 74: 201-228.
Direct Link - Finkelstein, R.R., S.S.L. Gampala and C.D. Rock, 2002. Abscisic acid signaling in seeds and seedlings. Plant Cell, 14: S15-S45.
Direct Link - Gawronska, H., W. Burza, E. Bolesta and S. Malepszy, 2000. Zygotic and somatic embryos of cucumber (Cucumis sativus L.) substantially differ in their levels of abscisic acid. Plant Sci., 157: 129-137.
PubMed - George, E.F., M.A. Hall and G.J. de Klerk, 2008. Plant Propagation by Tissue Culture. 3rd Edn., Springer, Dordrecht, Netherlands, Pages: 501.
CrossRefDirect Link - Ghnaya, A.B., G. Charles and M. Branchard, 2008. Rapid shoot regeneration from thin cell layer explants excised from petioles and hypocotyles in four cultivars of Barssica napus L. Plant Cell Tissue Org. Cult., 92: 25-30.
CrossRef - Golovina, E.A., F.A. Hoekstra and A.C. van Aelst, 2001. The competence to acquire cellular desiccation tolerance is independent of seed morphological development. J. Exp. Bot., 52: 1015-1027.
PubMed - Jimenez, V.M., 2005. Involvement of plant hormones and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regul., 47: 91-110.
CrossRef - Schmidt, Th., D.M. Tucker, E.B. Cahoon and W.A. Parrott, 2005. Towards normalization of soybean somatic embryo maturation. Plant Cell Rep., 24: 383-391.
CrossRef - Schmidt, Th., A. Ewald, M. Seyring and A. Hohe, 2006. Comparative analysis of cell cycle events in zygotic and somatic embryos of Cyclamen persicum indicates strong resemblance of somatic embryos to recalcitrant seeds. Plant Cell Rep., 25: 643-650.
CrossRef - Shiota, H., K. Tachibana, K. Watabe and H. Kamada, 1999. Successful long-term preservation of abscisic-acid-treated and desiccated carrot somatic embryos. Plant Cell Rep., 18: 749-753.
CrossRefDirect Link - Sreedhar, L., W.F. Wolkers, F.A. Hoekstra and J.D. Bewley, 2002. In vivo characterization of the effects of abscisic acid and drying protocols associated with the acquisition of desiccation tolerance in alfalfa (Medicago sativa L.) somatic embryos. Ann. Bot., 89: 391-400.
CrossRef - Sun, M., H. Kieft and A.A.M. Lammeren, 1998. Cotyledon-derived diploid and haploid protoplast culture and diploid plant regeneration in Brassica napus cv. ' topas. Can. J. Bot., 76: 530-541.
CrossRefDirect Link - Suzuki, M. and D.R. McCarty, 2008. Functional symmetry of the B3 network controlling seed development. Curr. Opin. Plant Biol., 11: 548-553.
CrossRef - Wakui, K., Y. Takahata and N. Kaizuma, 1999. Scanning electron microscopy of desiccation-tolerant and sensitive microspore-derived embryos of Brassica napus L. Plant Cell Rep., 18: 595-600.
CrossRef - Wang, X.J., C.S. Loh, H.H. Yeoh and W.Q. Sun, 2002. Drying rate and dehydrin synthesis associated with abscisic acid-induced dehydration tolerance in Spathoglottis plicata orchidaceae protocorms. J. Exp. Bot., 53: 551-558.
PubMed - Wasilewska, A., F. Vlad, C. Sirichandra, Y. Redko and F. Jammes et al., 2008. An update on abscisic acid signaling in plants and more. Mol. Plant, 1: 198-217.
CrossRefPubMedDirect Link - Wesley-Smith, J., N.W. Pammenter, P. Berjak and C. Walters, 2001. The effects of two drying rates on the desiccation tolerance of embryonic axes of recalcitrant jackfruit (Artocarpus heterophyllus Lamk.) seeds. Ann. Bot., 88: 653-664.
CrossRefDirect Link - Wilen, R.W., R.M. Mandel, R.P. Pharis, L.A. Holbrook and M.M. Moloney, 1990. Effects of abscisic acid and high osmoticum on storage protein gene expression in microspore embryos of Brassica napus. Plant Physiol., 94: 875-881.
CrossRefDirect Link - Wolkers, W.F., F.A.A. Tetteroo, M. Alberda and F.A. Hoekstra, 1999. Changed properties of the cytoplasmic matrix associated with desiccation tolerance of dried carrot somatic embryos. An in Situ fourier transform infrared spectroscopic stud. Plant Physiol., 120: 153-163.
CrossRefDirect Link