
Weili Yu
College of Soil and Water Conservation, Beijing Forest University, Beijing 100083, China
Lee D. Hansen
Department of Chemistry and Biochemistry, Brigham Young University, Provo, Utah 84602, USA
Wenying Fan
Inner Mongolia Academy of Forest and Science, Huhhot, Inner Mongolia 010010, China
Wenyi Zhao
Inner Mongolia Academy of Forest and Science, Huhhot, Inner Mongolia 010010, China
E. Durant McArthur
USDA Forest Service, Rocky Mountain Research Station,Shrub Sciences Laboratory, Provo, Utah 84606, USA
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 67-72







ABSTRACT
Growth and respiratory characteristics of Caragana korshinskii from Wushen and two different seed sources of C. davazamcii from Helinger and Yijinhuole, all grown at the same conditions, were determined by measuring metabolic heat and CO2 production rates by isothermal calorimetry at 5°C intervals from 10 to 40°C. Substrate carbon conversion efficiencies and growth (anabolic) rates were calculated from the measured data. Differences in substrate carbon conversion efficiency, respiratory rates and temperature responses of respiratory rates among the three accessions all contribute to produce differing temperature responses of growth rate. Planting seeds from these seed sources outside their native ranges will probably not be successful because of the differences in temperature adaptation.
PDF Abstract XML References Citation
How to cite this article
Weili Yu, Lee D. Hansen, Wenying Fan, Wenyi Zhao and E. Durant McArthur, 2008. Adaptation of Growth and Respiration of Three Varieties of Caragana to Environmental Temperature. Asian Journal of Plant Sciences, 7: 67-72.
DOI: 10.3923/ajps.2008.67.72
URL: https://scialert.net/abstract/?doi=ajps.2008.67.72
DOI: 10.3923/ajps.2008.67.72
URL: https://scialert.net/abstract/?doi=ajps.2008.67.72
INTRODUCTION
Temperature is one of the most significant determinants of plant distribution. Plants adapt their growth to their niche by adapting their respiratory metabolism to the temperature pattern of the environment (Criddle et al., 2005). Altering rates and the temperature dependence of metabolism and metabolic efficiency may all be important in adapting growth characteristics to the local climate. This study compares growth and respiration characteristics of Caragana korshinskii from Wuhen and C. davazamcii from Helinger and Yijinhuole, Inner Mongolia. These shrubs are used for combating desertification, for water and soil conservation (Zhang et al., 2002; Niu, 2002), as feed for cattle and wild animals in deserts and grasslands and are the main shrub used for rangeland restoration in Inner Mongolia.
C. korshinskii and C. davazamcii are distributed from the west to the middle of Inner Mongolia. C. davazamcii is a species of the eastern Gobi, occurring in the steppe desert and desert steppe zones on sand lands and Loess hills of the Ordos Plateau and northern Loess Plateau where it is a dominant species of shrub vegetation. C. korshinskii is a species of the southern Alashan and western Ordos where it occurs in fixed and semi-fixed dunes of the steppe desert and the typical desert zone. The zonal distribution of Caragana forms an obvious geographic substituted distribution from east to west or from north to south (Zhao, 2005). C. korshinskii and C. davazamcii are distinguished by characteristics of flowers, seeds, leaves and especially bark and height. C. korshinskii has yellow-gold, lustrous bark and grows taller (300-500 cm, Zhao, 2005) than C. davazamcii (50-150 cm, Zhang et al., 2002). The morphology of leaves of Caragana show a regular geographic gradient, leaf area becomes smaller and well developed palisade tissue in the intercellular space also tends to become smaller from east to west (Yan et al., 2002). Drought tolerance also increases from east to west. There are disagreements on the taxonomy of C. davazamcii and C. korshinskii; Sanchir (1974), Fu (1977), Liu (1984) and Zhao (1990) argue that they are independent species, Yakovlev (1988) argues that they are the same species and Zhang and Zhu (2004) argue that C. davazamcii is a variant of C. korshinskii. For nomenclature purposes only, in this study C. korshinskii and C. davazamcii are considered to be independent species.
In this study, CO2 production rates (RCO2) and metabolic heat rates (Rq) are measured as a function of temperature by calorespirometry on young, growing leaf tissue of C. korshinskii and C. davazamcii. These data allow calculation of growth (or anabolic) rates and substrate carbon conversion efficiencies as functions of temperature. Selection of accessions for restoration can then be done by matching growth responses to local temperature patterns to maximize the likelihood of success. This laboratory method cannot completely replace field experiments, but can save much time and effort and make field experiments much more efficient. Determining the relationship between metabolism and growth and thence the mechanism of adaptation of different accessions to environmental temperature patterns will provide better understanding for use of these shrubs.
MATERIALS AND METHODS
This study was conducted at the Shrub Sciences Laboratory of the USDA Forest Service and the Department of Chemistry and Biochemistry of Brigham Young University, both in Provo, Utah, USA.
Theory: Plant growth depends on photosynthesis to supply substrate carbon, but under many conditions the limiting factor for growth is not photosynthate, but micronutrients and/or the rate of utilization of photosynthate by respiration (Vitousek and Howarth, 1991; Hansen et al., 1998). Further, because the processes of photosynthesis and respiration are separated in time, photosynthesis rate cannot be used to predict growth rate. However, growth rate of vegetative tissue is equal to the anabolic rate in respiration and calorespirometry can be used to obtain both the anabolic and catabolic rates and thus the growth rate and substrate carbon conversion efficiency (Hansen et al., 2004, 2005). Catabolism of photosynthate provides the energy to drive anabolism and anabolism produces structural biomass from photosynthate. Because nearly all of the heat of respiration is produced by catabolism, the metabolic heat rate (Rq) measures the rate of catabolism;
CS + O2 + xADP + xPi → CO2 + H2O + xATP + heat | (1) |
Where, Cs is the carbon substrate (assumed to be carbohydrate in this reaction as written). ATP generated during catabolism supplies energy to drive anabolism,
CS + yATP → zCBio + (1-z)CO2 + yADP + yPi | (2) |
Where:
| Cs= | Carbon substrate which is assumed to be carbohydrate in this reaction as written |
| Cbio= | New growth, i.e., the rate of formation of Cbio is equal to the rate of growth or development |
Heat energy stored in ATP by the catabolic reaction is released in the anabolic reaction, but, because the ATP reactions cancel in the overall process, this heat can be assigned to either reaction and the calculations are greatly simplified if it is assigned to catabolism.
The sum of the catabolic and anabolic reactions is:
CS + aO2 + (N,P,K, etc.) → εCBio +(1-ε)CO2 | (3) |
Where, ε is the substrate carbon conversion efficiency, i.e., the fraction of Cs converted to CBio.
Equations relating mass-specific growth (anabolic) rate (RSG) to ε and to measured RCO2 and Rq have been derived (Hansen et al., 2005, 2004);
RSG/RCO2 = ε/(1-ε) or RSG = RCO2 [ε/(1-ε)] | (4) |
and
RSG= [-ΔHCO2 RCO2-Rq]/ΔHB | (5) |
Where, ΔHB and ΔHCO2 are, respectively, enthalpy changes for the difference in heats of combustion of substrate and biomass and for the heat of combustion of substrate to CO2. For closely related tissues, ΔHB can be regarded as a constant and for vegetative tissue similar to that used in this study, can be assumed equal to +30 kJ Cmol-1 (Lamprecht, 1999). From Thornton`s rule, ΔHCO2 is approximately equal to -455(1-γs/4)kJ Cmol-1 where γs is the chemical oxidation state of carbon in the substrate (e.g., for carbohydrate γs = 0, for lipid γs ≈ -1.8 and for protein γs ≈ -1.0). Specifically for carbohydrate, ΔHCO2 is -470 kJ Cmol-1; so in the absence of evidence for other substrates, ΔHCO2 is assumed equal to -470 kJ Cmol-1.
Combining Eq. 4 and 5 to eliminate RSG gives an equation for ε
ε/(1-ε) = [(-Rq/RCO2) -ΔHCO2]/ΔHB or
Rq/RCO2=-ΔHCO2-ΔHB[ε/(1-ε)] | (6) |
Determination of anabolic and catabolic rates and ε as functions of temperature for different genotypes grown under the same conditions allows determination of the influence of genotype and thus of natural selection by environmental temperature on these characteristics.
Materials: Seeds of Caragana korshinskii were collected near and Northwest of Wushen, Inner Mongolia and
| Table 1: | Results of linear least squares fitting (Excel, linest function) of plots of average values of RCO2 versus Rq at 10, 15, 20, 25, 30, 35 and 40°C. Uncertainties are given as the standard deviation |
 | |
| a: The substrate carbon conversion efficiency ε was calculated with equation 6 assuming ΔHCO2 = -470 kJ Cmol-1 and ΔHB = +30 kJ Cmol-1, *: Indicates these values differ significantly from the others in the column | |
seeds of Caragana davazamcii were collected near Helinger and east Yijinhuole, Inner Mongolia (Table 1). Seeds were planted in small containers with potting soil and grown in a greenhouse to a seedling height of 10-15 cm. Stems with attached leaves from the top branch were used for the calorespirometric measurements.
Measurement method: Hart Scientific Model 7707 and Calorimetry Sciences Corporation Model 4100 calorimeters, each with three 1 mL sample and one reference ampule, w ere used in isothermal mode. Seventy to 110 mg fresh tissue was placed into a sample ampule and the ampule inserted into the calorimeter. The calorimeter was set to a selected temperature and allowed to equilibrate for 20-30 min to obtain a steady-state metabolic heat rate which was obtained during the last 5-10 min of this period. The ampule was then taken out and a 40 μL vial of 0.4 M NaOH placed in the ampule with the tissue, the ampule was replaced in the calorimeter and the heat rate measured again. The heat rate from this second measurement is the sum of metabolic heat and heat from NaOH reacting with CO2,
CO2 (g) + 2OH-1(aq) = CO3 2–(aq) + H2O(aq) ΔH = -108.5 kJ mol-1 | (7) |
After removing the vial of NaOH, the metabolic heat rate is measured again. The metabolic heat rates from the first and third measurements are averaged and subtracted from the heat rate from the second measurement to obtain the heat rate for formation of carbonate (in μW or μJ sec-1). Dividing this rate by 108.5 μJ nmol-1 gives the production rate (RCO2) of CO2 in nmol sec-1. Averaging the two measurements of metabolic heat rate corrects for any small drift in the rate during the measurement period. This protocol was repeated at 10, 15, 20, 25, 30, 35 and 40°C for each seed source. A given tissue sample was measured in sequence at 20, 15 and 10°C, at 20, 25 and 30°C, or at 30, 35 and 40°C. To verify that sample history had no effect, additional samples were measured at only one temperature. Because the calorimeters measure the heat rates from three samples simultaneously, there were three to nine replicates for each species and seed source, with each ampule being a unit. Dry Weights (DW) were obtained after drying the samples for 24 h in a vacuum oven at 70-80°C. Metabolic heat and CO2 rates were divided by the dry weights to obtain mass-specific rates.
RESULTS
Figure 1 and 2, respectively show the average heat and CO2 production rates versus temperature. These plots are not linear when plotted on Arrhenius axes, i.e., ln(rate) versus reciprocal absolute temperature. The metabolic heat and CO2 rates have similar responses to temperature, but the responses differ among the three accessions. At temperatures below 25°C, Rq and RCO2 of the three accessions are comparable. At temperatures above 25°C, Rq and RCO2 of Yijinhuole are higher than those of Helinger, which are higher than those of the Wushen variety.
To determine whether the substrate carbon conversion efficiency (ε) changed with temperature, plots of RCO2 versus Rq were constructed and fit to a linear equation by least squares. The results are shown in Table 1. If such a plot is linear and has a zero intercept, it
f1
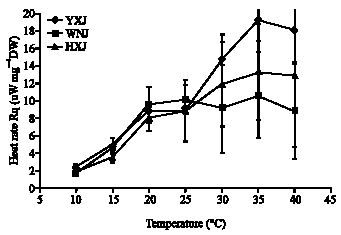 | |
| Fig. 1: | Metabolic heat rates (μW or μJ sec-1 mg-1 dry weight) of leaf tissue from Caragana korshinskii from Wushen (WNJ) and C. davazamcii from Helinger (HXJ) and from Yijinhuole (YXJ), Inner Mongolia. Error bars show the 95% confidence intervals. The scatter is largely due to differing responses of tissue samples since the instrument scatter is <5% |
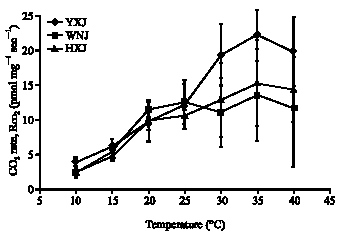 | |
| Fig. 2: | CO2 production rates of leaf tissue from Caragana korshinskii from Wushen (WNJ) and C. davazamcii from Helinger (HXJ) and from Yijinhuole (YXJ), Inner Mongolia. Error bars show the 95% confidence intervals. The scatter is largely due to differing responses of tissue samples since the instrument scatter is <10% |
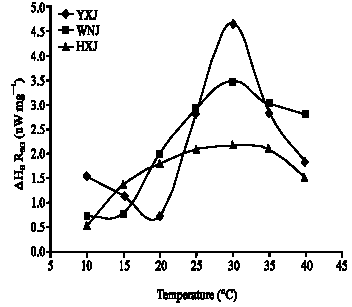 | |
| Fig. 3: | Growth rates calculated from average metabolic heat and CO2 production rates of leaf tissue from Caragana korshinskii from Wushen (WNJ) and C. davazamcii from Helinger (HXJ) and from Yijinhuole (YXJ), inner Mongolia. The temperature ranges estimated from the growth curves indicate the range for optimal growth |
can be concluded that ΔHCO2, ΔHB and ε do not change with temperature (Eq. 6) and thus that the substrate and biomass composition and catabolic-anabolic coupling do not change with temperature over the linear range of the plot. The intercepts in Table 1 are near zero and the plots are linear, showing that none of these characteristics change significantly with temperature.
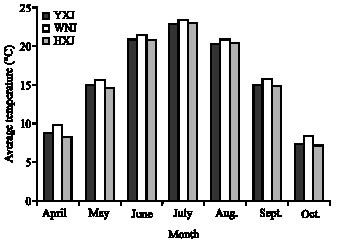 | |
| Fig. 4: | Monthly average temperatures for Wushen (WNJ), Helinger (HXJ) and Yijinhuole (YXJ), Inner Mongolia |
Because it is highly unlikely that ΔHCO2 or ΔHB differ among the accessions, we conclude that the substrate carbon conversion efficiency and thus the anabolic-catabolic coupling of the Wushen variety is significantly greater than the substrate carbon conversion efficiencies of the Yijinhuole and Helinger varieties.
Figure 3 shows that the temperature of maximum growth rate is similar among the three accessions, but the maximum growth rate differs. The maximum growth rate decreases in the order Yijinhuole>Wushen>Helinger, but note that total growth is proportional to the integral of the growth rate over the temperatures and time of the growth season. These curves show Wushen is adapted to grow at environmental temperatures from 12-43°C; Yijinhuole from 18-41°C and Helinger from 5-46°C. Yijinhuole may have a second, lower growth rate maximum around 10°C. Such bimodal curves of growth rate versus temperature have previously been found for plants adapted to climates with large, consistent diurnal temperature variations (McCarlie et al., 2003; Ward, 2007). In agreement with these results, Helinger is native to the coldest region, Yijinhuole is in the middle and Wushen is native to the warmest climate. Monthly average temperature and precipitation during the growth season are given in Fig. 4 and 5. Wushen is slightly warmer on average and the average temperatures are not significantly different between the other two sites, but averaging apparently hides significant differences in the temperature patterns. It is unfortunate that data on high and low temperatures at these sites are not available to compare with the temperature ranges derived from the growth curves
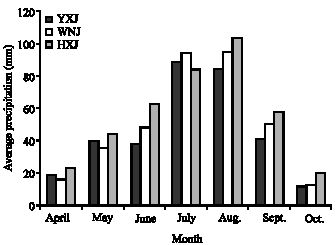 | |
| Fig. 5: | Monthly precipitation for Wushen (WNJ), Helinger (HXJ) and Yijinhuole (YXJ), Inner Mongolia |
.
DISCUSSION
As shown in Eq. 4, growth rate is equal to the respiratory CO2 production rate times an efficiency factor. Quantitative differences in efficiency and respiratory rate and differences in the temperature response of the respiration rate all contribute to the differences in growth characteristics of the three populations. Because the plants used in this study were grown from seed in a common environment, the differences in the growth curves are due to genetic adaptations and show the plants have limited ability to acclimate. The curves of growth rate versus temperature in Fig. 3 show that plants from the three sites are adapted to very different temperature patterns. That is, the maximum and minimum temperatures for growth are expected to match the extreme temperatures commonly encountered during the growth season and total seasonal growth is expected to be optimal when the curve of growth rate versus temperature matches the shape of the distribution curve for environmental temperatures at the native site (Criddle et al., 2005; Criddle and Hansen, 1999). The areas under the growth curves in Fig. 3 are approximately the same as expected for plants that grow to a similar size during the same length of growth season. The other principle governing the shape and position of curves of growth rate versus temperature for well adapted plants is that plants grow at the temperatures extant when water is available. The differences in the temperature patterns at the native sites as indicated by the growth curves in Fig. 3 are not captured in the average temperature and precipitation data in Fig. 4 and 5 because averaging data over time can hide large differences in temperature and precipitation patterns.
In conclusion, this study shows that no simple relation exists between any one respiratory characteristic and the growth properties of these three accessions of closely related plants. The temperature responses of the growth rates result in similar total seasonal growth of plants of each accession when grown in the native climate, but planting seeds from these seed sources outside their native ranges will probably not be successful because of the differences in temperature adaptation.
REFERENCES
- Criddle, R.S., L.D. Hansen, B.N. Smith, C. Macfarlane, J.N. Church, T. Thygerson, T. Jovanovic and T. Booth, 2005. Thermodynamic law for adaptation of plants to environmental temperatures. Pure Applied Chem., 77: 1425-1444.
Direct Link - Hansen, L.D., B.N. Smith and R.S. Criddle, 1998. Calorimetry of plant metabolism: A means to rapidly increase agricultural biomass production. Pure Applied Chem., 70: 687-694.
Direct Link - Hansen, L.D., C. Macfarlane, N. McKinnon, B.N. Smith and R.S. Criddle, 2004. Use of calorespirometric ratios, heat per CO2 and heat per O2, to quantify metabolic paths and energetics of growing cells. Thermochim. Acta, 422: 55-61.
Direct Link - Liu, Y.X., 1984. Some new species of Caragana and Chesneya from China. Acta Phytotaxon. Sin., 22: 210-210.
Direct Link - McCarlie, V.W., L.D. Hansen, B.N. Smith, S.B. Monsen and D.J. Ellingson, 2003. Anabolic rates measured by calorespirometry for eleven subpopulations of Bromus tectorum match temperature profiles of local microclimates. Russ. J. Plant Physiol., 50: 183-191.
Direct Link - Vitousek, P.M. and R.W. Howarth, 1991. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry, 13: 87-115.
CrossRefDirect Link - Yan, L., H. Li and Y. Liu, 2002. The anatomical ecology studies on the leaf of 13 species in Caragana genus. J. Arid Land Resour. Environ., 10: 100-105.
Direct Link - Zhang, M.L., Y.M. Huang, Y. Kang and Y.W. Wang, 2002. Floristics and vegetation of the genus Caragana in ordos plateau. Bull. Bot. Res., 22: 497-502.
Direct Link - Zhang, M.L. and G.H. Zhu, 2004. A new combination of Caragana (Fabaceae) from China. Bull. Bot. Res., 24: 5-6.
Direct Link - Zhao, Y.Z., 2005. The distribution pattern and ecological adaptation of Caragana microphylla, C. davazamcii and C. korshinskii. Acta Ecol. Sin., 25: 3411-3414.
Direct Link