
M.M. Tahat
Faculty of Agriculture, Universiti Putra Malaysia 43400 UPM,Serdang, Selangor Darul Ehsan, Malaysia
S. Kamaruzaman
Faculty of Agriculture, Universiti Putra Malaysia 43400 UPM,Serdang, Selangor Darul Ehsan, Malaysia
O. Radziah
Faculty of Agriculture, Universiti Putra Malaysia 43400 UPM,Serdang, Selangor Darul Ehsan, Malaysia
J. Kadir
Faculty of Agriculture, Universiti Putra Malaysia 43400 UPM,Serdang, Selangor Darul Ehsan, Malaysia
H.N. Masdek
Malaysian Agricultural Research and Development Institute, Malaysia
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 5 | Page No.: 479-484







ABSTRACT
The ability of endomycorrhizal fungi to colonize tomato (Lycopersicum esculentum. Mill) roots, was studied under glasshouse conditions. Two indigenous species; Glomus mosseae and Scutellospora sp. and non-indigenous species Gigaspora margarita were used in the study. Pot culture technique was used for re-culturing G. mosseae and Scutellospora sp., sorghum was used as a trap host. Gigaspora margarita was re-cultured by test tube technique. All species had the ability to colonize tomato root with different colonization levels. Significantly higher root were colonized by G. mosseae (80%) compared to G. margarita (20%). A G. mosseae significantly increased shoot dry weight (2.82 g) and flowers number (32.75 g) and root growth. Tomato plants treated by G. mosseae were higher significantly after seven week of plant growth. The colonization of tomato root by G. mosseae lead to bigger root size and more branching which increase positively the number of root tips, length, surface area and root volume. Higher spores (455/100 g) were counted in Glomus mosseae inoculated plant compared to Scutellospora sp. (250/100 g) and G. margarita plant (132/100 g).
PDF Abstract XML References Citation
How to cite this article
M.M. Tahat, S. Kamaruzaman, O. Radziah, J. Kadir and H.N. Masdek, 2008. Response of (Lycopersicum esculentum Mill.) to Different Arbuscular Mycorrhizal Fungi Species. Asian Journal of Plant Sciences, 7: 479-484.
DOI: 10.3923/ajps.2008.479.484
URL: https://scialert.net/abstract/?doi=ajps.2008.479.484
DOI: 10.3923/ajps.2008.479.484
URL: https://scialert.net/abstract/?doi=ajps.2008.479.484
INTRODUCTION
More than 80% of land plants are colonized by Arbuscular Mycorrhizal Fungi (AMF) (Ulrich et al., 2002). This reflects two aspects of AMF ecophysiology, firstly, AMF can adapt to many kinds of soil conditions, secondly, AMF have been shown to enhance plant growth mainly by increasing nutrient acquisition (Göher and Paszkowski, 2006).
AMF are obligate symbionts that require a plant host to complete their life cycle and produce spores (Smith and Read, 1997). Association has been found to be mutually beneficial, the fungus assists the plant in mineral nutrients uptake, while the plant supplies the fungus with carbon (from its fixed photosynthates). As a result of this association, colonization of the root system by AM fungi confers benefits to the host plant growth and development, through the acquisition of phosphate and other minerals from the soil. Colonization may also enhance the plants resistance to biotic and a biotic stresses (Smith and Read, 1997). The symbiosis effect directly to the host plant growth, effectiveness varies in individual plant-fungus combination (Smith et al., 2003), improving rhizospheric and soil conditions, producing plant growth hormones (Liderman, 1994), defending root against soil borne diseases (Azcón-Aguilar and Barea, 1997). Effect of Mycorrhiza on root morphology have attributed to improvement of P uptake by AMF, but AMF effect on hormone production may be responsible (Berta et al., 1993). Shoot dry mass of tomato inoculated with mycorrhiza was up to 243% greater than that of non-inoculated plants (Sylvia et al., 2001).
Around 17 species of mycorrhizal fungi were recorded from the rihzosphere of soil VAM tomato plants after 90 days of growth (Manian et al., 1995). The responses of plant to AMF influenced by many soil properties such as, soil pH (Clark, 1997), soil phosphorus level, plant species as well as inoculum levels (Schroeder and Janos, 2005). The lack of demonstrated benefit may be due to the use of inappropriate strains of fungi, relatively high available P in the soil, inappropriate inoculation of mycorrhizal fungi, inability of introduced AM fungi to establish in the soil and large variation in rates of plant grwoth because of heterogeneity of field soils (Jasper, 1994).
The main objectives of this research were to evaluate the potential of three tropical endomycorrhizal fungi species to colonize tomato plant root and spore production under glasshouse conditions using sterilized soil.
MATERIALS AND METHODS
Mycorrhizal spores production: This study was conducted during February-July 2007, by reculturing Glomus mosseae (Nicolson and Gerdman) and Scutellospora sp. (Walker and Sanders) using pot culturing technique (Mukerji, 2002). The inoculums of both species were collected using wet sieves technique (Gerdman and Niclson, 1968). Mature and healthy spores were selected under binocular microscope. Spores of Glomus mosseae and Scutellospora were obtained originally from the laboratory of soil microbiology, Land Management Department, Faculty of Agriculture, Universiti Putra Malaysia, Malaysia.
Gigaspora margarita (Gerdeman and Trappe) was obtained from Agricultural University Kampus IPB, Darmaga-Bogor-Indonesia. Test tube technique was used to reculture this species as described by Setiadi (2002). Glass test-tubes (2 cm diameter) were filled with 20 g of 1 mm mesh autoclaved sieved sand. Five spores/g sand were added to each test tube and one seed of sorghum (Sorghum vulgare L.) was seeded into the test tube above the spores. Two grams of sterilized sand were added to cover the seed and the spores. The system was watered every week with modefide Hogland solution nutrients (Vosatka and Gryndler, 1999).
Macronutrients (KNO3 = 240 mg, Ca (NO3) 2.4 H2O = 295 mg, MgSO4.7H2O = 720 mg, KH2PO4 = 12.2 mg, FeNaEDTA = 4.5 mg, NaCl = 0.7 mg) Micronutrients ZnSO4.7H2O = 0.75 mg, CuSO4.5H2O = 0.001 mg, MnCl2.4H2O = 0.75 mg, NaMoO4.2H2O = 0.00017 mg, H3BO3 = 1.5 mg and with distilled and sterilized water every three days. The tubes were placed in the growth chamber (28 C° with continuous white florescent light). The spore colonization was observed after three weeks of planting, the spores were collected after 45 days and recultured again in glasshouse for mass production of G. margarita inoculum using sorghum as trap host.
Soil preparation: Serdang series soil was used as a growth medium. The soil contained, 0.13% N, 0.023% P, 0.30% K, 0.063% Ca, 0.034% Mg, 0.063% S, 1.52% Fe, 0.0034% Mn, 0.0057% Zn, 0.00064% Mo, 0.0003% B, 0.0015% Cu and pH, 6.00,. Plastic pots (20 cm diameter) were filled with mixture of soil: sand (3:1 v:v). The soil was checked for microorganisms using Potato Dextrose Agar media (PDA) before used. The soil mixture was sterilized using autoclave at 121°C for one hour twice and it was allowed to cool for 24 hours before used.
Spore inoculation: Healthy, mature and clean spores of mycorrhizal fungi from pot sorghum culture were collected (50 spores/100 g dry soil) and mixed well with the top 5 cm of soil under the seedsbed (Al-Raddad, 1995).
| Table 1: | Effect of different mycorrhizal fungi treatments Glomus mosseae (GM), Scutellospora sp. (SS), Gigaspora margarita (GIM) and Control (C), on tomato plant height (3rd-7th week). |
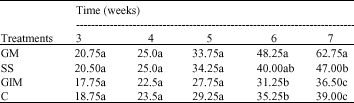 | |
| Column followed by the same letter(s) are not significantly different, according to Tukey Post Hoc test (p<0.05) |
Tomato plant: Commerical tomato seeds were rinsed with 0.1% NaOCl for five minutes and planted directly (three seeds/pot) in clean plastic pot. After 2 weeks seedling were thinned down to one seedling per pot. Plants were watered with tap water to field capacity. One hundred grams of peat were added to each pot to enhance seedling growth and spores germination. Tomato plants were grown under glasshouse conditions with day and night temperature 25-30°C. Four treatments were arranged randomly, Glomus mosseae (GM), Scutellospora sp. (SS), Gigaspora margarita (GIM), Control (C) (no inoculum).
Shoot growth: Plant height was measured five times during plant growth starting from the 3th week until the 7th week (Table 1). Flowers number were recorded in the last week. Harvest of plants was carried out under the cool conditions to minimized the water evaporation. Plants were dried in the oven (70°C for 24 h) for dry weight determination.
Root growth: Root tips, root length, root surface area and root volume were measured at the end of this experiment using the root analyzer machine.
Root dry weight: Plant root were collected and the fresh weight was measured. For root dry weight evaluation, the roots were kept in the oven (70°C for 24 h), then the dry weight was measured for each single plant.
Root colonization (%): The percentage of adventitious and lateral root colonized by AMF was evaluated microscopically followed by clearing of roots in 10% KOH and staining with 0.05% trypan blue in lactophenol according to method described by Phillips and Hayman (1970). The following formula was used to calculate the root colonization (Giovannetti and Mosse, 1980).
Mycorrhizosphere spores determination: Spores were separated from the soil by wet sieving and decanting technique. One hundred grams soil were collected and mixed with water. The mixture was pour through different sieve sizes (250, 106 and 45 μM). After several time of sieve washing the liquid was collected in petri dish and the spores were counted under binocular- microscope (Gerdman and Nicolson, 1968).
Statistical analysis: Treatments were arranged in Completely Randomized Design (CRD). Tukey test was used for means separation. Four replicates and one plant /replicate was used for each treatment. Statistical analysis was carried out with SPSS 15.0 software.
RESULTS
Shoot growth
Plant height: Growth was slow at the first weeks. Significant differences observed at week 6 week. Tomato growing with Glomus mosseae inoculum was significantly higher in comparison to other treatments at week 7 (Table 1) followed by plants inoculated with Scutellospora. There was no significant affect of G. margarita on plant growth.
Shoot dry weight: Shoot dry weight was highest in Glomus mosseae treatment. Compared to other species (Table 2). Glomus mosseae increased plant growth by 60% compared to Scutellospora and Giaspora margarita treatments.
Flower numbers: The flowers emergence started at the 7th week ( Table 2), the number of flowers was recorded at the end of this experiment. Plant inoculated with Glomus mosseae, produced the highest number of flowers relative to other species. Gigaspora margarita treatment recorded the lowest number of flowers compared to all other treatments (Table 2).
Root growth: Glomus mosseae inoculum was able to increase some root morphology, root dry weigh, root tips, root length significantly higher than Gigaspora margarita and Scutellospora. Root surface area was not significant affected by treatments. Root dry weight was highest in Glomus mosseae followed by Scutellospora (Table 3).
AMF root colonization: The colonization percentage of tomato roots was affected by the species Glomus mosseae used for colonization (Fig. 1). Giaspora margarita had the lowest colonization percentage (Table 4).
AMF spores: Similarly, highest number of spores was collected from the soil inoculated with Glomus mosseae treatment, followed by Scutellospora (Table 4).
| Table 2: | Effect of different mycorrhizal species on tomato dry shoot weight and flowers number |
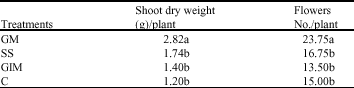 | |
Means in columns followed by the same letter(s) are not significantly different, according to Tukey Post Hoc test (p<0.05) |
| Table 3: | Effect of different mycorrhizal species on tomato root Dry Weight (DW), Root Tips (RT), Root Length (RL), Root Surface Area (RSA) and Root Volume (RV) |
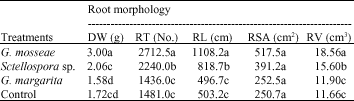 | |
Means in columns followed by the same letter(s) are not significantly different, according to Tukey Post Hoc test (p<0.05). Values are shown in means |
| Table 4: | Effect of different mycorrhizal fungi treatments Glomus mosseae Scutellospora sp. Gigaspora margarita and control on tomato root colonization percentage and soil spore number |
 | |
| Means followed by the same letter(s) are not significantly different, according to Tukey Post Hoc test (p<0.05) |
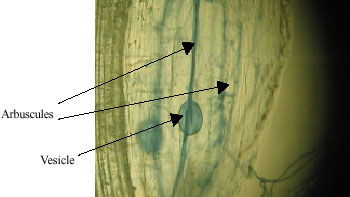 | |
| Fig. 1: | Root colonized by Glomus mosseae mycorrhizal fungi species |
DISCUSSION
The current investigation showed that AMF Glomus mosseae was able to significantly increase the shoot height and shoot dry of tomato. In the present study the higher shoot dry of Glomus mosseae maybe related to the higher photosynthesis activity. Also, present study indicated that the AM fungi could be provided a greater effective root surface area to explore greater volumes of soil and to overcome water and nutrient depletion zones around active root surfaces. Present results in the same line with the results obtained by many researchers (Davies et al., 1992; Clark, 1997), they reported that the main benefits of AMF are: enhance plant acquisition of mineral nutrients and increase the ability of host plants to withstand or reduced acquisition of toxic elements to growth.
Mycorrhizal fungi have increased significantly plant dry weight, height, fruit number and root colonization percentage more than non-mycorrhizal plant (Utkhede, 2006). Average diameter of fungal hyphae = 3-4 μM, which is smaller than root hairs diameter (>10 μM), therefore, fungal hyphae penetrate soil pores and contact with soil that roots hair would not able to contact. AM roots greatly enhance the acquisition of mineral nutrient in plant (Jakobsen, 1998). Mendeiros et al. (1994) reported that immobile minerals can increase shoot dry and fresh weight. AMF are known to supply P to the plant by scavenging available P through the large surface area of their hyphae, by their high-affinity P uptake mechanisms, by organic acid production and observed that different AMF species have different effects on the growth of host plants in term of dry mass.
We conducted that the number of flowers was corresponding with the inoculation by Glomus mosseae, the results obtained in this study concurred that reports by Aboul-Nassr (1996), on Zinnia and Tagetes plants, he reported an increased number of flowers after mycorrhization.
Endomycorrhizal fungi species were able to colonize tomato roots and increase plant growth significantly, this may be due to the structure of tomato root, spores density used and low nutrient in serdang serie soil (classified as a low available nutrient soil especially P). The results are similar to results obtained by other greenhouse. Present finding confirmed by the study of Perner et al. (2007), they reported that mycorrhizal colonization increased the number of buds and flowers, as well as shoot P and potassium (K) concentration. One of the possible mechanisms involved in P absorption is that, plants colonized by AMF is the changes in growth regulating components such as gibberelins, auxins and cytokinins (Linderman, 1992).
The high spore number of Glomus mosseae maybe either due to conducive edaphic conditions for sporulation like nutrient status and high aeration or undisturbed conditions of the soil, which allowed sufficient time for build-up of mycorrhizal spore (Azizah and Omar, 1991).
These results indicated that indigenous strains of Glomus mosseae may adapt to edaphic conditions within the time of reculturing. In our experiment, tomato seeds were planted directly in pots to accelerate spore germination by the production of exudates from seeds germination.
A possible mechanism by which AMF increased root size could be that AMF modify root function and change root architecture (Andrade et al., 1998). In this study the number of AM propagules significantly increased root morphology. Glomus intraradices inoculation induces a higher plant growth in autoclaved soil than in the non-disinfected soil. The present study confirmed that autoclaved soil was not stressed or disturbed by the soil microbial activity.
Further investigation of mycorrhizal adaptation (indigenous and non-indigenous) and the factors affecting the adaptation of AMF to local soils such as, soil pH, plant host, soil type, temperature, inocula amount, nutrients, interaction between soils and inoculums, could be useful in the application and establishment of new strains of mycorrhizal fungi in local agroecosystem.
CONCLUSION
Inoculation with arbuscular mycorrhizal fungi was significantly enhanced and increased tomato shoot growth and root morphology structure. Th endogenous species, Glomus mosseae was the best species among three species tested. The introduced species was an able to adapt with the Malaysian local conditions. Glomus mosseae may be an effective and environmentally sustainable organisms to develop and increase tomato production under glasshouse conditions.
ACKNOWLEDGMENTS
The authors are grateful to acknowledge the Universiti Putra Malaysia, for funding this study (grant No. IRPA 01-02-04-0394-EA001). We wish to thank Dr. Yadi Setyadi from Indonesia for providing us with Gigaspora margarita inoculum and Dr. Siddig Ibraheem for his assistance with data analysis.
REFERENCES
- Aboul-Naser, A., 1996. Effect of vesicular-arbuscular mycorrhiza on Tagetes erecta and Zinnia elegans. Mycorrhiza, 6: 61-64.
CrossRefDirect Link - Al-Raddad, A., 1995. Mass production of Glomus mosseae spores. Mycorrhiza, 5: 229-231.
CrossRefDirect Link - Andrade, G., K.L. Mihara and R.G. Liderman, 1998. Soil aggregation status and hizobacteria in the mycorrhizosphere. Plant Soil, 202: 89-96.
CrossRefDirect Link - Azcon-Aguilar, C. and J.M. Barea, 1997. Applying mycorrhiza biotechnology to horticulture: Significance and potentials. Sci. Hortic., 68: 1-24.
CrossRefDirect Link - Berta, G., A. Fusconi and A. Trotta, 1993. VA mycorrhizal infection and the morphology of root systems. Environ. Exp. Bot., 33: 159-173.
CrossRefDirect Link - Clark, R.B., 1997. Arbuscular mycorrhizal adaptation, spore germination, root colonization and host plant growth and mineral acquisition at low pH. Plant Soil, 192: 15-22.
CrossRefDirect Link - Davies, F.T., J.R. Potter and R.G. Linderman, 1992. Mycorrhiza and repeated drought exposure affect drought resistance and extraradical hyphae development of pepper plants independent of plant size and nutrient content. J. Plant Physiol., 139: 289-294.
Direct Link - Gerdemann, J.W. and T.H. Nicolson, 1963. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc., 46: 235-244.
CrossRefDirect Link - Giovannetti, M. and B. Mosse, 1980. An evaluation of techniques to measure vesicular-arbuscular infection in roots. New Phytol., 84: 489-500.
CrossRefDirect Link - Gorhe, V. and U. Paszkowski, 2006. Contribution of the arbuscular mycorrhizal symbiosis to heavy metal phytoremediation. Planta, 223: 1115-1122.
CrossRefDirect Link - Jakobsen, I., 1998. Transport of Phosphorus and Carbon in VA Mycorrhizas. In: Mycorrhiza: Structure, Function, Molecular Biology and Biotechnology. 2nd Edn. Varam, A. and B. Hock (Eds.). Springer-Verlay, ISBN: 3-540-63981-0 pp: 305-332.
Direct Link - Jasper, D.A., 1994. Management of Mycorhhizas in Revegetation. In: Management of Mycorrhizas in Agriculture, Horticulture and Forestry. 1st Edn. Robson, A.D., L.K. Abbot and N. Malajczuk (Eds.). Kluwer, Dordrecht, ISBN-13: 978-0792327004 pp: 211-219.
Direct Link - Harrier, L.A., 2001. The arbuscular mycorrhizal symbiosis: A molecular review of the fungal dimension. J. Exp. Bot., 52: 469-478.
CrossRefDirect Link - Liderman, R.G., 1992. Vesicular-Arbuscular Mycorrhizae and Soil Microbial Interactions. In: Mycorrhizae in Sustainable Agriculture. Betnlaenfalvay, G.J. and R.G. Linderman (Eds.). American Society of Agronomy, Madison, WI., ISBN: 0-89118-112-1, pp: 45-70.
Direct Link - Linderman, R.G., 1994. Role of VAM Fungi in Biocontrol. In: Mycorrhizae and Plant Health. Pfleger, F.L. and R.G. Linderman (Eds.). American Phytological Society, St. Paul. MN, APS Press, USA., ISBN-10: 0890541582.
Direct Link - Manian, S., T.T. Edathil and K. Udaiyan, 1995. Vesicular-arbuscular mycorrhizal colonization and growth of tomato (Lycopersico esculentum) in autoclaved soil. Pertanica. J. Trop. Agric. Sci., 18: 95-101.
Direct Link - Medeiros, C.A.B., R.B. Clark and J.R. Ellis, 1994. Growth and nutrient uptake of sorghum cultivated with vesicular-arbuscular mycorrhiza isolates at varying pH. Mycorrhiza, 4: 185-191.
CrossRefDirect Link - Mukerji, K.G., C. Manoharachary and B.P. Chamola, 2002. Techniques in Mycorrhizal Studies. 1st Edn., Kluwer Academic Publishers., London-Netherlands, ISBN-10: 1402005326, pp: 285-296.
Direct Link - Phillips, J.M. and D.S. Hayman, 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc., 55: 158-161.
CrossRefDirect Link - Simpson, D. and M. Daft, 1990. Spore poroduction and mycorrhizal development in various tropical crop Hosts Infected with Glomus fasciculatum. Plant Soil, 121: 171-178.
CrossRefDirect Link - Smith, S.E., F.A. Smith and I. Jokobsen, 2003. Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol., 133: 16-20.
PubMedDirect Link - Smith, S.E. and D.J. Read, 1997. Mycorrhizal Symbiosis. 2nd Edn., Academic Press, London, UK., ISBN-13: 978-0-12-652840-4, Pages: 605.
Direct Link - Sylvia, D.M., A.K. Alagely, D.O. Chellemi and L.W. Demchenko, 2001. Arbuscular mycorrhizal fungi influence tomato competition with bahiagrass. Biol. Fert. Soil, 34: 448-452.
CrossRefDirect Link - Ravnskov, S. and I. Jakobsen, 1995. Functional compatibility in arbuscular mycorrhizas measured as hyphal as P transport to the plant. New Phytol., 129: 611-618.
CrossRefDirect Link - Schroeder. M.S. and D.P. Janos, 2005. Plant growth, phosphorus nutrient and root morphological responses to arbuscular mycorrihzas, phosphorus fertilization and intraspecific density. Mycorrhiza, 15: 203-210.
CrossRefDirect Link - Perner, H., D. Schwarz, C. Bruns, P. Mader and G. Eckhard, 2007. Effect of arbuscular mycorrhizal colonization and two levels of compost supply on nutrient uptake and flowering of pelargonium plants. Mycorrhiza, 17: 469-474.
CrossRefDirect Link - Marschner, H. and B. Dell, 1994. Nutrient uptake in mycorrhizal symbiosis. Plant Soil, 159: 89-102.
CrossRefDirect Link - Hildebrandt, U., K. Janetta and H. Bothe, 2002. Towards growth of arbuscular mycorrhizal fungi independent of a plant host. Applied Environ. Microbiol., 68: 1919-1924.
CrossRefDirect Link - Utkhede, R., 2006. Increased growth and yield of hydroponcally grown green house tomato plants inoculated with arbuscular mycorrhizal fungi and Fusarium oxysporum f. sp. radicis-lycopersici. Biol. Control, 51: 393-400.
CrossRefDirect Link - Walker, T.S., H.P. Bais, E. Grotewold and J.M. Vivanco, 2003. Root exudation and rhizosphere biology. Plant Physiol., 132: 44-51.
CrossRefPubMedDirect Link - Al-Momany, A.A.R., 1999. Response of bean, broadbean and chickpea plants to inoculation with Glomus species. Sci. Hortic., 46: 195-200.
CrossRef - Vosatka, V. and M. Gryndler, 1999. Treatment with culture fractions from Pseudomonas putida modifies the development of Glomus fistulosum mycorrhiza and the response of potato and maize plants to inoculation. Applied Soil Ecol., 11: 245-251.
CrossRefDirect Link