
H.O. Edeoga
Department of Biological Sciences, Michael Okpara University of Agriculture, Umudike, P.M.B. 7267, Umuahia, Abia State, Nigeria
G. Omosun
Department of Biological Sciences, Michael Okpara University of Agriculture, Umudike, P.M.B. 7267, Umuahia, Abia State, Nigeria
G.G.E. Osuagwu
Department of Biological Sciences, Michael Okpara University of Agriculture, Umudike, P.M.B. 7267, Umuahia, Abia State, Nigeria
O.O. Emezue
Department of Biological Sciences, Michael Okpara University of Agriculture, Umudike, P.M.B. 7267, Umuahia, Abia State, Nigeria
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 4 | Page No.: 424-426







ABSTRACT
Leaf epidermal investigations were carried out on three species of Mimosa, M. pudica, M. invisa and M. pigra common in Nigeria. The results obtained revealed the nature of the irregularly shaped epidermal cell walls of M. pigra and M. pudica. The epidermal cell walls of M. invisa were however mostly rectangular to pentagonal in shape. Morphological type of stomata was the diacytic type in all the species. The average stomatal index of 13.43% on the upper epidermis of M. invisa and that of 40.91% on the lower epidermis was significantly different from the stomatal index of the other two species. The result of this investigation will help in the taxonomy and also contribute to more information on the biology of these three obnoxious species of Mimosa.
PDF Abstract XML References Citation
How to cite this article
H.O. Edeoga, G. Omosun, G.G.E. Osuagwu and O.O. Emezue, 2008. Micromorphology of the Leaf Cuticle in Mimosa Species (Leguminosae-Mimosoideae). Asian Journal of Plant Sciences, 7: 424-426.
DOI: 10.3923/ajps.2008.424.426
URL: https://scialert.net/abstract/?doi=ajps.2008.424.426
DOI: 10.3923/ajps.2008.424.426
URL: https://scialert.net/abstract/?doi=ajps.2008.424.426
INTRODUCTION
Fabaceae is regarded as the modern and accepted name of the family of which Mimosa belongs (Marshman, 2004). The subfamilies are Mimosoideae, Caesalpiniodeae and Papilionoideae (which is now regarded as Faboideae). Vidyarthi and Tripathi (2002) reported that the subfamily has 56 genera and 2000 species, which are commonly found in tropical and subtropical countries.
The Mimosa species are characterized by their nyctinastic movement. It is seen most especially and rapidly in M. pudica (Arbonnier, 2004). The nyctinastic movement (opening and closing) of the leaves is a circadian phenomenon and persists even when plants are kept continuously in light or darkness. M. pudica is a widespread weed of native America but was introduced to West Africa in 1839 and 1840 (Burkill, 1995).
Mimosa invisa Mart. commonly called giant sensitive plant is a biennial or perennial legume. It starts as an erect plant and grows into a spiny climbing shrub with the capacity to smoother competing plants in its domain. It has a variety known as M. invisa var Inermis Adelb. They are similar but can be distinguished by the absence of prickles on the stem, leaves and fruits of the variety Inermis. Burkill (1995) described M. invisa as one of the worst weeds due to its nasty thorns. According to Alabi et al. (2001), M. invisa limits the yield of cassava (Manihot sp.) in South Western Nigeria where more than 30% of total national production is generated.
M. pigra often found in riverbanks and fresh water swamps has been found common throughout the regions from Senegal to Nigeria and widespread over tropical Africa. It is also known as M. asperata Linn. (Burkill, 1995). M. pigra is an invading woody, prickly, leguminous shrub and it is capable of a dramatic population explosion (Miller and Lonsdale, 1993). M. pigra is similar in appearance to M. invisa. It can be distinguished by its very sensitive leaves, very strong thorns and pale mauve flower balls which differ from the pinkish flower of M. invisa.
This study is carried out with a view to using available differences in the micromorphology of the leaf cuticle in characterizing these three common and economically important Mimosa species available in Nigeria.
MATERIALS AND METHODS
This study was carried out between January and July, 2005 at Michael Okpara University of Agriculture, Umudike, Nigeria. The fresh leaves were collected from plants growing in natural conditions in different locations of Eastern Nigeria. The leaf samples were boiled in concentrated HNO3 for 2 min. The samples were then carefully washed in water and the adaxial and abaxial epidermis teased from the mesophyll using fine forceps and dissecting needles. The peelings of the leaves were then mounted in glycerine on glass slides.
Photomicrographs were taken using a Leitz Wetzler Ortholux microscope fitted with a Vivitar-V335 camera.
RESULTS
The characteristic epidermal features of the three taxa are shown in Fig. 1-2 and Table 1. The morphological type of stomata found in the three Mimosa species is the
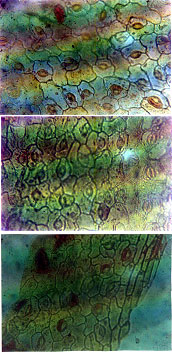 |
Fig 1: | (a) Epidermal morphology showing diacytic stomatal arrangement in lower epidermis of M. pudica. (b) Upper epidermal cells of M. pudica with irregularly shaped epidermal cell walls. (c) Lower epidermis of M. pigra with irregularly shaped epidermal cell walls |
| Table 1: | Epidermal characters of the Mimosa species studied |
 | |
diacytic stomata type. The epidermal cells were mostly irregularly shaped, but the epidermal cell walls of M. invisa were mostly rectangular to pentagonal in shape (Fig. 2a, b).
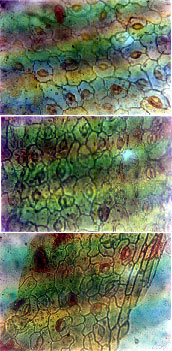 |
| Fig 2: | (a) Lower Epidermis of M. invisa with polygonal shaped cells. (b) M. invisa showing less number of stomata on upper epidermis. (c) Irregularly shaped epidermal cell walls of upper epidermis of M. pigra |
The anticlinal cell walls of the three taxa were straight in nature. Stomata was present on both the lower and upper epidermis of all the species investigated, i.e., they were amphistomatic. From data collected on the stomata index, stomata were more on the lower epidermis than on the upper epidermis. The stomatal index in the lower epidermis varied from 22.95% (M. pigra through 13.43 (M. invisa) to 24.39% in M. pudica.
DISCUSSIONS
Leaves are probably the most varied organs of the angiosperms. Carlquist (1961) has reported the valuable role of leaf anatomy in taxonomy. Irvine (1961), Metcalfe and Chalk (1983) have reported the use of epidermal characters such as leaf surface, epidermal cell wall pattern, nature of stomata as identifiable aids of some families and genera and sometimes for species. In the presently investigated Mimosa species, the epidermal cell walls varied from irregular in shape to rectangular or pentagonal in both the upper and lower epidermis.
The stomata type was diacytic in all species. Remarks have been made by some researchers in different plants on the use of epidermal characters in taxonomy (Edeoga and Ugbo, 1997; Edeoga and Ikem, 2002; Chandra et al., 1969). Stomatal frequency is one of the most widely used characters in taxonomy and pharmacognosy. (Karishnamurthy and Sundaram, 1970). Many other researchers like Ahmad (1964) have established the significance of stomatal frequency as a taxonomic tool. In the Mimosa species studied, the stomatal frequency varied significantly. The very low occurrence of stomata on the upper epidermis of M. invisa and the relatively high stomatal index of the lower epidermis of M. invisa also can be conveniently used as a distinguishing character from other Mimosa species studied.
The results obtained from the study of the micromorphology of the three Mimosa species will help in separating these taxa from other similar species or related taxa. This study, apart from the general morphology of these plants will help in the recognition of the Mimosa species of economic and medicinal importance. It will also contribute to the biology of the obnoxious species of Mimosa.
From the shape of the epidermal cell wall, M. invisa could be separated from the three taxa due to its rectangular-pentagonal shaped epidermis. This is in line with earlier affirmation by other researchers (Chandra et al., 1969). Similarly, M. pigra could be separated from M. pudica due its relatively lower value of stomatal index (22.95) as against the relatively high stomatal index of M. pudica (27.50). This is not strange as well since similar observations have been made by other researchers in other taxa (Edeoga and Jimoh, 2005; Chandra et al., 1969).
An interesting area of further research is that of their exhibition of nyctinastic movement by these taxa. Differences in leaf epidermal micromorphology could be involved in the various degrees of response to the sense of touch by these Mimosa species, investigated. However, further research is required in this direction so as to understand clearly the probable role of leaf characters in nyctinastism.
REFERENCES
- Ahmad, K.I., 1964. Cuticular studies with special reference to abnormal stomata cells in Cestrum. J. Ind. Bot. Soc., 43: 165-177.
Direct Link - Alabi, B.S., A.O. Ayeni, A.A. Agboola and B.A. Majek, 2001. Giant sensitiveplant interference in cassava. Weed Sci., 49: 171-176.
CrossRefDirect Link - Arbonnier, M., 2004. Trees, Shrubs and Lianas of West African Dry Zones. 1st Edn., CIRAD, Margraf Publishers GMBH MNHN, USA., ISBN: 2876145790, Pages: 574.
Direct Link - Chandra, V., S.L. Kapor, P.L. Sharma and D.L. Kapoor, 1969. Epidermal and venation studies in Apocyanaceae: 1. Bull. Bot Surv. India, 11: 286-289.
Direct Link - Edeoga, H.O. and H.N. Ugbo, 1997. Histochemical localization of calcium oxalate crystals in leaf epidermis of some Commelina L. (Commelinaceae) and its bearing on taxonomy. Acta Phytotax. Geobot., 48: 23-30.
Direct Link - Edeoga, H.O. and C.I. Ikem, 2002. Tannin, saponin and calcium oxalate crystals in some Nigerian species of Boerhavia L. (Nyctagniaceae). S. Afr. J. Bot., 68: 386-388.
Direct Link - Lonsdale, W.M. and I.L. Miller, 1993. Fire as a management tool for a tropical woody weed: Mimosa pigra in Northern Australia. J. Environ. Manage., 39: 77-87.
CrossRefDirect Link