
Ozgur Tatar
Department of Field Crops, Faculty of Agriculture, Ege University, Bornova, Izmir, Turkey
Mithat Nuri Gevrek
Department of Field Crops, Faculty of Agriculture, Ege University, Bornova, Izmir, Turkey
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 4 | Page No.: 409-412







ABSTRACT
The effect of water stress on Triticum aestivum L. was investigated before and during a period of stress at the seedling stage. For this purpose Turkish bread wheat cultivar Gönen 98 was used in this research. Changing of proline and lipid peroxidation content (malonyl dialdehyde, MDA) at five different harvesting times (7, 14, 21, 28 and 35 days after sowing) during early vegetative stage was determined in relation to dry matter production and relative water content of leaves. After onset of stress treatment (14 days after sowing) decline of dry matter production by water stress was observed. The decreasing of relative water content and increasing of proline concentration of stressed plants harvested 28 days after sowing was markedly more than at other harvesting times. Water stress caused an increase of MDA content in leaves of plants. It reached highest level 28 days after sowing. The data showed that proline accumulation increased after lipid peroxidation content became higher and relative water content of leaves became lower. Therefore proline appeared to be mainly involved in protection against oxidative stress than osmotic adjustment during initial steps of water stress.
PDF Abstract XML References Citation
How to cite this article
Ozgur Tatar and Mithat Nuri Gevrek, 2008. Influence of Water Stress on Proline Accumulation, Lipid Peroxidation and Water Content of Wheat. Asian Journal of Plant Sciences, 7: 409-412.
DOI: 10.3923/ajps.2008.409.412
URL: https://scialert.net/abstract/?doi=ajps.2008.409.412
DOI: 10.3923/ajps.2008.409.412
URL: https://scialert.net/abstract/?doi=ajps.2008.409.412
INTRODUCTION
The percentage of drought affected land areas more than doubled from the 1970s to the early 2000s in the world (Isendahl and Schmidt, 2006). Agricultural production was also affected by the limitation of water sources because of the serious drought problem in many countries. For the purpose of crop production, yield improvement and yield stability under water stress conditions, developing of drought tolerant varieties is the best option (Siddique et al., 2000). Therefore physiological and biochemical approaches have a great importance in order to understand the complex responses of plants to water deficiency and develop rapidly new varieties.
Drought inhibits plant growth due to its detrimental effects on main physiological and biochemical processes in plants. Water availability mostly affects growth of leaves and roots, stomatal conductance, photosynthesis and dry matter accumulation (Blum, 1996). One of the initial responses of plants to water stress is the decrease of leaf elongation rate and closing of stomata in order to reduce water consumption via transpiration. Accumulation of some organic compatible solutes such as sugars, betaines and proline which adjust the intercellular osmotic potential is also early reaction of plants to water stress. Stomatal limitation as a response to water deficiency attributes to decrease in both photosynthetic rate and internal CO2 concentration (Reddy et al., 2004). Decreasing of CO2 concentration in the tissues of drought stressed leaves and dissipation of excess light energy in the PSII core and antenna cause generating active oxygen species, which are potentially dangerous under water stress conditions (Peltzer et al., 2002). Sairam and Saxena (2000) reported that oxidative stress which caused metabolic damage in water stress, increases lipid peroxidation, resulting in greater membrane injury and pigment bleaching. Production of enzymatic and non-enzymatic antioxidants in plant cell plays an important role to avoid detrimental effects of free radicals.
Because of limited water in soil around the root, some compounds such as proline accumulate in cell to obtain suitable conditions for taking up the water. Proline, which is usually considered as an osmoprotection agent, is also known to be involved in reducing the oxidative damage by scavenging and/or reducing the free radicals. Zlatev and Stoyanov (2005) suggested that proline accumulation of plants could be only useful as a possible drought injury sensor instead of its role in stress tolerance mechanism. However, Vendruscolo et al. (2007) found that proline is involved in tolerance mechanisms against oxidative stress and this was the main strategy of plants to avoid detrimental effects of water stress. Although there is a consensus that most of the stress factors cause increasing of proline accumulation in plants, it is still not clear whether it is involved in tolerance mechanism or not.
The effect of water stress on proline accumulation, lipid peroxidation and water relations of wheat was investigated in this research.
MATERIALS AND METHODS
The seeds of Turkish bread wheat cultivar Gönen 98 were used as plant material. The experiments were conducted in the Laboratories of Ege University, Faculty of Agriculture, Department of Field Crops, Turkey in 2007. Eight seeds were sown in pots containing soil. Seven Days After Sown (DAS), uniform seedlings were selected and the pots were transferred to climate chamber. The temperature was maintained at 20°C during 12 h of light condition (under light intensity of 10000 lux) and 17°C during 12 h of dark conditions. Humidity was between 40-50%. After 14 days of growth with normal water supply, stress was applied by withholding water from half of the pots. Four plants were harvested five times during the experiment (7, 14, 21, 27 and 35 DAS). Water use of plants was calculated after evaporation was subtracted.
Relative Water Content (RWC) of leaves was estimated according to the method of Weatherly (1950).
Determination of the content of lipid peroxides: The level of lipid peroxidation was measured in terms of Malonyl Dialdehyde (MDA) content, a product of lipid peroxidation following method of Heath and Packer (1968). The plant material (0.3 g) was grinded with 3 mL of 0.1% Trichloroacetic Acid (TCA). The homogenate was centrifuged at 10000 g for 20 min. A 0.5 mL aliquot of the supernatant was mixed with 1.5 mL solution of 20% TCA containing 0.5% Thiobarbituric Acid (TBA). The mixture was incubated in a boiling water bath for 30 min then quickly cooled in an ice bath and then warming to room temperature. The extinction was measured at 532 nm and 600 nm. The MDA content was calculated and expressed as μmol MDA per gram fresh weight.
Determination of proline content: The proline content was measured by the method of Bates et al. (1973). The plant material (0.5 g) was grinded with 10 mL 3% sulfosalicylic acid. The homogenate was filtered. Acetic acid (2 mL) and acidic ninhydrin regent (2 mL) were added to a 2 mL aliquot. The mixture was thoroughly stirred and incubated in a boiling water bath for 1 h. Then it was transferred to ice bath and warmed to room temperature. Toluene (4 mL) was added to the mixture and the extinction of upper toluene level was measured at 518 nm.
RESULTS AND DISCUSSION
Dry matter production was significantly reduced by water stress (Fig. 1). While continual increase of dry matter production was observed in control plants (284 mg 35 DAS), the increase was slightly in stressed plants (165 mg 35 DAS). Closure of stomata and decrease in carbon dioxide concentration as an initial response to water stress inhibited dry matter production due to limitation of photosynthesis (Reddy et al., 2004). In the first days after onset of treatment a slight decrease in relative water content occurred but decreasing became significant 14 days after withholding water (Fig. 2). Kameli and Lösel (1996) found similar decrease in RWC in wheat approximately 10 days after stress treatment. However, they reported that it was a reversible, decrease, which
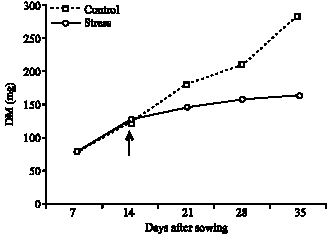 | |
| Fig. 1: | Changes in Dry Matter (DM) content of leaves in control and stressed plants of wheat before and during water stress treatment (Arrow indicate the start of stress treatment) |
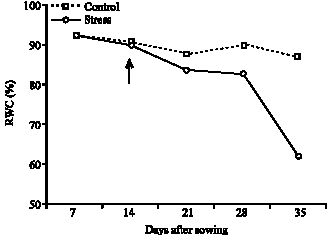 | |
| Fig. 2: | Changes in Relative Water Content (RWC) of leaves in control and stressed plants of wheat before and during water stress treatment (Arrow indicate the start of stress treatment) |
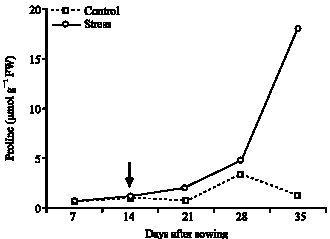 | |
| Fig. 3: | Effects of water stress on proline content of leaves in control and stressed plants of wheat before and during water stress treatment (Arrow indicate the start of stress treatment) |
recovered quickly after re-watering. Siddique et al. (2000) reported that there was a positive relation between RWC and photosynthetic rate. The results of the present study also showed that the dry matter as a photosynthetic product has also a correlation with RWC (Fig. 1, 2). Proline content didn`t show difference between stressed and control leaves but increased sharply 21 days after water was withheld (Fig. 3). While proline concentration was 1.32 μmol g-1 in well-watered conditions, it was 18.05 μmol g-1 in stressed plants at the last harvesting time of the experiment (21 days). Higher proline content in wheat plants after water stress application has been reported by Vendruscolo et al. (2007). Increasing amount of proline was also established in several stress conditions such as salinity (Poustini et al., 2007), cold (Charest and Phan, 1990) and UV (Tian and Lei, 2007) in wheat.
MDA content of leaves started to increase by the time when stress was applied. Fourteen days after treatment (28 DAS) it reached highest level (6.1 μmol g-1) while it was 2.9 μmol g-1 in control plants (Fig. 4). Noticeably, after that MDA content remained at the same level (Fig. 4). Higher lipid peroxidation and lower relative water content of leaves have also been reported in water stress conditions by Sairam and Saxena (2000). In general, it is known that proline has an important role as an osmolyte and a reservoir of carbon and nitrogen. However recent studies have shown that it has a protective role against free radical-induced damage (Matysik et al., 2002). The results of our study also showed that proline accumulation increased drastically after lipid peroxidation level reached highest value while remaining unchanged until the end of the stress treatment (Fig. 3, 4). Therefore it may be suggested that increasing levels of proline helps
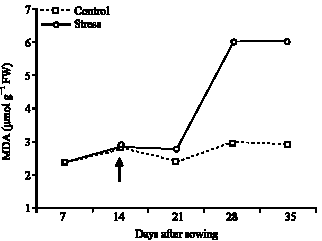 | |
| Fig. 4: | Effects of water stress on MDA content of leaves in control and stressed plants of wheat before and during water stress treatment (Arrow indicate the start of stress treatment) |
to protect membranes from oxidation instead of osmotic regulation as an initial response to water stress.
ACKNOWLEDGMENTS
We are grateful to visiting M.Sc. student Katka Markova from Chezch Republic and Ahmet Tümay for their helps in conducted the experiment. The seeds of Gönen were kindly supplied by Dr. Fatma Aykut Tonk, Ege University, Turkey.
REFERENCES
- Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Blum, A., 1996. Crop responses to drought and the interpretation of adaptation. J. Plant Growth Regul., 20: 135-148.
CrossRefDirect Link - Charest, C. and C.T. Phan, 1990. Cold acclimation of wheat (Triticum aestivum): Properties of enzymes involved in proline metabolism. Physiol. Plant., 80: 159-168.
CrossRefDirect Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Kameli, A. and D.M. Losel, 1996. Growth and sugar accumulation in durum wheat plants under water stress. New Phytol., 132: 57-62.
CrossRef - Matysik, J.A., B. Bhalu and P. Mohanty, 2002. Molecular mechanism of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci., 82: 525-532.
Direct Link - Peltzer, D., E. Dreyer and A. Polle, 2002. Temperature dependencies of antioxidative enzymes in two contrasting species. Plant Physiol. Biochem., 40: 141-150.
CrossRefDirect Link - Poustini, K., A. Siosemardeh and M. Ranjbar, 2007. Proline accumulation as a response to salt stress in 30 wheat (Triticum aestivum L.) cultivars differing in salt tolerance. Genet. Resour. Crop Evol., 54: 925-934.
CrossRefDirect Link - Reddy, A.R., K.V. Chaitanya and M. Vivekanandan, 2004. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol., 161: 1189-1202.
CrossRefPubMedDirect Link - Sairam, R.K. and D.C. Saxena, 2000. Oxidative stress and antioxidants in wheat genotypes: Possible mechanism of water stress tolerance. J. Agron. Crop Sci., 184: 55-61.
CrossRefDirect Link - Siddique, M.R.B., A. Hamid and M.S. Islam, 2000. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin., 41: 35-39.
Direct Link - Tian, X.R. and Y.B. Lei, 2007. Physiological responses of wheat seedlings to drought and UV-B radiation, effect of exogenous sodium nitroprusside application. Russian J. Plant Phys., 54: 676-682.
CrossRefDirect Link - Weatherley, P.E., 1950. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol., 49: 81-97.
CrossRefDirect Link - Zlatev and Z. Stoyanov, 2005. Effects of water stress on leaf water relations of young bean plants. J. Central Eur. Agric., 6: 5-14.
Direct Link