
S.R. Upadhyay
Nepal Agricultural Research Council, Kathmandu, Nepal
K.B. Koirala
Nepal Agricultural Research Council, Kathmandu, Nepal
D.C. Paudel
Nepal Agricultural Research Council, Kathmandu, Nepal
S.N. Sah
Nepal Agricultural Research Council, Kathmandu, Nepal
D. Sharma
Nepal Agricultural Research Council, Kathmandu, Nepal
D.B. Gurung
Nepal Agricultural Research Council, Kathmandu, Nepal
R.C. Prasad
Nepal Agricultural Research Council, Kathmandu, Nepal
R.B. Katuwal
Nepal Agricultural Research Council, Kathmandu, Nepal
B.B. Pokhrel
Nepal Agricultural Research Council, Kathmandu, Nepal
R.K. Mahato
Nepal Agricultural Research Council, Kathmandu, Nepal
R. Dhakal
Nepal Agricultural Research Council, Kathmandu, Nepal
N.B. Dhami
Nepal Agricultural Research Council, Kathmandu, Nepal
T.P. Tiwari
CIMMYT, South Asia Regional Office, P.O. Box 5186, Kathmandu, Nepal
G. Ortiz- Ferrara
CIMMYT, South Asia Regional Office, P.O. Box 5186, Kathmandu, Nepal
R.C. Sharma
CIMMYT, South Asia Regional Office, P.O. Box 5186, Kathmandu, Nepal
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 4 | Page No.: 375-381







ABSTRACT
This study was conducted in the hills of Nepal in four years to determine performance stability of open pollinated QPM genotypes in comparison to open pollinated cultivars of normal maize. Replicated field experiments were conducted in 29 environments using 20 QPM and seven normal maize genotypes. The normal maize genotypes included released cultivars, advanced breeding lines, one improved (Manakamana-3) and one local check (farmers` variety). Grain yield, days to flowering, plant and ear height, prolificacy, husk cover tightness and plant and ear aspect were analyzed. Stability and genotype superiority for grain yield was determined using genotype and genotypexenvironment (GGE) biplot analysis that compares among a set of genotypes with a reference ideal genotype, which will have the highest average value of all genotypes and be absolutely stable. The highest yielding QPM genotype in each year had significantly higher grain yield than the local check and higher or comparable yield to the improved check. Across years, many QPM genotypes produced significantly higher grain yield than the local check. Two genotypes (S03TLWQ-AB-01 and Obatampa) produced significantly higher grain yield than the improved check. GGE-biplot analysis showed that five of the seven most superior genotypes for grain yield were QPM (S03TLWQ-AB-01, Obatampa, S01SIYQ S99TLWQ-HG-AB and S99TLWQ-HG-A). Deuti and Manakamana-3 were the most superior among the normal maize cultivars. These genotypes also had acceptable to superior agronomic traits. Grain yield showed significant positive correlation with plant and ear height and prolificacy. The results show that superior open pollinated QPM genotypes were comparable to the outstanding cultivars of normal maize in performance stability and agronomic traits. The findings of this study provide new information on stability of the open pollinated QPM genotypes tested across warm rainfed hill environments. These cultivars are also adapted to other developing countries and this information could be useful for international and national QPM improvement programs.
PDF Abstract XML References Citation
How to cite this article
S.R. Upadhyay, K.B. Koirala, D.C. Paudel, S.N. Sah, D. Sharma, D.B. Gurung, R.C. Prasad, R.B. Katuwal, B.B. Pokhrel, R.K. Mahato, R. Dhakal, N.B. Dhami, T.P. Tiwari, G. Ortiz- Ferrara and R.C. Sharma, 2008. Performance of Quality Protein Maize Genotypes in the Warm Rainfed Hill Environments in Nepal
. Asian Journal of Plant Sciences, 7: 375-381.
DOI: 10.3923/ajps.2008.375.381
URL: https://scialert.net/abstract/?doi=ajps.2008.375.381
DOI: 10.3923/ajps.2008.375.381
URL: https://scialert.net/abstract/?doi=ajps.2008.375.381
INTRODUCTION
Maize (Zea mays L.) is an important food and a strategic crop in many parts of the developing world where livelihoods of millions of resource poor farmers depend on maize cultivation. Besides, several hundred million people in the developing world rely on maize as their principal daily food. In many countries in Latin America, Africa and Asia, maize is the staple food and at times, the only source of protein in human diet, which is deficient in amino acids lysine and tryptophan that are essential for human and monogastric animals (Bressani, 1992). Hence, nutritional quality of maize is a concern where it is consumed as human food. Quality Protein Maize (QPM) has superior nutritional and biological value (Prasanna et al., 2001). The importance of QPM in improving nutrition and livelihoods of the poor is well documented (Cordova, 2001). The superior nutritional value of QPM, compared to normal (non-QPM) maize and other cereals, has been reported by previous researchers (Prasanna et al., 2001; Vasal, 2001). The importance of QPM is also reflected through the release of QPM cultivars in many developing countries (Prasanna et al., 2001; Vasal, 2001; Krivanek et al., 2007).
Performance and stability of commercial QPM hybrids are well documented in the literature (Pixley and Bjarnason, 2002; Bhatnagar et al., 2003; Hossain et al., 2006). However, only limited information is available on the performance of open pollinated varieties (OPVs) of QPM (Pixley and Bjarnason, 2002; Akande and Lamidi, 2006). In early years of QPM development, agronomic traits of high yielding genotypes were a concern. However, in recent years many improved QPM cultivars have been developed by CIMMYT and partners in developing countries (Prasanna et al., 2001; Vasal, 2001; Akande and Lamidi, 2006; Badu-Apraku et al., 2006; Yasin et al., 2007), which are being promoted for commercial cultivation (CIMMYT, 1999; Krivanek et al., 2007). Studies have also shown that QPM hybrids are becoming more competitive with normal maize cultivars for performance, particularly in the tropical environments (Pixley and Bjarnason, 2002). However, information is needed on performance stability of OPVs of QPM for warm rainfed environments in particular. Further, information is also needed on comparative analysis of performance stability of OPVs of QPM and normal maize evaluated together. For their adoption, the OPVs of QPM cultivars must show grain yield and agronomic traits comparable to or better than the commercial OPVs of normal maize. This study was conducted to determine performance stability and suitability of agronomic traits of QPM genotypes in the hill environments in Nepal. One specific objective was to compare the performance of QPM with the commercial cultivars of normal maize.
MATERIALS AND METHODS
This study included 27 maize genotypes comprised of 20 OPVs of QPM and seven of normal type (Table 1). The normal type maize included advanced breeding lines and improved open pollinated cultivars. One improved (Manakamana-3) and one local check (farmers` variety) were also used in the study. Manakamana-3 is a high yielding medium maturing open-pollinated variety with tall height, one to two ears per plant, tolerance to lodging and turcicum leaf blight (Exserohilum turcicum). (CIMMYT, 2002).
Field trials were conducted in the 2004, 2005, 2006 and 2007 main maize seasons (April to September) in different mid-hill sites, spread from east to west Nepal. There were 29 different environments (year-site combinations) in the four years. Each trial was conducted in a randomized complete block with three replicates. Each experimental plot of 9 m2 was seeded at the standard seeding rate of 20 kg ha-1. Fertilizers were applied prior to seeding at the rate of 90, 30 and 30 kg ha-1 respectively of N, P2O5 and
| Table 1: | Mean values for various traits of 27 maize genotypes evaluated across 29 environments in the mid-hills of Nepal |
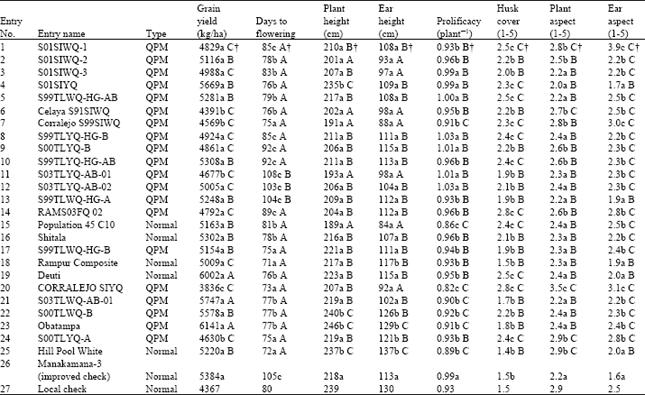 | |
| †a = Better than local check (LC), b = Equal to LC, c = Worse than LC, A = Better than improved check (IC), B = Equal to IC and C = Worse than IC for a given trait based on LSD0.05 | |
K2O in addition to 15 t farm yard manure ha-1. The plots were kept free of weeds by hand weeding. The trials were managed under summer and monsoon rainfed conditions. Other trial management practices were as per recommended maize crop husbandry in the country.
All traits in each plot were recorded according to the procedures described by CIMMYT for conducting standard maize trials. Days to flowering was recorded as the number of days from planting until the date on which 50% of the plants in a plot had 2-3 cm long silk. Plant and ear height were measured on the same five randomly selected plants in each plot between two and three weeks after flowering. Plant height was recorded as the distance from the plant base to the point where the tassel started to branch. Ear height was measured as the distance from plant base to the node bearing the uppermost ear. All plants and ears in each plot were counted. Prolificacy for each plot was determined as the number of ears divided by the number of plants times 100. Extremely small secondary ears were not recorded. Data on plant aspect (plant and ear height, uniformity of plants, disease and insect damage and lodging) in each plot was recorded at the brown husk stage on a scale of 1 to 5, where 1 and 5 represent excellent and poor, respectively. At maturity, husk cover was rated on a scale of 1 to 5, where 1 represents husk tightly covering the ear tip and extending beyond it and 5 signifies clearly exposed tips. After harvest, all ears from a plot were placed in a pile and ear aspect (size, disease and insect damage, grain-filling and uniformity) was recorded on a scale of 1 to 5, where 1 and 5 signify the best and the poorest, respectively. The plots were harvested individually, the cobs were threshed and grains weighed to record grain yield. Grain moisture content for each plot was recorded and grain yield was adjusted to 15% moisture basis.
Statistical analyses were conducted on various parameters recorded in the study. Since the values for plant and ear aspect and husk cover were between 1 and 5, data transformation was accomplished for these traits using (X + 0.5)½ as outlined by Gomez and Gomez (1984). The transformed data were used for analysis, but means have been reported after reverting the values to the original scale. The statistical analysis included an analysis of variance for each environment and a combined analysis across environments using SAS (SAS, 2003) software. After confirming the homogeneity of variance (Gomez and Gomez, 1984), a combined analysis of variance was also conducted. Each year-site combination was considered a unique and random environment, while genotypic effect was analyzed as fixed. The test of significance using F-ratios was conducted according to the procedure outlined by McIntosh (1983) for analysis of combined experiments. Since several experimental genotypes changed across years, environment was considered a bigger blocking factor, with replications of genotypes in each environment as replications nested within environments. Hence, analysis was conducted using Proc GLM. Significance of superiority of experimental genotypes to the checks was tested in pair wise comparisons performed using statistical LSD option in MEANS statement (Joshi et al., 2007).
To determine stability and identify superior lines across environments, genotype and genotype x environment (GGE) biplot analyses was conducted using GGE biplot software (Yan and Kang, 2002). GGE biplot is a method of graphical analysis of multi-environment data (Yan et al., 2000). The method differs from regular biplot analysis in that it simultaneously displays both genotypes and environments (Gabriel, 1971). The GGE biplot is a statistical tool that displays the main genotype effect (G) and the genotype x environment interaction of multi-environment tests. It is constructed by plotting the first two principal components (PC1 and PC2, also referred to as primary and secondary effects, respectively) derived from singular value decomposition of the environment-centered data. In this model, only the main effects of the genotypes plus G x E are absorbed into the bilinear terms. A specific option in GGE biplot analysis allows comparison among a set of genotypes with a reference genotype. This method defines the position of an ideal genotype, which will have the highest average value of all genotypes and be absolutely stable; that is, it expresses no genotype by environment interaction. A set of concentric circles are generated using the ideal genotype as the concentric center. The ideal genotype is used as a reference to rank the other genotypes. A performance line passing through the origin of the biplot is used to determine mean performance of a genotype. The arrow on the performance line represents increasing mean performance. A stability line perpendicular to the performance line also passes through the origin of the biplot; the two arrows in opposite directions represent decrease in stability. A genotype farther from the biplot origin on either side on the stability line represents relatively lower stability. A genotype closer to the performance line is considered more stable than the one placed farther. Average simple correlation coefficients (r) over environments were calculated using Fisher`s z-transformation (Sharma et al., 2006).
RESULTS
The analysis of variance revealed significant effect of environment on grain yield, days to flowering, plant and
| Table 2: | ANOVA for various traits for the 27 maize genotypes tested across 29 hill environments in Nepal |
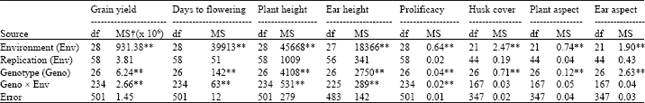 | |
**Significant at p = 0.01, xMS = Mean square | |
| Table 3: | Performance of the highest yielding QPM genotypes tested across 29 hill environments across four years in Nepal |
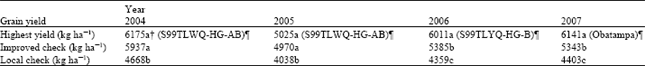 | |
| † Means within a column followed by different letter(s) are significantly different based on LSD0.05, ¶Highest yielding genotype in a given year | |
ear height, prolificacy, husk cover tightness and plant and ear aspect (Table 2). The 27 maize genotypes differed significantly for all traits. Genotype x environment interaction was significant for grain yield, days to flowering, plant and ear height and prolificacy.
In each of the four years, the highest yielder QPM genotype produced significantly higher grain yield than the local check (Table 3). The highest yielder QPM genotype produced significantly higher grain yield than the local and improved checks in 29 and 18 trials, respectively (data not shown). In 23 of the 29 trials, the highest yielder QPM genotype had grain yield not significantly different from the highest yielding normal maize cultivars. In 6 of the 29 trials, the highest yielder QPM genotype produced significantly lower (11 to 16%) grain yield than the highest yielder normal maize cultivar.
The 27 maize genotypes produced arrays of variation for grain yield (Table 1). The improved check (Manakamana-3) showed significantly higher grain yield, later flowering, shorter plant and ear height and superior plant and ear aspect than the local check. Fifteen QPM genotypes showed significantly higher grain yield than the local check. Two QPM genotypes [S03TLWQ-AB-1 (No. 21) and Obatampa (No. 23)] also produced significantly higher grain yield than the improved check, which was equal to the highest yielding normal maize (Deuti). Seven other QPM genotypes produced grain yield not significantly different from the improved check. The local check flowered significantly earlier than the improved check. Four QPM genotypes (7, 17, 20 and No. 24) flowered significantly earlier than the local check. Seventeen QPM genotypes flowered earlier than the improved check. Improved check had significantly shorter plant and ear height than the local check. There were four QPM genotypes (2, 6, 7 and No. 11) with significantly shorter plant height than both checks. Further, there were 13 other QPM genotypes with plant height not significantly different from the improved check. There were six QPM genotypes with significantly lower ear height than both checks. There were seven QPM genotypes with significantly higher prolificacy than the local check. None of the QPM genotypes showed significantly higher prolificacy than the improved check. However, there were 15 QPM genotypes with prolificacy comparable to the improved check. The two checks did not differ in husk cover tightness. None of the QPM genotypes showed tighter husk cover than the two checks. However, there were 11 QPM genotypes with husk cover tightness comparable to the two checks. There were 11 QPM genotypes with more superior plant aspect than the local check. None of the QPM genotypes showed plant aspect superior to the improved check. However, there were 16 QPM genotypes with plant aspect similar to the improved check. There were only two QPM genotypes with ear aspect better than the local check. None of the QPM genotypes showed better ear aspect than the improved check. However, there were two QPM genotypes with ear aspect equal to the improved check.
The GGE biplot analysis for grain yield revealed that three QPM (No. 4 = S01SYIQ, No. 21 = S03TLWQ-AB-1 and No. 23 = Obatampa) and one normal (No. 19 = Deuti) genotypes were close to the point of an ideal genotype (Fig. 1). Their performance and stability for grain yield were very close to that for an ideal genotype making them the highest yielding and most stable genotype across environments. Two other QPM (No. 13 = S99TLWQ-HG-A and No. 5 = S99TLWQ-HG-AB) and the improved check (No. 26 = Manakamana-3) also were relatively stable by being closer to the point of the ideal genotype.
Grain yield was significantly positively correlated with plant and ear height and prolificacy and significantly negatively correlated with husk cover tightness and plant and ear aspect (Table 4). There was a significant negative correlation of days to flowering with plant height and plant and ear aspect. Prolificacy showed a significant
| Table 4: | Average simple correlation coefficient of among various traits recorded on 27 maize genotypes tested across 29 warm rainfed environments in Nepal |
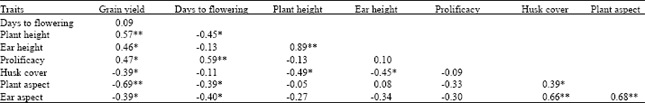 | |
*, **Correlation coefficient significantly different from zero at 0.05 and 0.01 probability levels, respectively. N = 27 | |
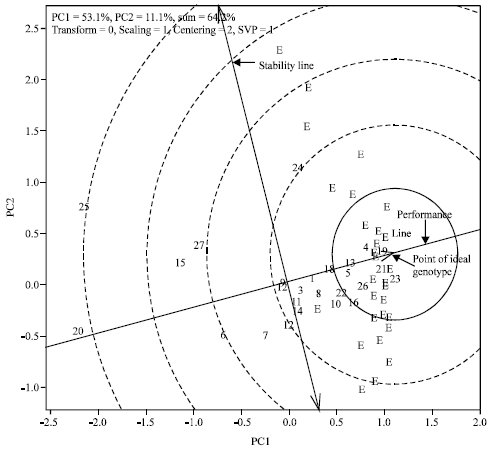 | |
| Fig. 1: | GGE biplot showing a comparison of 27 maize genotypes with an ideal genotype for grain yield tested on-station across 29 hill environments, Nepal. The environments are represented by the letter E and treated as random samples of the target environments (Refer to Table 1 for name of the genotypes) |
positive correlation with days to flowering. Husk cover tightness was significantly positively correlated with plant and ear aspect.
DISCUSSION
Mean grain yield of the maize genotypes differed across years and sites, which may be due to differing environmental conditions over time and sites. The sites themselves differed greatly in key attributes, such as geographic location, temperature and rainfall that affected performance (data not shown). The 27 genotypes represented a range of variability for grain yield and other agronomic characters (Table 1), with opportunities for selecting maize genotypes for high yield and acceptable agronomic characters. This was also reflected through a significant positive correlation of grain yield with days to flowering, plant and ear height and prolificacy.
This study identified several high yielding QPM genotypes such as S01SYIQ (No. 4), S99TLWQ-HG-AB (No. 5), S99TLYQ-HG-AB (No. 10), S99TLWQ-HG-A (No. 13), S03TLWQ-AB-1 ( No. 21), S03TLWQ-AB-01 (No. 22) and Obatampa (No. 23). The above QPM genotypes were also relatively stable (Fig. 1). Obatampa is a white dent and flint endosperm QPM with elevated levels of lysine and tryptophan and was released in several countries (Prasanna et al., 2001; Akande and Lamidi, 2006; Badu-Apraku et al., 2006). S99TLYQ-HG-AB (No. 10) was reported to be a high yielding QPM in Indonesia (Yasin et al., 2007).
There were three QPM (No. 4 = S01SYIQ, No. 21 = S03TLWQ-AB-1 and No. 23 = Obatampa) and one normal (Deuti)) genotypes that could be considered highly stable for grain yield across environments (Fig. 1). Besides, two other QPM (No. 5 = S99TLWQ-HG-AB and No. 13 = S99TLWQ-HG-A) genotypes and the improved check (No. 26 = Manakamana-3) could also be considered superior. All these genotypes could be of value for maize breeding programs in Nepal and in the region attempting to develop high yielding maize cultivars for warm rainfed environments.
In general, the high yielding QPM genotypes had satisfactory to superior maturity, plant and ear height and plant aspect compared to the local check. However, they were inferior to the checks in terms of husk cover and ear aspect. This suggests that husk cover tightness and ear characteristics of the high yielding QPM genotypes need to be improved for their adoption. Tight husk cover protects the ears on standing plants from rain water during monsoon which is common in Nepal. It also helps in storing the cobs during off season. Tight husk cover has also been reported important in other countries for resistance to earworm (Helicoverpa zea) (Archer et al., 1994; Butron et al., 2002) and Fusarium ear rot (Fusarium moniliforme) (Farrar and Davis, 1991).
The highest yielding QPM genotype showed significantly higher grain yield than the local check in four years. The highest yielder QPM genotype always produced higher grain yield the local check in all 29 environments. These findings demonstrate consistent genetic superiority of the QPM genotypes. This also suggests that, if properly chosen, farmers would never lose money for growing an appropriate QPM in place of the local check of the normal maize. On the contrary, the farmers growing a local cultivar of normal maize would frequently earn additional income by choosing an appropriate QPM. Several QPM genotypes also compared well with the best commercial cultivars and improved check of normal maize suggesting that QPM bears potential for the warm rainfed environments in the hills of Nepal. The finding that QPM cultivars are competitive and at times could be more productive than normal maize is in agreement with a previous report (Yasin et al., 2007). Pixley and Bjarnason (2002) reported that QPM OPVs were more stable than hybrids for grain yield with the latter producing 13% higher grain yield. Akande and Lamidi (2006) also reported that QPM hybrids outyielded OPVs and suggested that the later should be tested across diverse target environments before making their recommendation for commercial cultivation.
The adoption of QPM in Nepal and other developing countries would depend not only on its nutritional value, but also on its yield performance. The results of this study highlights the opportunities of this type of maize in increasing food sustainability, livelihoods and the nutritional aspects of millions of resource-poor farmers in the hills of Nepal. It is expected that these results would also be of value in other national, regional and/or international breeding programs aiming at improving and promoting QPM.
CONCLUSION
Exotic QPM genotypes tested under the diverse hill environments in Nepal showed significant variation for grain yield, days to flowering, plant and ear height, prolificacy, husk cover tightness and plant and ear aspect. There were QPM genotypes that outyielded the checks, were also highly stable, indicating that superior QPM germplasm is becoming available in the region through the international collaborative work.
Three QPM genotypes, Obatampa (No. 23), S03TLWQ-AB-01 (No. 21) S01SIYQ (No. 4), were comparable to the improved check (Manakamana-3) and the highest yielder (Deuti) normal maize cultivars in terms of grain yield and its stability, days to flowering, plant height (No. 21 only), ear height (4 and No. 21 only), prolificacy (No. 4 only), husk cover tightness (21 and No. 23), plant aspect and ear aspect (No. 4 only). With improvement in ear characteristics and husk cover tightness; these genotypes could be well adopted in the region. These high yielding QPM genotypes could also serve as improved parents for QPM improvement. Superiority of these exotic QPM genotypes underscores continuous development and dissemination of superior maize germplasm across continents in terms of saving resources.
A regular stability analysis often does not provide relative ranking of superior entries in reference to an ideal genotype that results in making a subjective judgment in selecting a cultivar. The GGE biplot approach used in this study could help breeders to better decide what genotypes should be promoted or released: the visual combined assessment of performance and its stability is a big advantage and adds confidence in the decision to promote a superior genotype.
ACKNOWLEDGMENTS
The authors appreciate the assistance of Mr. Surath Pradhan in preparing the tables and figures. This study was conducted as a part of the Hill Maize Research Project under financial support by Swiss Agency for Development and Cooperation (SDC) to the Government of Nepal and CIMMYT (Grant No. M0142).
REFERENCES
- Akande, S.R. and G.O. Lamidi, 2006. Performance of quality protein maize varieties and disease reaction in the derived-savanna agro-ecology of South-West Nigeria. Afr. J. Biotechnol., 5: 1744-1748.
Direct Link - Archer, T.L., B.R. Wiseman and A.J. Bockholt, 1994. Factors affecting corn earworm (Lepidoptera: Noctuidae) resistance in food corn. J. Agric. Entonol., 11: 9-16.
Direct Link - Badu-Apraku, B., S. Twumasi-Afriyie, P.Y.K. Sallah, W. Haag, E. Asiedu, K.A. Marfo, S. Dapaah and B.D. Dzah, 2006. Registration of obatanpa GH maize. Crop Sci., 46: 1393-1396.
Direct Link - Bhatnagar, S., F.J. Betrán and D.K. Transue, 2003. Agronomic performance, aflatoxin accumulation and protein quality of subtropical and tropical QPM hybrids in Southern USA. Maydica, 48: 113-124.
Direct Link - Bressani, R., 1992. Nutritional Value of High-Lysine Maize in Humans. In: Quality Protein Maize, Mertz, E.T. (Ed.). American Association of Cereal Chemists, St. Paul, MN., USA., ISBN: 0913250759, pp: 205-224.
Direct Link - Butron, A., N.W. Widstrom, M.E. Snook and B.R. Wiseman, 2002. Recurrent selection for corn earworm (Lepidoptera: Noctuidae) resistance in three closely related corn southern synthetics. J. Econ. Entomol., 95: 458-462.
CrossRefPubMedDirect Link - Farrar, J.J. and R.M. Davis, 1991. Relationships among ear morphology, western flower thrips and Fusarium ear rot of corn. Phytopathology, 81: 661-666.
Direct Link - Gabriel, K.R., 1971. The biplot graphic display of matrices with application to principal component analysis. Biometrika, 58: 453-467.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Hussain, M., S.R. Chughtai, H.I. Javed, H.N. Malik and M.H. Munawwar, 2006. Performance of locally constituted quality protein maize hybrids-a fortune for malnourished people and feed industry in Pakistan. Asian J. Plant Sci., 5: 385-389.
CrossRefDirect Link - Joshi, A.K., G. Ortiz-Ferrara, J. Crossa, G. Singh, R.C. Sharma, R. Chand and R. Prasad, 2007. Combining superior agronomic performance and terminal heat tolerance with resistance to spot blotch (Bipolaris sorokiniana) of wheat in the warm humid gangetic plains of South Asia. Field Crops Res., 103: 53-61.
CrossRefDirect Link - Krivanek, A.F., H. De Groote, N.S. Gunaratna, A.O. Diallo and D. Friesen, 2007. Breeding and disseminating Quality Protein Maize (QPM) for Africa. Afr. J. Biotechnol., 6: 312-324.
Direct Link - Pixley, K.V. and M.S. Bjarnason, 2002. Stability of grain yield, endosperm modification and Protein quality of hybrid and open-pollinated Quality Protein Maize (QPM) cultivars. Crop Sci., 42: 1882-1890.
Direct Link - Prasanna, B.M., S.K. Vasal, B. Kassahun and N.N. Singh, 2001. Quality protein maize. Curr. Sci. Assoc., 81: 1308-1319.
Direct Link - Sharma, R.C., B. Pandey-Chhetri and E. Duveiller, 2006. Heritability estimates of spot blotch resistance and its association with other traits in spring wheat crosses. Euphytica, 147: 317-327.
CrossRefDirect Link - Yan, W., L.A. Hunt, Q. Sheng and Z. Szlavnics, 2000. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci., 40: 597-605.
CrossRefDirect Link