
M. Matinizadeh
Department of Forest Research, Research Institute of Forests and Rangelands, P.O. Box 13185-116, Tehran, Iran
S.A.A. Korori
Department of Forest Research, Research Institute of Forests and Rangelands, P.O. Box 13185-116, Tehran, Iran
M. Teimouri
Department of Forest Research, Research Institute of Forests and Rangelands, P.O. Box 13185-116, Tehran, Iran
W. Praznik
Department of Chemistry, BOKU-University of Natural Resources and Applied Life Sciences,Muthgasse 18 A-1190 Vienna, Austria
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 4 | Page No.: 368-374







ABSTRACT
The seasonal dynamics of acid phosphatase, alkaline phosphatase and dehydrogenase and the relationship of their activities to soil depth were studied in the undisturbed forest soils under oak and compared with the disturbed one. At each site, four soil samples were randomly taken at depth of 0-20 cm in May, July, September and November and at three different depths (0-10, 10-20 and 20-30 cm) in November. Enzymes were assessed by reaction with substrate and photometrical method. The activity of acid and alkaline phosphatase and dehydrogenase was greater at undisturbed site than disturbed sites. These results signalized a high activity of roots for the secretion of acid phosphatases. High alkaline phosphatase activity reflected a good status of soil microorganisms. Moreover alkaline phosphatase activity was 2.1 to 2.93 times greater than acid phosphatase activity at both sites. Alkaline and acid phosphatase and dehydrogenase activity showed a seasonal pattern with maxima in summer and minimum in autumn. Assay of acid phosphatase activity showed significant (p<0.01) increasing of its activity with soil depth increment at undisturbed site whereas this increasing trend was not observed at disturbed site. The depth pattern of alkaline phosphatase activity was totally different. At undisturbed site, alkaline phosphatase activity significantly (p<0.01) decreased with soil depth and visa versa in the disturbed site. At both sites, dehydrogenase activity significantly (p<0.01) decreased with soil depth increment. In conclusion the higher activity of phosphatase and dehydrogenase at undisturbed sites shows the effect of land use management and for understanding soil ecosystem functioning.
PDF Abstract XML References Citation
How to cite this article
M. Matinizadeh, S.A.A. Korori, M. Teimouri and W. Praznik, 2008. Enzyme Activities in Undisturbed and Disturbed Forest Soils Under Oak (Quercus brantii var. persica) as Affected by Soil Depth
and Seasonal Variation. Asian Journal of Plant Sciences, 7: 368-374.
DOI: 10.3923/ajps.2008.368.374
URL: https://scialert.net/abstract/?doi=ajps.2008.368.374
DOI: 10.3923/ajps.2008.368.374
URL: https://scialert.net/abstract/?doi=ajps.2008.368.374
INTRODUCTION
In the west of Iran in Zagrosian forests (with an area of 5 million hectares), oak manna trees (Quercus brantii var. persica) are the dominant species in 1000-2300 m above sea level. This forest has the most extensive habitat among oak species in Iran. It is usually found in calcareous soils with alkaline pH. This species has high resistance against drought and low and high temperatures. In the last decades, this natural ecosystem has been disturbed due to intensive harvested, industrial development and agricultural activities. Recently programs have been developed for the protection of natural ecosystems and the rehabilitation of damaged regions in local or global levels. Having fundamental information about different aspects of forest ecosystems such as its soil quality and functioning is an important criteria in the success of management programs for ecosystem preservation. Visser and Parkinson (1992) have suggested that the biological and biochemical properties that are most useful for detecting the deterioration of soil quality are those that are most closely related to nutrient cycles, including soil respiration, microbial biomass, nitrogen mineralization capacity and the activities of soil enzymes. In particular, enzyme activities are especially significant in soil quality assessments because of their major contribution to the soil ability to degrade organic matter (Taylor et al., 2002; Schloter et al., 2003). The activities of enzymes (i.e., hydrolases) are important soil quality indicators because they are intimately involved in the cycling of nutrients, affect fertilizer use efficiency and reflect in part the microbiological activity in soil (Jordan et al., 1995; Dick et al., 2000; Taylor et al., 2002).
The phosphatases are a group of enzymes key to soil forest P cycling. Phosphatase activities can be a good indicator of the organic phosphorus mineralization potential and biological activity of soils (Speir and Ross, 1978; Dick and Tabatabai, 1993). Soil phosphatases, extracellularly secreted by plants and microorganisms, play a key role in the phosphorus cycle, allowing the formation of inorganic phosphorus, the only phosphate-form taken up by plants and microorganisms (Antonietta Rao et al., 2000). Phosphatase activity is related to soil and vegetation conditions (Herbien and Neal, 1990) and changes in response to management practices (Adams, 1992; Clarholm, 1993), soil temperature and moisture (Speir and Cowling, 1991). Phosphatases in soils are derived mostly from the microbial population (Tabatabai, 1994; Li et al., 1997) and have been suggested as a satisfactory index of microbial activity (Tabatabai, 1994; Tarafdar et al., 2001). In the other hand, acid phosphatase is mainly produced by plants but also soil microorganisms release acid phosphatase (Tabatabai, 1994). Alkaline phosphatase is produced by soil microorganisms and soil fauna (Findenegg and Neiemans, 1993; Tarafdar, 1995) whereas higher plants are devoid of alkaline phosphatase (Dinkelaker and Marschner, 1992; Tarafdar et al., 2001).
Dehydrogenase activity is used to estimate of overall microbial activity due to its presence in all microorganisms (Garcia et al., 1997; Taylor et al., 2002). It is often used as an indicator of microbial activity (Nannipieri et al., 1990; Dick, 1994). The dehydrogenase activity of a soil is the result of the activity of different dehyrogenases, which are an important component of the enzyme system of all microorganisms (enzymes of respiratory metabolism, citrate cycle and nitrogen metabolism). Dehydrogenase activity is thus an indicator of biological redox-systems and can be taken as a measure for the intensity of microbial metabolism in soil (Schloter et al., 2003).
Due to the importance of understanding soil quality in forest ecosystems related to metabolic capacities and biogeochemical cycling, the primary objectives of this study were to investigate selected soil chemical properties of an undisturbed region (UD) and disturbed region (DI) of the Zagrosian forest in Iran and to investigate the activities of dehydrogenase and two phosphomonoesterases (alkaline and acid) activities at the UD and DI forest regions as affected by seasonal variation, soil depth and soil properties.
MATERIALS AND METHODS
Site description: The study was carried out in two different sites in high forest with Quercus brantii var.
persica in Monj, Chahar Mahal va Bakhtiari province, Iran at a distance of 696 km from Tehran. Two sampling locations were chosen in Monj: UD in an almost undisturbed site (co-ordinates, 313013N, 503525E and elevation 2056 m) and DI in a disturbed site (co-ordinates, 313015N, 503628E and elevation 1955 m). The climate in Zagrosian forests is semi arid with unpredictably extreme change of temperatures in both summer and winter. In fact disturbed site is a degraded-unmanaged forest that trees are being cut from fifty years ago. The harvesting is done by traditional methods to obtain wood for domestic purposes. Rainfall and snowfall begin from mid-autumn and continue until spring March/April and the climate is dry from May until October. Long-term average precipitation is 520 mm and long-term average of temperature is 17.7°C. Mean maximum and minimum temperatures are 23 and 5.9°C, respectively. During the study period, annual temperature was 1.1°C below the long-term average and annual precipitation was 61 mm above the long-term average. Soil texture was sand-clay-loamy with pH 7.5-7.7 (Table 1). The DI site had less mature seeding crops and more gaps than UD site because of harvesting.
Soil sampling: At each site, four soil samples were randomly taken at depth of 0-20 cm in May, July, September and November 2003 and at three different depths (0-10, 10-20 and 20-30 cm) in November 2003. Soil samples were placed in tightly sealed plastic bags and transferred immediately to laboratory at 4°C. The soil samples were passed through a 2 mm sieve and divided into two parts: one fraction for the determination of physical and chemical factors, which was stored at room temperature and the other fraction for measuring of soil enzymes activities which was stored at 4°C. The soil were analyzed for pH in a 1:2.5 (soil: water) using a glass electrode, organic matter content by wet oxidation (Walkley and Black, 1934), total nitrogen by micro-Kjeldahl digestion procedure (Bremmer and Mulvaney, 1982) and soil texture and particle size analysis by the pipette method (Kilmer and Alexander, 1949). Olsen`s bicarbonate extractable P (PO43–) was also measured (Olsen et al., 1954). Soil microbial biomass was determined by method as described by Joergensen and Brookes (1990).
| Table 1: | Some characteristics of soil (0-20 cm) in every site |
 | |
| UD: Undisturbed region; DI: Disturbed region | |
Phosphatase assay: The activities of both phosphatases (Acid phosphatase and alkaline phosphatase) were determined in the field soil samples using the method described by Ohlinger (1996a).
After the addition of a buffered ρ-nitrophenyl phosphate solution, soil samples are incubated for 1 h at 37°C. The nitrophenol released by phosphomonoesterase activity is extracted and colored with sodium hydroxide and determined photometrically at 400 nm.
Dehydrogenase assay: Dehydrogenase activity was determined in the soil samples according to Ohlinger (1996b). In brief, field moist soils were suspended in a triphenyl tetrazolium chloride solution and incubated for 16 h at 25°C. The triphenyl formazan (TPF) produced was extracted with acetone and measured photometrically at 546 nm.
Statistical analyses: The results were analyzed with the SAS program and the means presented in this study for all measurements represents means (n = 4). All data in the study were subjected by analysis of variance and means were separated by Duncan`s multiple range tests.
RESULTS
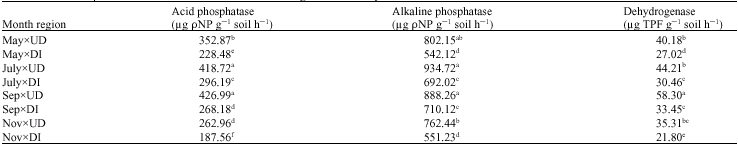
Phosphatase activities as affected by seasonal variation: Alkaline and acid phosphatase activities were significantly different (p<0.01) in both undisturbed and disturbed sites throughout all sampling times (Table 3). In this study, average alkaline phosphatase activity was 2.1-2.9 times higher than average acid phosphatase activity. Alkaline phosphatase activity ranged from 542.1 at disturbed site to 934.7 μg ρ-nitrophenol g-1 soil h-1 at undisturbed site (Table 4). Acid phosphatase activity ranged from 187.6 μg ρ-nitrophenol g-1 soil h-1 at disturbed site to 427.0 μg ρ-nitrophenol g-1 soil h-1 at undisturbed site (Table 4). Both enzyme activities followed a similar seasonal pattern at undisturbed and disturbed sites. The enzyme activities increased a few from fall to spring, continued increasing through spring and early summer, reached to the maximum activity in summer and then dropped again in fall.
Seasonal changes of alkaline phosphatase activity were not significantly different in undisturbed site (Table 2, 4). The highest activity was 934.7 μg ρ-nitrophenol g-1 soil h-1 in July and the lowest one was 762.7 μg ρ-nitrophenol g-1 soil h-1 in November in undisturbed site. In disturbed site the highest activity was 710.1 μg ρ-nitrophenol g-1 h-1 in September and the lowest was 542.1 μg ρ-nitrophenol g-1 soil h-1 in May. In this site, alkaline phosphatase activity in November was significantly lower than other sampling times. Acid phosphatase activity had significant differences at both sites in November in comparison to summer (Table 2, 4). The highest activity was 427.0 and 418.7 μg ρ-nitrophenol g-1 soil h-1 in September and July and the lowest one was 263.0 in November. In disturbed site the highest activity
| Table 2: | Mean comparison of season effect on soil enzymes activities |
 | |
| ρNP: ρ-nitrophenol; TPF: Triphenyl formazan. Alphanumeric indicates significance differences between treatments | |
| Table 3: | Mean comparison of region effect on soil enzymes activities |
 | |
| ρNP: ρ-nitrophenol; TPF: Triphenyl formazan; UD: Undisturbed region; DI: Disturbed region. Alphanumeric indicates significance differences between treatments | |
| Table 4: | Mean comparison of interaction between season and region on soil enzymes activities |
 | |
| ρNP: ρ-nitrophenol; TPF: Triphenyl formazan; UD: Undisturbed region; DI: Disturbed region. Alphanumeric indicates significance differences between treatments | |
| Table 5: | Mean comparison of depth effect on soil enzymes activities |
 | |
| ρNP: ρ-nitrophenol; TPF: Triphenyl formazan. Alphanumeric indicates significance differences between treatments | |
| Table 6: | Mean comparison of region effect on enzymes activities in different depths of soil |
 | |
| ρNP: ρ-nitrophenol; TPF: Triphenyl formazan; UD: Undisturbed region; DI: Disturbed region. Alphanumeric indicates significance differences between treatments | |
| Table 7: | Mean comparison of interaction between season and region on soil enzymes activities |
 | |
| ρNP: ρ-nitrophenol; TPF: Triphenyl formazan; UD: Undisturbed region; DI: Disturbed region. A1, A2 and A3: Soil depths of 0-10, 10-20 and 20-30 cm, respectively. Alphanumeric indicates significance differences between treatments | |
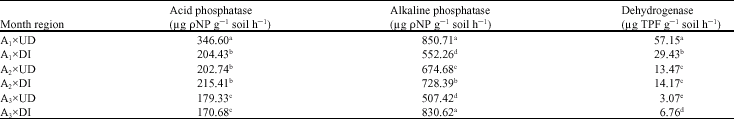
Phosphatase activities as affected by soil depth: At undisturbed site, acid phosphatase activity was greater at 0-10 cm depth 346.6 μg ρ-nitrophenol g-1 soil h-1 compared to 202.7 μg ρ-nitrophenol g-1 soil h-1 at 10-20 and 179.3 μg ρ-nitrophenol g-1 soil h-1 at 20-30 (Table 5, 7). At disturbed site, acid phosphatase activity did not vary with soil depths. Acid phosphatase activity was significantly different between undisturbed and disturbed sites at 0-10 and at 20-30 cm.
Alkaline phosphatase activity remarkably decreased with depth in soil profile from 850.7 to 507.4 μg ρ-nitrophenol g-1 soil h-1 at 0-10 and 20-30 cm depths. But at the disturbed site, this enzyme activity increased with depth in soil profile from 552.3 to 830.6 μg ρ-nitrophenol g-1 soil h-1 at 0-10 and 20-30 cm depths, respectively. In 10-20 cm depth, alkaline phosphatase activity was not significantly different between undisturbed and disturbed sites.
Dehydrogenase activities as affected by seasonal variation: Dehydrogenase activity were significantly different (p<0.01) in both undisturbed and disturbed sites throughout all sampling times (Table 6). Dehydrogenase activity ranged from 21.8 at disturbed site to 58.3 μg triphenyl formazan g-1 soil h-1 at undisturbed site. Same as acid and alkaline phosphatase, dehydrogenase followed a similar seasonal pattern at undisturbed and disturbed sites. Activities increased a few from fall to spring, continued increasing through spring and early summer, reached to the maximum activity in summer and then dropped again in fall.
The highest activity was 58.3 μg triphenyl formazan g-1 soil h-1 in September that were significantly different (p<0.01) with other sampling times. The lowest one was 35.31 μg triphenyl formazan g-1 soil h-1 in November at the undisturbed site. At the disturbed site the highest activity was 33.46 μg triphenyl formazan g-1 soil h-1 in September and the lowest was 21.8 μg triphenyl formazan g-1 soil h-1 in November. In this site dehydrogenase activity in November was significantly (p<0.01) lower than September.
Dehydrogenase activities as affected by soil depth: Dehydrogenase activity remarkably (p<0.01) decreased with depth increment from 57.15 to 13.47 and to 3.06 μg triphenyl formazan g-1 soil h-1 at 0-10, 10-20 and 20-30 cm, respectively (Table 5). This trend was also observed in the disturbed site. The enzyme activity decreased with depth in soil profile from 29.43 to 14.18 and to 6.76 μg triphenyl formazan g-1 soil h-1 at 0-10, 10-20 and 20-30 cm depths, respectively. Dehydrogenase activity at 10-20 and 20-30 cm depths at the disturbed site was more than the undisturbed site. This difference was
significant (p<0.01) at the 20-30 cm depth (Table 7).
DISCUSSION
In our study, the undisturbed site represented the undisturbed system with the higher levels of microbial biomass C and total C content when compared to the disturbed site. Similarly, other studies have found higher microbial biomass in undisturbed systems like native pasture in compared to touched ones like the agricultural systems (Acosta-Martínez et al., 2004). The undisturbed site showed higher total C content and microbial biomass than disturbed site most likely due to positive impact surface cover, vegetation, on soil properties (Acosta-Martínez et al., 2004) and due to differences in their root systems and root exudates of different systems (Fang et al., 2001).
Enzyme activities under undisturbed and disturbed sites: In this study, the activities of acid phosphatase, alkaline phosphatase and dehydrogenase were higher at undisturbed forest sites compared to the disturbed forest sites due to the higher organic C content in the undisturbed forest sites. In addition, microbial biomass which is the principal source of enzymes in soils was greater at undisturbed site than disturbed forest soils (Acosta-Martínez et al., 2007). Previously, it has been reported that greater microbial biomass correlates with organic carbon matter content (Moore et al., 2000). Generally, enzyme activities are correlated to soil organic matter content because the latter plays a key role as a precursor for enzyme synthesis and in enzyme physical stabilization (Tabatabai, 1994).
During the time of this study, alkaline phosphatase activities were up to 2.1 to 2.93 times greater than acid phosphatase at both studied sites. The observed differences are related to different origin of these enzymes in soils. For example plant roots are major source of acid phosphatase in soil (Speir and Cowling, 1991; Dinkelaker and Marschner, 1992) while soil microorganisms (including bacteria, fungi and fauna) are major source of alkaline phosphatase in soil (Findenegg and Neiemans, 1993; Tarafdar, 1995). The higher activity of alkaline phosphatase activity of undisturbed site may be related to the higher microbial biomass (Table 1). Furthermore, alkaline phosphatase generally exceeds acid phosphatase activity in high pH soils (Eivazi and Tabatabai, 1977). Therefore, soil microbes appear to be the important producers of phosphatase in these oak forest soils.
Enzyme activities as affected by seasonal variation: Alkaline and acid phosphatase activity showed a seasonal pattern with maxima in summer and minimum in autumn, suggesting that temperature might be one of the major controlling factors for these enzymes activities. Similar results were reported to find maximum microbial population and enzyme activities in the summer (Tiwari et al., 1989; Rastin et al., 1988; Kaiser and Heinemeyer, 1993). But Boerner et al. (2005) did not observe seasonal effects on the activities of all of enzymes whose activity they measured such as acid phosphatase. Dick et al. (1988) observed little seasonality in acid phosphatase or b-glucosidase activity in North American agricultural fields. The differences in seasonal variation among the enzyme activities studied in this forest are due to their different origin. For example, dehydrogenase and alkaline phosphatase activities were generally higher in September compared to acid phosphatase activity, which was generally higher during July. These findings may demonstrate the beneficial effects of higher soil moisture and lower soil temperatures during September by the microbial populations, which are more directly, correlated to dehydrogenase and alkaline phosphatase activities than acid phosphatase. In the other hand, acid phosphatase, which is thought to be more extracellular in nature, could have retained it activity due to the organic matter protection during July.
Enzyme activities as affected by soil depth: The decreases observed in alkaline phosphatase, acid phosphatase and dehydrogenase activities with soil depth at the undisturbed sites may be related to the higher abundance of soil microorganisms and organic matter content at shallower soil layers of non-disturbed sites. Especially, forests soil surface tend to have high organic matter content compared to other systems such as agricultural and pasture soils (Lavahun et al., 1996). These results were in agreement with the literature data in forest and agriculture soils (Ekelund et al., 2001; Aon et al., 2001; Taylor et al., 2002; Zaman et al., 2002; Turner et al., 2002; Baum et al., 2003; Chen, 2003). At the disturbed forest site, acid phosphatase activity did not vary with soil depths while alkaline phosphatase activities increased with soil depth. The opposite trend found at the disturbed site may demonstrate an irregular distribution of soil microorganisms in disturbed regions compared to undisturbed regions.
CONCLUSION
Our study found differences in microbial biomass, organic matter, acid and alkaline phosphatase and dehydrogenase activities between undisturbed and disturbed forest soils under oak that may be related to the disturbing of forest areas influencing the soils quality negatively. Thus, the significant reductions in soil enzyme activities under undisturbed area compared to disturbed areas should be taken in consideration as indicators of soil quality in the forest areas. Higher soil enzyme activities and microbial biomass were encouraged by the preserving management that may lead to increases in other soil quality parameters such as organic matter content, aggregation and soil water infiltration, soil sustainability and productivity and consequently soil and ecosystem functioning.
ACKNOWLEDGMENTS
The authors are grateful to M. Khoshnevis, H. Jahanbazi and Y. Iranmanesh for their technical support and Dr. E. Sharifi for helping in statistical analysis. This study was supported by Research Institute of Forests and Rangelands in Iran and Austrian Academic Exchange Service.
REFERENCES
- Acosta-Martinez, V., T.M. Zobeck and V. Allen, 2004. Soil microbial, chemical and physical properties in continuous cotton and integrated crop-livestock systems. Soil Sci. Soc. Am. J., 68: 1875-1884.
CrossRefDirect Link - Acosta-Martínez, V., L. Cruz, D. Sotomayor-Ramírez and L. Pérez-Alegría, 2007. Enzyme activities as affected by soil properties and land use in a tropical watershed. Applied Soil Ecol., 35: 35-45.
CrossRefDirect Link - Adams, M.A., 1992. Phosphatase activity and phosphorus fractions in Karri (Eucalyptus diversicolor F. Muell.) forest soils. Biol. Fert. Soils, 14: 200-204.
CrossRefDirect Link - Rao, M.A., A. Violante and L. Gianfreda, 2000. Interaction of acid phosphatase with clays, organic molecules and organo-mineral complexes: Kinetics and stability. Soil Biol. Biochem., 32: 1007-1014.
CrossRefDirect Link - Aon, M.A., M.N. Cabello, D.E. Sarena, A.C. Colaneri, M.G. Franco, J.L. Burgos and S. Cortassa, 2001. I. Spatio-temporal patterns of soil microbial and enzymatic activities in an agricultural soil. Applied Soil Ecol., 18: 239-254.
CrossRefDirect Link - Baum, C., P. Leinweber and A. Schlichting, 2003. Effects of chemical conditions in re-wetted peats temporal variation in microbial biomass and acid phosphatase activity within the growing season. Applied Soil Ecol., 22: 167-174.
CrossRefDirect Link - Boerner, R.E.J., J.A. Brinkman and A. Smith, 2005. Seasonal variations in enzyme activity and organic carbon in soil of a burned and unburned hardwood forest. Soil Biol Biochem., 37: 1419-1426.
CrossRefDirect Link - Chen, H.J., 2003. Phosphatase activity and P fractions in soils of an 18-year-old Chinese fir (Cunninghamia lanceolata) plantation. For. Ecol. Manage., 178: 301-310.
CrossRefDirect Link - Clarholm, M., 1993. Microbial biomass P, labile P and acid phosphatase activity in the humus layer of a spruce forest, after repeated additions of fertilizers. Biol. Fert. Soils, 16: 287-292.
CrossRefDirect Link - Dick, R.P., P.E. Rasmussen and E.A. Kerle, 1988. Influence of long-term residue management on soil enzyme activities in relation to soil chemical properties of a wheat-fallow system. Biol. Fert. Soils, 6: 158-164.
CrossRefDirect Link - Dick, W.A., L. Cheng and P. Wang, 2000. Soil acid and alkaline phosphatase activity as pH adjustment indicators. Soil Biol. Biochem., 32: 1915-1919.
CrossRef - Dinkelaker, B. and H. Marschner, 1992. In vivo demonstration of acid phosphatase activity in the rhizosphere of soil-grown plants. Plant Soil, 144: 199-205.
CrossRefDirect Link - Eivazi, F. and M.A. Tabatabai, 1977. Phosphatases in soils. Soil Biol. Biochem., 9: 167-172.
CrossRef - Ekelund, F., R. Ronn and S. Christensen, 2001. Distribution with depth of protozoa, bacteria and fungi in soil profiles from three Danish forest sites. Soil Biol. Biochem., 33: 475-481.
CrossRefDirect Link - Fang, C., M. Radosevich and J.J. Fuhrmann, 2001. Characterization of rhizosphere microbial community structure in five similar grass species using FAME and BIOLOG analyses. Soil Biol. Biochem. 33: 679-682.
CrossRefDirect Link - Findenegg, G.R. and J.A. Neiemans, 1993. The effect of phytase on the availability of P from myo-inositol hexaphosphate (phytate) for maize roots. Plant Soil, 154: 189-196.
CrossRef - Garcia, C., T. Hernandez and F. Costa, 1997. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plant Anal., 28: 123-134.
CrossRefDirect Link - Herbien, S.A. and J.L. Neal, 1990. Soil pH and phosphatase activity. Commun. Soil Sci. Plant Anal., 21: 439-456.
CrossRefDirect Link - Joergensen, R.G. and P.C. Brookes, 1990. Ninhydrin-reactive nitrogen measurements of microbial biomass in 0.5 m K2SO4 soil extracts. Soil Biol. Biochem., 22: 1023-1027.
CrossRefDirect Link - Jordan, D., R.J. Kremer, W.A. Bergfield, K.Y. Kim and V.N. Cacnio, 1995. Evaluation of microbial methods as potential indicators of soil quality in historical agricultural fields. Biol. Fert. Soil, 19: 297-302.
CrossRef - Kaiser, E.A. and O. Heinemeyer, 1993. Seasonal variations of soil microbial biomass carbon within the plough layer. Soil Biol. Biochem., 25: 1649-1656.
CrossRef - Kilmer, V.J. and L.T. Alexander, 1949. Methods of making mechanical analysis of soils. Soil Sci., 68: 15-24.
Direct Link - Lavahun, M.F.E., R.G. Joergensen and B. Meyer, 1996. Activity and biomass of soil microorganisms at different depths. Biol. Fert. Soils, 23: 38-42.
CrossRef - Li, M.G., M. Osaki, M. Honma and T. Tadano, 1997. Purification and characterization of phytase induced in tomato roots under phosphorus-deficient conditions. Soil Sci. Plant Nutr., 43: 179-190.
CrossRefDirect Link - Moore, J.M., S. Klose and M.A. Tabatabai, 2000. Soil microbial biomass carbon and nitrogen as affected by cropping systems. Biol. Fertil. Soils, 31: 200-210.
CrossRefDirect Link - Olsen, S.R., C.V. Cole, F.S. Watanabe and L.A. Dean, 1954. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. U.S. Department of Agriculture, Washington, D.C., Pages: 19.
Direct Link - Rastin, N., K. Rosenplanter and A. Huttermann, 1998. Seasonal variation of enzyme activity and their dependence on certain soil factors in a beech forest soil. Soil Biol. Biochem., 20: 637-642.
CrossRef - Schloter, M., O. Dilly and J.C. Munch, 2003. Indicators for evaluating soil quality. Agric. Ecosyst. Environ., 98: 255-262.
CrossRefDirect Link - Speir, T.W. and J.C. Cowling, 1991. Phosphatase activities of pasture plants and soils: Relationship with plant productivity and soil P fertility indices. Biol. Fert. Soils, 12: 189-194.
CrossRef - Tarafdar, J.C., 1995. Visual demonstration of in vivo acid phosphatase activity of VA mycorrhizal fungi. Curr. Sci., 69: 541-543.
Direct Link - Tarafdar, J.C., R.S. Yadav and S.C. Meena, 2001. Comparative efficiency of acid phosphatase originated from plant and fungal sources. J. Plant Nutr. Soil Sci., 164: 279-282.
CrossRef - Taylor, J.P., B. Wilson, M.S. Mills and R.G. Burns, 2002. Comparison of microbial numbers and enzymatic activities in surface soils and subsoils using various techniques. Soil Biol. Biochem., 34: 387-401.
Direct Link - Turner, B.L., R. Baxter and B.A. Whitton, 2002. Seasonal Phosphatase activity in three characteristic soils of the English uplands polluted by long-term atmospheric nitrogen deposition. Environ. Pollut., 120: 313-317.
CrossRefPubMedDirect Link - Visser, S. and D. Parkinson, 1992. Soil biological criteria as indicators of soil quality: Soil microorganisms. Am. J. Alternative Agric., 7: 33-37.
Direct Link - Walkley, A. and I.A. Black, 1934. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci., 37: 29-38.
CrossRefDirect Link - Zaman, M., K.C. Cameron, H.J. Di and K. Inubushi, 2002. Changes in mineral N microbial and enzyme activities in different soil depths after applications of dairy shed effluent and chemical fertilizer. Nutr. Cycl. Agroecosyst., 63: 275-290.
CrossRefDirect Link