
M. Sie
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin
Y. Sere
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin
S. Sanyang
Forum for Agricultural Research in Africa (FARA), P.O. Box 1628, Accra, Ghana
L.T. Narteh
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin
S. Dogbe
Institut Togolais de Recherche Agronomique (ITRA), BP 1163, Lomé, Togo
M.M. Coulibaly
Institut d`Economie Rurale (IER), BP 258, Bamako, Mali
A. Sido
Institut National de Recherches Agronomiques du Niger (INRAN), BP 429, Niamey, Niger
F. Cisse
Institut d`Economie Rurale (IER), BP 258, Bamako, Mali
E. Drammeh
National Agricultural Research Institute (NARI), P.M.B. 526, Serrekunda, Gambia
S.A. Ogunbayo
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin
L. Zadj
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin
B. Ndri
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin
B. Toulou
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 2 | Page No.: 130-139







ABSTRACT
The immense potential of the lowlands in West and Central Africa for durable intensification of rice cropping have not been realised due to biotic and abiotic constraints. There is a need to replace existing rice varieties with others that are better adapted to the lowland conditions. After the success of the upland interspecific varieties, Africa Rice Center (WARDA) and its partners developed NERICA varieties suitable for irrigated and rainfed lowlands. The stable varieties resulting from this work were evaluated under preliminary yield trials in eight countries at 19 sites. The entries included 61 interspecific (O. glaberrima x O. sativa indica) varieties and 9 intraspecific (O. sativa indica x O. sativa indica) varieties. The aim of the study is to introduce new lowland NERICAs through a participatory approach and to identify ideotypes that are adapted to lowland conditions. Variations did exist among the 73 rice varieties with respect to the five traits that were evaluated. Total number of tillers, panicle number and flowering dates were observed to greatly influence the yield among the 73 varieties that were evaluated. A principal components plot clustering analysis were used to group the accessions. The interspecific varieties formed the most interesting group and have a better capacity for adaptation to the diversity of lowlands. They have acceptable yields, sometimes higher than those of intraspecific varieties and checks. Thus, most lowland NERICAs varieties tested in three ecologies could produce more than 5 t ha-1. The results obtained were quite encouraging and showed that, the varieties possess good agronomic traits that are well adapted to intensified lowland rice farming. The recent naming of some of these interspecific varieties as NERICA-L (New Rice for Africa Lowland) by Africa Rice Center has confirmed that they compare well with the traditional varieties. Thus, from this study, we now have a new set of interspecific varieties that are adapted to lowland conditions and which the national research programs can use in various tests for satisfying farmers` needs.
PDF Abstract XML References Citation
How to cite this article
M. Sie, Y. Sere, S. Sanyang, L.T. Narteh, S. Dogbe, M.M. Coulibaly, A. Sido, F. Cisse, E. Drammeh, S.A. Ogunbayo, L. Zadj, B. Ndri and B. Toulou, 2008. Regional Yield Evaluation of the Interspecific Hybrids (O. glaberrimax O. sativa) and Intraspecific (O. sativa x O. sativa) Lowland Rice. Asian Journal of Plant Sciences, 7: 130-139.
DOI: 10.3923/ajps.2008.130.139
URL: https://scialert.net/abstract/?doi=ajps.2008.130.139
DOI: 10.3923/ajps.2008.130.139
URL: https://scialert.net/abstract/?doi=ajps.2008.130.139
INTRODUCTION
Rice is one of the most important crops that provide food for about half of the world population, particularly in Asia, Africa and Latin America where the demand for rice is a top priority. Its adoption as a principal staple food is increasing in Africa (Sasaki, 1999, 2002; Sie et al., 2005; Rodenburg et al., 2006; Ogunbayo et al.,2007). Rice production in Africa increased from 8.6 million tones of paddy in 1980 to 18.6 million tones in 2005. Despite such dramatic growth, demand continues to exceed supply and the region relies on imported rice (WARDA, 2004, 2006; Ogunbayo et al., 2005). In West Africa alone there are an estimated 20 million hectares of cultivable lowlands. If just 2 million hectares of lowlands is grown to rice, producing an average yield of only 3 tonnes per hectare, the region could easily stop its costly rice import. The lowlands, therefore, offer great potential for the sustainable expansion and intensification of rice and can help to feed the growing population in the region. The immense potential of the lowlands in West and Central Africa for durable intensification of rice cropping have not been realised due to biotic and abiotic constraints (WARDA, 2006).
In Africa, there are two major cultivated rice species, Oryza glaberrima (Steud) and Oryza sativa (L.), both species have distinct and complementary advantages and disadvantages for use in African farming systems (Rodenburg et al., 2006). The Asian rice (O. sativa) has good agronomic traits but susceptible to most African stresses (Pernes et al., 1984; Semon et al., 2005). African indigenous species (O. glaberrima) is an interesting genetic resource due to its resistance to many of rice constraints (Sie, 1991; Jones et al., 1997b; Johnson et al., 1998; Jones and Singh, 1999; Sie, 1999; Futakuchi et al., 2001).
In 1992, the Africa Rice Center (WARDA) and its partners started the Interspecific Hybridization Project (IHP) in an attempt to combine the useful traits of both cultivated rice species (O. sativa and O. glaberrima). Crossing the two species was complicated by their incompatibility, which led to hybrid mortality. This problem was overcome through backcrossing with the O. sativa parent coupled with anther culture, resulting in the first interspecific rice progenies from cultivated varieties (Jones et al., 1997a,c).
In addition to the upland NERICA varieties, WARDA and national programs of West African countries developed NERICA varieties suitable for irrigated and rainfed lowlands, one of the most complex rice ecologies in the world. Key to this success was the unique R and D partnership model forged between WARDA and the national programs in West and central African countries through the Rice Research and Development Network for West and Central Africa (ROCARIZ), which facilitated the shuttle-breeding approach to accelerate the selection process and achieve wide adaptability of the lowland NERICAs (WARDA, 2006).
The stable varieties resulting from this research were evaluated in 2004 wet season under preliminary yield trial in eight countries at 19 sites with support of ROCARIZ. The sites were chosen to include a diversity of ecologies- valley fringe, valley bottom and irrigated lowlands. The goal of this study was to introduce new lowland NERICAs through a participatory approach and to identify ideotypes that are adapted to lowland conditions.
MATERIALS AND METHODS
The study was conducted in 2004 wet season in 8 countries: Burkina Faso (2 sites at Banfora), Côte d`Ivoire (4 sites, 2 at M`bé and 2 at Gagnoa), Gambia (1 site at Niani Sukuta), Mali (3 sites, 2 at Sikasso and 1 at Niono) Niger (2 sites, Sébéry and Bonfeba), Nigeria (4 sites, 3 in Ibadan and 1 in Edozhigi), Togo (2 sites, Bémé and Atime), Guinea (1 site). The sites were chosen to include a diversity of ecologies-valley fringe, valley bottom and irrigated lowlands. The entries included 61 interspecific (O. glaberrima x O. Sativa indica) varieties and 9 intraspecific (O. Sativa indica x O. Sativa indica) fixed varieties from Burkina-Faso, Mali, Senegal and Togo. The check varieties were FKR 19 for the valley fringe and FKR 54 for the irrigated and valley bottom ecologies, while the third check served as a reference for each site and was optional. The infesting band comprised three varieties (ITA 306, BOUAKE 189, TOX 3055-10-1-1-1) susceptible to AfRGM, RYMV and Blast. The checks were placed after every 20, 40 and 60 test varieties.
Simple non-replicated block design was used with five rows of 5 m each and entries were sown perpendicular to an infected variety susceptible to AfRGM, RYMV and Blast. A pre-planting base application of 200 kg ha-1 of NPK (15-15-15) was made, followed by a total of 100 kg ha-1 of urea in two applications at panicle initiation and booting stages. Morphological data were collected for 5 quantitative characters at appropriate growth stage of rice plant following the Standard Evaluation Systems for rice (IRRI, 1996). Traits evaluated were plant height at maturity; tillering at 60 days after sowing; days to flowering; number of panicles per m2 and yield. The data collected on 5 agro-botanical traits from the genotypes were subjected to statistical analysis using SAS/PC version 9.1 (SAS, 1999) and GGE biplot version 5.2 (Yan and Kang, 2003). A GGE biplot was constructed using the first two principal components (PC1 and PC2) derived from subjecting the environment-centered data to singular-value decomposition and it has many visual: (i) The polygon view of a GGE biplot allows visualization of the which-won-where pattern (i.e., which variety had the highest yield in which environment) (ii) the average environment coordination view allows simultaneous visualization of the mean performance and stability of the treatments, the discriminating ability vs. representativeness of the environments and (iii) the environment vector view allows visualization of the interrelationship among environments (Yan, 2001, 2002; Yan and Kang, 2003). In addition, attempting to characterize the environments and to relate the mean yield of the environments to three ecologies, a biplot based on an environment x factor two-way table was constructed, which was similar to that based on a genotype x trait two-way table described by Yan and Rajcan (2002). All biplots presented in this paper were generated using the software GGEbiplot package that runs in a Windows environment.
Principal components grouping of the traits was employed to examine the percentage contribution of each trait to total genetic variation and to spot characters that reflected the greatest proportion of variations among the 5 variables. This is because the PCA has been reported to be able to choose independent (orthogonal) axes that are minimally correlated and then represent linear combinations of the original characters (Akoroda, 1983).
The relative discriminating power of the axes and their associated characters were measured by the Eigen values and factor scores, respectively. Cluster analysis based on hierarchical matrices was also employed on agro-botanical data using SAS program.
RESULTS
The correlation matrix (Table 1) showed that flowering date was positively and significantly associated with number of panicles m-2, yield and number of tillers. However, plant height had negative but significant association with panicle m-2 and positive significant and association with yield. Panicle m-2 was positively and significantly correlated to yield and number of tiller. Also, yield were also positively and significantly correlated to number of tillers.
Table 2 shows the principal components analysis showing the contribution (factor scores) of each character to total variation among the 73 varieties. The three principal components axes accounted for about 79.53% of total variance with the first and second principal components taking 37.89 and 22.52%, respectively. The relative discriminating power of the principal axes as indicated by the Eigen values was highest (1.89) for axis 1 and lowest (0.96) for axis 3. The first principal component that accounted for the highest proportion (37.89%) of total variation was mostly correlated with panicle number, total number of tillers and flowering days, suggesting that the varieties had good tillering ability, short height, high panicle number and good yield. Characters that were mostly correlated with the second principal component were plant height, yield, panicle number and flowering days.
Table 3 shows characteristics of morphological groups defined by topology as each of the three groups has its own distinct characteristics. In group 1 plant height at maturity ranged from 55 to 117 cm indicating that varieties are semi - dwarf to medium height type; number of tillers after 60 days ranged from 23 to 375 tillers m-2; flowering occurred between 70 and 103 days; there were 85 to 598 panicles m-2 (suggesting the development of productive secondary tillers) while grain yield ranged from 181 to 8,844 kg ha-1. In group 2 plant height at maturity ranged from 57 to 152 cm indicating that the accessions are semi - dwarf to tall height type. Number of tillers after 60 days ranged from 48 to 663 tillers m-2; flowering occurred between 76 and 129 days; there were 106 to 576 panicles m-2, indicating a sustained development of productive secondary tillers; while grain yield ranged from 290 to 9,119 kg ha-1.
| Table 1: | Correlation coefficients of five traits used in characterizing seventy-three rice varieties |
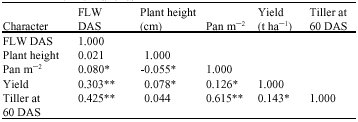 | |
| *, **: Significant at 5 and 1% probability levels, respectively | |
| Table 2: | Principal components analysis showing the contribution (factor scores) of each character among the seventy-three varieties |
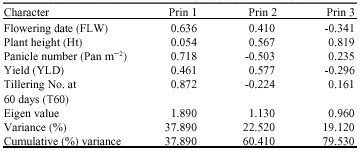 | |
| Table 3: | Characteristics of morphological groups defined by topology |
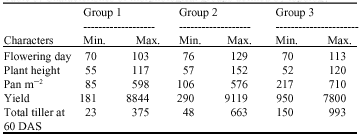 | |
In group 3, plant height at maturity ranged from 52 to 120 cm indicating that the lines are semi-dwarf to medium height type; number of tillers after 60 days ranged from 150 to 993 tillers m-2; flowering occurred between 70 and 113 days; there were 217 to 710 panicles m-2, indicating sustained development of productive secondary tillers; while grain yield ranged from 950 to 7,800 kg ha-1.
Table 4 shows performance of selected varieties across valley bottom and irrigated ecologies. The results showed that, WAS 122-IDSA-10-WAS-3-1-TGR 3 in group 2 has the highest yield (7159 kg ha-1) followed by WAS 161-B-4-B-2 (7145 kg ha-1). Generally, most of lowland NERICA performed better than checks.
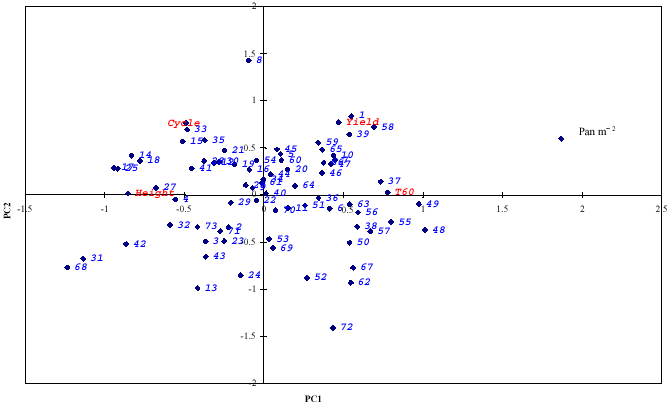
A plot of relationship between the 73 varieties as shown by the first and second principal components axes (Prin 1 and 2) (Fig. 1) revealed that all the varieties were ordered into four distinct PCA clusters. Most of the varieties in group 2 were centrally distributed more towards the first principal axis and they have good yield.
| Table 4: | Performance of selected varieties across valley bottom and irrigated lowland ecologies |
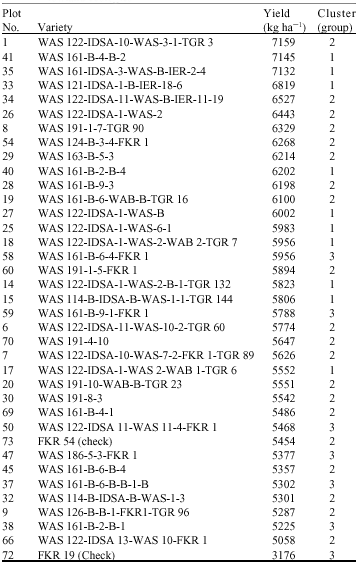 | |
However, accessions selected from group 1 were also distributed towards the upper left side of first principal axis while group three were distributed towards the lower right side.
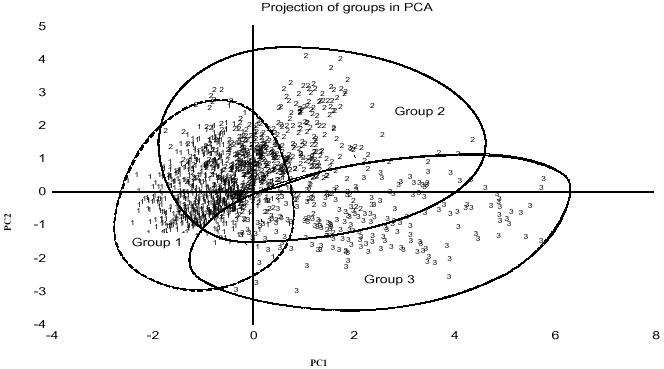
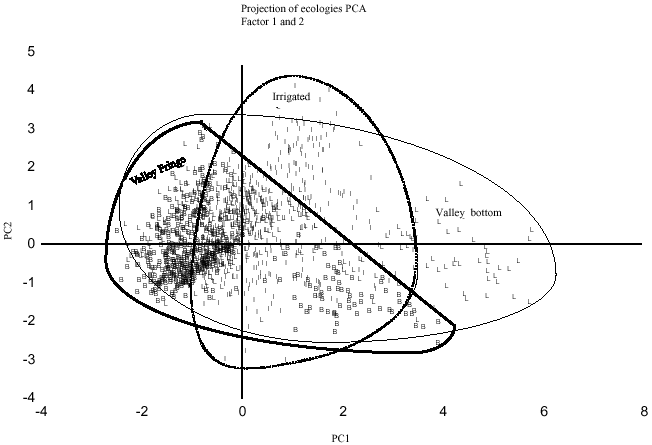
Figure 2 shows classification of the genotypes by principal components analysis obtained from the similarity matrix. The PCA grouped 73 varieties according to their similarity suggesting that close genetic relationships was associated with close morphological relationship. Figure 3 shows classification of ecologies and may be used in planned crosses, if one to increase the genetic basis of lowland germplasm.
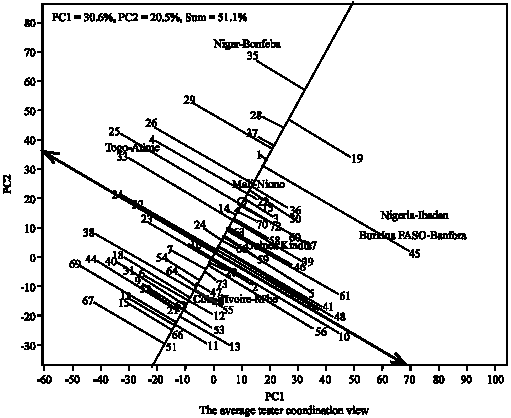
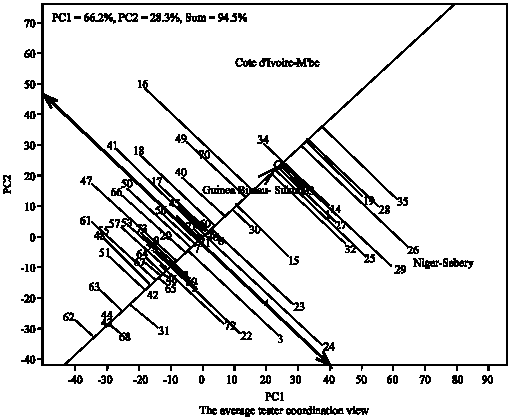
Figure 4 shows the average tester coordination view, showing the performance of the genotypes across the locations and their stability under valley bottom condition. Visualization of the mean and stability of genotypes is achieved by drawing an Average Environment Coordinate (AEC) on the genotype-focused biplot. First, an average environment, represented by the small circle, is defined by the mean PC1 and PC2 scores of the environments. The small circle near Mali-Niono location suggests Niono site as the average location in relation to other locations in term of yield performance. The line connecting the biplot origin and the circle (Niono) is referred to as the average-tester axis. Based on their mean performance, the genotypes are ranked along the average-tester axis with the arrow pointing towards genotype with greater value. Thus, the genotype are ranked along the AEC abscissa, with the arrow pointing to higher mean performance.
Based on this, Genotype (35) was clearly the highest-yielding, on average, in these environments, followed by (19, 28, 27, 29, 1, 45, 26, 36, 22 and 30) and the least in terms of performance was the genotype (67). The double arrow indicates that a greater projection onto the AEC ordinate, regardless of the direction, means greater instability. Therefore, genotype (4) near the top and (3) near the bottom of the biplot are more variable and less stable than other genotypes.
Figure 5 showing the performance of the genotypes across the locations and their stability under irrigated ecology. The small circle near Niger-Sebery and Côte d`Ivoire locations suggests Sebery and M`be sites as the average locations in relation to other locations in term of yield performance. Genotype (35) was ranked first and the least in terms of performance was the genotype (62).
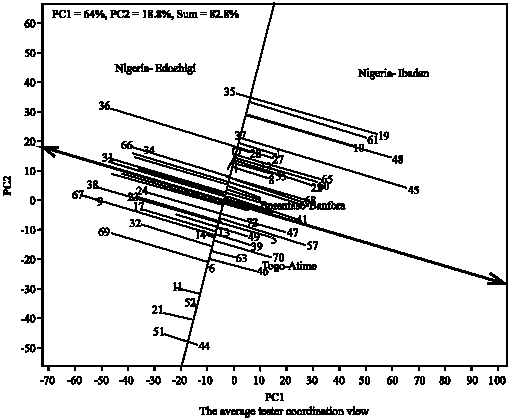
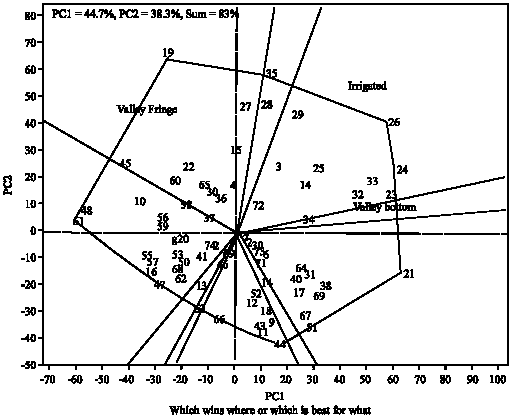
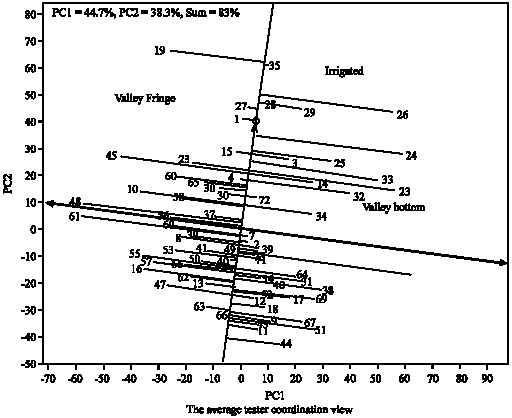
Figure 6 showing the performance of the genotypes across the locations and their stability under valley fringe ecology. The small circle near Nigeria-Edozhigi and Nigeria-Ibadan locations suggests Edozhigi and Ibadan sites as the average locations in relation to other locations in term of yield performance. Genotype (35) was ranked first and the least in terms of performance was the genotype (44). Figure 7 defined the genotypes that performed best in the three ecologies (which genotype won in which ecology). The polygon is formed through connecting the best genotypes in each ecology to other. Starting from the biplot origin, perpendicular lines are drawn to each side of the polygon, which divide the biplot into 10 sectors. The which-won-where pattern is examined as follows. The varietal number at the vertex of polygon in any sector is the genotype that produces the highest yields in all ecologies that fall in that sector. Thus, genotypes (3, 14, 23, 24, 25, 26, 29, 32, 33, 34 and 72) won in irrigated and valley bottom while genotype (26) produced the highest yields in two ecologies. The genotypes that won in valley fringe are: 1, 4, 15, 22, 27, 30, 36, 37, 45, 58, 60 and 65. The genotype (19) produced the highest yield in valley fringe ecology. Out of the three checks (i.e., 71, 72 and 73) used in the study only one check (72) performed well under irrigated and valley bottom ecologies the remain two checks (71 and 73) with other genotypes did not seem to perform well in any of the ecologies (they did not win in any location). Figure 8 showing the performance of the genotypes and their stability across the irrigated, valley bottom and valley fringe ecologies. The bi-plot confirmed the conclusions drawn from Fig. 1-7.
 | |
| Fig. 1: | Plot of Prin 1 and Prin 2 showing the relationship between clusters of 73 varieties |
 | |
| Fig. 2: | Classification of the genotypes by principal components analysis obtained from the similarity matrix |
 | |
| Fig. 3: | Classification of the ecologies by analysis of principal components obtained from the similarity matrix |
 | |
| Fig. 4: | The mean performance vs. stability of the 73 rice varieties across the seven test locations under valley bottom condition |
 | |
| Fig. 5: | The mean performance vs. stability of the 73 rice varieties across the three test locations under irrigated lowland condition |
 | |
| Fig. 6: | The mean performance vs. stability of the 73 rice varieties across the four test locations under valley fringe condition |
 | |
| Fig. 7: | A polygon view of the GGE biplot of genotypexecology, showing which genotype won where or best for which ecology |
 | |
| Fig. 8: | The mean performance vs. stability of the 73 rice varieties across the irrigated lowland, valley bottom and valley fringe ecologies |
DISCUSSION
Variations did exist among the 73 genotypes with respect to the five traits that were evaluated. Total number of tillers, panicle number and flowering days were observed to greatly influence the yield among the 73 genotypes that were evaluated. Thus it was not surprising that positive and significant associations were observed between yield and days to flowering (r = 0.303); panicle number (r = 0.126) and total number of tiller (r = 0.425). In addition, the hierarchical cluster groupings of accessions, confirmed the contributions of the three traits to grain yield among the 73 genotypes. The implication is that if selection is to be made between cluster groups for a future breeding exercise, panicle number, total number of tillers, days to heading, should be given high priorities. The interspecific varieties formed the most interesting group and have a better capacity for adaptation to the diversity of lowlands. They have acceptable yields, sometimes higher than those of intraspecific varieties and checks. Thus, most genotypes tested in three ecologies can produce more than 5 t ha-1.
Principal components analysis and hierarchical clustering generated from similarity or genetic distance matrices has provided an overall pattern of variation as well as the degree of relatedness among the genotypes. The results indicate that the genotypes in group 1 will be adapted to the low-input cropping system of the valley fringe, while those in group 3 would be more appropriate for intensive cultivation in the valley bottom and irrigated lowland ecologies. The GGE bi-plot generated several graphic bi-plots, strong genotype by environment interaction was confirmed. It also revealed the relationship among genotypes in terms of their responses and stability to the environments and ecologies. The results confirmed that the interspecific genotypes performed well across the locations and they were very stable. The majority of the genotypes are derived from crosses involving Tog 5681 and IR 64 as parents and crossing numbers are WAS 161 and WAS 122 and were selected in Burkina Faso. Five among them (WAS 122-IDSA 10-WAS 1-1-FKR 1, WAS 122-IDSA 10-WAS 6-1-FKR 1, WAS 122-IDSA 11-4-FKR 1, WAS 122-IDSA 14-WAS-B-FKR 1 and WAS 122-IDSA 11-WAS-8-2) were rated at Institut des Savanes (IDESSA), Côte d`Ivoire as RYMV-tolerant. Interestingly, interspecific varieties produced the greatest number of tillers and these observations confirm those made by Jones et al. (1996), who noted that interspecific O. glaberrima x O. sativa had a very high tillering capacity, which predisposed them to be more competitive with weeds.
In conclusion, the results obtained were quite encouraging and showed that, the varieties possess good agronomic traits that are well adapted to intensified lowland rice farming. The recent naming of some of these interspecific varieties as NERICA-L (New Rice for Africa Lowland) by WARDA has confirmed that they compare well with the traditional varieties. Based on the topology of the varieties, it is concluded that the interspecific crossings O. glaberrima x O. sativa indica can increase lowland rice biodiversity. Thus from this study, we now have a new set of interspecific lines that are adapted to lowland conditions and which the national research programs can use in various tests for satisfying farmers` needs.
REFERENCES
- Akoroda, M.O., 1983. Principal components analysis and metroglyph of variation among Nigerian yellow yams. Euphytica, 32: 565-573.
CrossRefDirect Link - Futakuchi, K., M.P. Jones and R. Ishii, 2001. Physiological and morphological mechanisms of submergence resistance in African rice (Oryza glaberrima). Jap. J. Trop. Agric., 45: 8-14.
Direct Link - IRRI, 1996. Standard Evaluation System for Rice. 4th Edn., International Rice Research Institute, Manila, Philippines, Pages: 52.
Direct Link - Jones, M.P., A. Audebert, S. Mande and K. Aluko, 1996. Characterization and utilization of Oryza glaberrima Steud. In upland rice breeding. Proceedings of the Workshop of Africa-Asia Joint Research on Interspecific Hybridisation Between the African and Asian Rice Species O. glaberrima and O. sativa, WARDA, December 16-18, 1996, Bouake, pp: 43-59.
- Jones, M.P., M. Dingkuhn, G.K. Aluko and M. Semon, 1997. Interspecific Oryza sativa L. X O. Glaberrima steud. Progenies in upland rice improvement. Euphytica, 94: 237-246.
CrossRefDirect Link - Jones, M.P., S. Mande and K. Aluko, 1997. Diversity and potential of Oryza glaberrima Steud. in upland rice breeding. Breed. Sci., 47: 395-398.
Direct Link - Johnson, D.E., M. Dingkuhn, M.P. Jones and M.C. Mahamane, 1998. The influence of rice plant type on the effect of weed competition on Oryza sativa and Oryza glaberrima. Weed Res., 38: 207-216.
CrossRefDirect Link - Ogunbayo, S.A., D.K. Ojo, R.G. Guei, O.O. Oyelakin and K.A. Sanni, 2005. Phylogenetic diversity and relationship among forty rice accessions using Morphological and RAPDs techniques. Afr. J. Biotechnol., 4: 1234-1244.
Direct Link - Ogunbayo, S.A., D.K. Ojo, A.R. Popoola, O.J. Ariyo and M. Sie et al., 2007. Genetic comparisons of landrace rice accessions by morphological and RAPDs techniques. Asian J. Plant Sci., 6: 653-666.
Direct Link - Rodenburg, J., A. Diagne, S. Oikeh, K. Futakuchi and P.M. Kormawa et al., 2006. Achievements and impact of NERICA on sustainable rice production in sub-Saharan Africa. Int. Rice Commission Newslett., 55: 45-58.
Direct Link - Sasaki, T., 2002. Rice genomics to understand rice plant as an assembly of genetic codes. Curr. Sci., 83: 834-839.
Direct Link - Semon, M., R. Nielsen, M.P. Jones and S.R. McCouch, 2005. The population structure of African cultivated rice Oryza glaberrima (Steud.): Evidence for elevated levels of linkage disequilibrium caused by admixture with O. sativa and ecological adaptation. Genetics, 169: 1639-1647.
CrossRefDirect Link - Sie, M., S.Y. Dogbe and M. Coulibaly, 2005. Selection of interspecific hybrids (O. sativa x O. glaberrima or lowland NERICAs) and intraspecifics adapted to rainfed lowland growing conditions. Int. Rice Comm. Newslett., 54: 47-51.
Direct Link - Yan, W., 2001. GGE biplot-a Windows application for graphical analysis of multienvironment trial data and other types of two-way data. Agron. J., 93: 1111-1118.
Direct Link - Yan, W., 2002. Singular-value partitioning in biplot analysis of multienvironment trial data. Agron. J., 94: 990-996.
Direct Link - Yan, W. and I. Rajcan, 2002. Biplot analysis of test sites and trait relations of soybean in Ontario. Crop Sci., 42: 11-20.
CrossRefDirect Link