
C. Bounphanousay
National Agriculture and Forestry Research Institute,Ministry of Agriculture and Forestry, P.O. Box 811, Vientiane, Lao PDR
P. Jaisil
Department of Plant Science and Agricultural Resources,Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
J. Sanitchon
Not Available
M. Fitzgerald
Grain Quality, Nutrition and Postharvest Centre, IRRI,DAPO Box 777, Metro Manila, Philippines
N.R. Sackville Hamilton
T.T. Chang Genetic Resources Center, IRRI,DAPO Box 777, Metro Manila, Philippines
J. Sanitchon
Department of Plant Science and Agricultural Resources,Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 1-7







ABSTRACT
Fifty three accessions of lowland Black Glutinous Rice (BGR) and three white rice varieties from Lao PDR were evaluated for variations in 2-acetyl-1-pyrroline (2-AP), a main aroma compound present in fragrant rice and a 257 bp genomic fragment associated with a gene encoding 2-AP. The objective of this study was to characterize the variation in aromatic characters of Lao BGR by using chemical and molecular techniques. Thirty six out of the 56 accessions had aromatic properties, containing 2-acetyl-1-pyrroline (2-AP). The concentrations of 2-AP in unpolished samples were about 50% higher than those of the polished samples. The concentrations of 2-AP in BGR accessions were lower than those of white aromatic rice varieties in which 2-AP was highest in Kai Noi Leuang (0.688 ppm). Seventeen accessions and two of white rice (Kai Noi Leuang and TDK5) had a common 355 bp allele corresponding to that found in Calrose, a non-fragrant variety. A 257 bp allele was found in the 36 varieties of BGR and a local aromatic white rice variety, Hom Nang Nouan, sharing a common allele as in the fragrant variety KDML 105. Surprisingly, Kai Noi Leuang, a popular Lao local aromatic rice variety with the highest 2-AP concentration did not possess a 257 bp fragment which has been identified previously as a fragrant allele. This is the first study of the chemical and molecular aromatic properties of lowland BGR varieties of the Lao PDR and the data obtained will be valuable in screening other germplasm and in planning a breeding program in the country geared towards fragrant rices with superior agronomic properties.
PDF Abstract XML References Citation
How to cite this article
C. Bounphanousay, P. Jaisil, J. Sanitchon, M. Fitzgerald, N.R. Sackville Hamilton and J. Sanitchon, 2008. Chemical and Molecular Characterization of Fragrance in Black Glutinous
Rice from Lao PDR. Asian Journal of Plant Sciences, 7: 1-7.
DOI: 10.3923/ajps.2008.1.7
URL: https://scialert.net/abstract/?doi=ajps.2008.1.7
DOI: 10.3923/ajps.2008.1.7
URL: https://scialert.net/abstract/?doi=ajps.2008.1.7
INTRODUCTION
Rice is a member of genus Oryza in the grass family (Gramineae) consisting of 22 species in the genus, of which only two are cultivated: O. sativa and O. glaberrima (Khush, 1997). O. sativa was domesticated in South Asia at least 10,000 years ago (Zhang and Jiarong, 1998) and O. glabberima was domesticated in West Africa between about 1500 and 800 BC (Murrey, 2004). Of the 20 wild species of Oryza, five are found in Laos (Kuroda et al., 2006).
South and Southeast Asia is generally accepted as the center of origin and domestication of glutinous (sticky) rice. Black glutinous rices are among the rice cultivars that distribute in many Asian countries and they are present in both indica and japonica background (Chaudhary and Mahyuddin, 2000). Glutinous rice is a single staple food in Lao PDR with the highest per capita production and consumption of glutinous rice in the world (Schiller et al., 2006). As part of the center of origin and domestication, Lao PDR proved to have one of the richest biodiversities in rice (Anonymous, 2007). In Lao PDR, rice cultivars with good aromatic characters are common for both white and black glutinous rices and they are well known among farmers as Khoa Hom or fragrant rices (Appa et al., 2006a).
The demand for fragrant rices for both local and improved varieties has increased markedly in recent years in local and international markets to such an extent that consumers are willing to pay a premium for fragrant rices. Because of this market opportunity, there is a need to validate the data on aromatic traits of Lao BGR in order to assist in the screening, breeding and development of high-yielding fragrant black glutinous rice varieties that also possess other desirable agronomic traits.
The pleasant aroma associated with aromatic varieties is not only released in cooked rice but is also often emitted by these varieties in the field at the time of flowering (Weber et al., 2000; Widjaja et al., 1996). Aroma is caused by extremely small amounts of volatile compounds, which are contained as a complex mixture. More than 100 compounds contributing to the aroma of rice have been identified (Tsugita, 1986; Widjaja et al., 1996). Some of these volatile compounds contribute to consumer acceptance of certain types of rice, whereas other compounds contribute to consumer rejection. The aromatic compound 2-acetyl-1-pyrroline (2-AP) is the primary component responsible for the fragrance of aromatic rice (Weber et al., 2000; Buttery et al., 1988).
Fragrant rice varieties differ in their degree of aroma and are broadly classified as strongly, moderately and weakly scented types (Singh et al., 2003). Khush and De La Cruz (1998) believed that aroma is a quantitative character as segregants with varying levels of aroma have been observed in the crosses between aromatic and non-aromatic varieties. As a quantitative trait, environmental factors are well known to affect the expression of aroma, such as temperature during the late reproductive stages of crop growth, soil properties, agricultural factors grain storage and processing. The potential effects of many of these factors on the expression of aroma and other grain characteristics have been reviewed by several authors (Singh et al., 2003; Khush and Dela Cruz, 1998; Goodwin et al., 1994a, b).
Numbers of genes controlling the inheritance of aromatic characters reported by several investigations vary from one to four pairs (Brijal and Gupta, 1998; Pinson, 1994; Lin, 1991). Bradbury et al. (2005a) reported that fragrance in rice is a recessive trait due to an 8 bp deletion and 3 Single Nucleotide Polymorphisms (SNPs) in chromosome 8 that introduce a stop codon upstream of important amino acid residues there by leading to loss of function mutation of the fragrance (fgr) gene product which expresses a putative Betaine Aldehyde Dehydrogenase (BAD2). This finding is supported by a more recent study which was able to determine the location of the gene using high-density markers, resolving the physical distance by 69 kb. Aside from BAD2, candidate gene analyses revealed two other putative genes, encoding eukaryotic-type carbonic anhydrase and 3-methylcrotonyl-CoA carboxylase beta chain, respectively (Chen et al., 2006). Based on recent studies, it appears that there is one major gene responsible for aroma but there might be several modifiers or quantitative trait loci involved in this commercially important trait.
Different methods have been used for studying fgr gene in rice and these methods facilitate marker-assisted selection of the aromatic traits. Genotyping strategies include the use of SSRs (Chen et al., 2006; Jain et al., 2006; Cordeiro et al., 2002) as well as SNPs (Jin et al., 2003). Bradbury et al. (2005b) designed a rapid and easy method of detecting the 8-bp deletion using Allele-Specific Amplification (ASA). This genotyping method is very convenient because it is simply resolved in agarose gel and it can distinguish between fragrant and non-fragrant rice varieties and can identify homozygous fragrant, homozygous non-fragrant and heterozygous individuals.
A total number of 13,992 cultivated rice accessions have been collected in during 1995 to 2003 (Appa et al., 2002a, 2002b). Of the total number, 477 samples were classified as aromatic rice mostly by their names. However, for some accessions, their names do not indicate aromatic characters but they are truly aroma (Appa et al., 2006b). These accessions have not been well characterized for aromatic characters. The objective of this study was to characterize the variation in aromatic characters of Lao BGR by using chemical and molecular techniques. The results are very important in developing molecular-assisted breeding of aromatic rice using this germplasm.
MATERIALS AND METHODS
Chemical analysis: The research was undertaken in 2006 in the IRGC at the International Rice Research Institute (IRRI), the Philippines. A total number of 56 accessions were studied. They are 53 accessions of black glutinous rice randomly selected from Lao`s national genebank, two accessions of white glutinous rice with aromatic property, 6732 (Kai Noi Leuang) and 1655 (Hom Nang Nouan) and an improved white glutinous rice without aromatic character TDK5 (Table 1).
The experiment was laid out in a split plot design in completely randomized design with three replications (split plot in CRD). Main plots were polished and unpolished treatments and subplots were 56 rice genotypes. All the samples were dehulled, one set was polished using a grain test tube mill and the other set were left unpolished. Broken grains were removed by hand. Determination of 2-acetyl-1-pyrroline (2-AP) aromatic compound was carried out from these two sets. All samples were stored in 4C cold room to prevent volatilization of aromatic compounds until use. The detailed procedure for determining 2-AP concentration has been described previously (Bergman et al., 2000). Briefly, samples rice were ground in Sartorius Mikro-Dismembrator U grinder for 15 sec at 1800 rpm and screened in a 20 mesh grid. A 0.39 g of coarsely ground rice was loaded in 12x32 mm amber vials. 2, 4, 6-trimethylpyridine in dichloromethane was added and immediately covered with crimp tops.
| Table 1: | Fragrance and non-fragrance black glutinous rice varieties were identified chemical and molecular analysis |
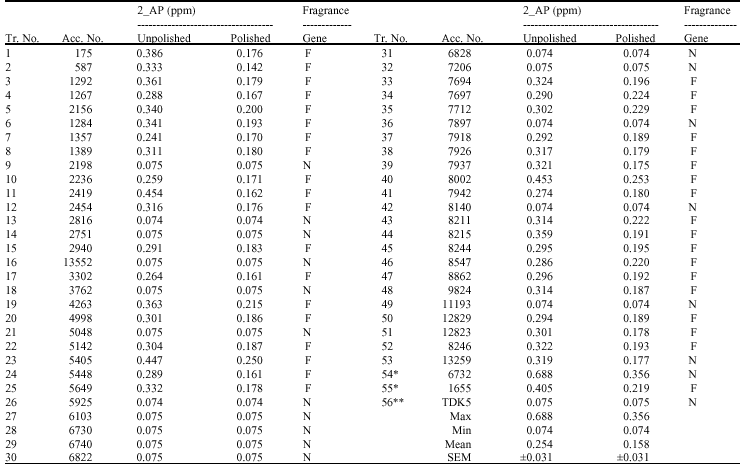 | |
| *: Indicates local aromatic white varieties Kai Noi Leuang (6732) and Hom Nang Nouan (1655); **: Indicates non-aromatic white improved variety Tr. No. = Treatment number; Acc. No. = Lao genebank accession number; F = homozygous fragrant, N = homozygous non-fragrant; SEM = standard error of the mean | |
Sample was heated at 85C in a water bath for 2.5 h. The sample was cooled and placed directly on an Agilent 6890N Gas Chromatograph (GC) equipped with flame ionization detector and SPB-5 column (35x250 μmx0.25 μm). GC parameters used were: inlet 155C (splitles), detector 300C, oven 35C held for 1.79 min, then increased to 70C at the rate 5C min-1, then to 100C at the rate of 10C min-1 and at 270C at the rate of 25C min-1 and held for 2 min. The concentration of 2-AP was quantified using a standard curve. Analysis of variance (ANOVA) and mean comparisons were performed using IRRISTAT version 4.02b (IRRI, 2000).
Molecular analysis
DNA extraction: Materials were those mentioned in the above section. KDML 105 and Calrose were also added as standard checks for fragrant and non-fragrant controls, respectively. Total genomic DNA was extracted from young leaves using a modification of the method described by Fulton et al. (1995). Approximately 5 mg of young leaf tissue was frozen in liquid nitrogen, followed by grinding using a plastic bit attached to an electric drill before the extraction buffer was added. The technical difference was that we used electric drill, while Fulton et al. (1995) used mortar and pestle. Other steps in extraction followed by Fulton et al. (1995). After extraction, pellets were dissolved in 150 μL TE (10 mM Tris, 1 mM EDTA, pH 8). DNAs were quantified by gel densitometry on agarose gels using Lambda DNA as a standard followed by normalization to the concentration of 5 ng μL-1 prior to use in PCR reaction.
PCR amplification: Typical PCR reaction mixture contained 2.5 mM MgCl2, 0.2 mM fgr primers, 1xBiomix and 10 ng DNA template. The details of primers used were shown in Table 2 (Bradbury et al., 2005b). Amplification was carried using Palm-Cycler (Corbett Life Science). The running conditions were 2 min at 94C, followed by 30 cycles of 30 sec at 94C, 30 sec at 55C, 35 sec at 72C, followed by a final extension period of 10 min at 72C and a hold at 15C until recovery.
PCR products were resolved by gel electrophoresis (Helixxtec iMupid) on 2% agarose. Each gel was photographed using Quantity One® 1-D analysis software, version 4.5.2 (Bio-Red, Hercules CA, USA) (reference), after SyberSafe (Invitrogen) gel staining. The software was used for imaging and analyzing one-dimensional electrophoretic gels, blots, dot blots and slot blots and for performing colony counts.
| Table 2: | Primers for analysis of fragrance in rice |
 | |
RESULTS AND DISCUSSION
Chemical analysis: Of 56 accessions, 39 genotypes contained high 2-AP, whereas the remaining 17 varieties contained only trace amounts of 2-AP (Table 1). The results were in agreement with those reported by Grimm et al. (2001) who could recover only trace amounts of 2-AP from non-aromatic rice. On average, the concentration of 0.254 ppm of 2-AP of unpolished sample was significantly higher than 0.158 ppm of polished sample. The results indicated that about 50% of 2-AP was lost after polishing. This finding was supported by the difference in peak area of each variety as shown by a sample chromatogram in Fig. 1. In this study, the difference between polished and unpolished for 2-AP was rather large, while other works reported the small difference (Bergman et al., 2000; Grimm et al., 2001), indicating uniform distribution of 2-AP in the grains. Nadaf et al. (2006) found higher 2-AP in unpolished samples than in polished samples. They pointed out that 2-AP did not uniformly distribute in the grains but located at higher concentration in the pericarps. Variation in 2-AP distribution might exist among a wide range of rice genotypes and could affect rice quality. Buttery et al. (1983) mentioned that the compound is released during cooking. However, the slow release of the compound might occur in storage or even in the field. The difference in the rates of the release might explain the differences among the results. It is interesting to note that polished and unpolished samples were not different for the samples with low 2-AP.
High variation in 2-AP concentration was found among all genotypes tested. The 2-AP concentrations varied from 0.074 to 0.688 ppm for unpolished samples and 0.074 to 0.356 ppm for polished samples (Table 1). For unpolished samples, the highest concentration of 0.688 ppm was found in the accession 6732 (Kai Noi Leuang), the local variety and the other local variety (accession 1655 or Hom Nang Nouan) had rather high concentration of 0.405 ppm that was similar to few black rice accessions with high 2-AP. For polished samples, the variation pattern was so similar except for lower 2-AP.
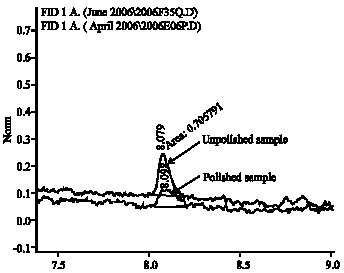 | |
| Fig. 1: | 2-AP chromatogram peaks of polished (low peak) and unpolished (high peak) sample of LG6103 rice samples |
Bergman et al. (2000) reported that 2-AP concentrations in rice ranged from 0.01 to 1.104 ppm, while Grimm et al. (2001) found that 2-AP concentrations could reach 1.256 ppm. Therefore, the levels of aromatic compound found in this germplasm were considered to be low to intermediate. Direct comparison of the results may not be appropriate because several factors can effect 2-AP concentrations in different experiments. However, it can give an idea about the extent of variation for this character that can be useful for breeding.
Molecular analysis: A recent study by Bradbury et al. (2005a, b) has discovered a 257 bp chromosomal fragment that links to fgr gene responsible for fragrance in rice and they have developed a protocol for screening this fragment. Bradbury et al. (2005a, b) proposed that a deletion in a gene homologous to Betaine Aldehyde Dehydrogenase 2 (BAD2) is the reason for fragrance in rice. The lowland black glutinous rice accessions were screened for presence or absence of the deletion using the method described by Bradbury et al. (2005b). Four fgr primers (ESP, IFAP, INSP and EAP) (Table 2) were used to amplify the genomic DNA of the 58 rice germplasm accessions. A primer combination of ESP and EAP could amplify a 580 bp fragment corresponding to the positive control in all 58 samples (Fig. 2). This band was used to indicate the success of the PCR amplification. A primer A 257 bp fragment produced by a primer pair of IFAP and ESP was identical to the fragrant diagnostic band.
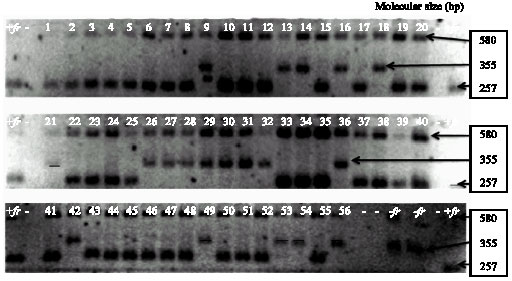 | |
| Fig. 2: | Three agarose gel 53 individuals from BGR varieties (1-53), three individual white rice varieties 54, 55 and 56 (Khao Kai Noi Leuang, Hom Nang Nouan and TDK5, respectively) and two standard varieties KDML 105 (+fr) and Calrose (-fr). The band of approximately 580 bp corresponds to the positive control amplified by both external primer (ESP and EAP). The 355 bp band corresponds to a PCR product amplified from the non-fragrant allele by the Internal Non-fragrant Sense Primer (INSP) and the External Anti-sense Primer (EAP). The 257 bp band corresponds to a PCR product amplified from the fragrant allele by the Internal Fragrant Anti-sense Primer (IFAP) and the External Sense Primer (ESP) pair of INSP and EAP produced a 355 bp fragment that occurred in 17 accessions of lowland black glutinous rice and two white glutinous rice varieties (6732 (or Kai Noi Leuang for local name) and TDK5) and the band was identical to that obtained in the non-fragrant standard variety, Calrose, indicating that they were non-fragrant. However, two genotypes (13259 and Kai Noi Leuang) with aromatic character (high 2-AP) were identified as non-fragrant by this fragment. Kai Noi Leuang has been well-known as aromatic local variety widely grown throughout the country. The contrasting results between 2-AP analysis and molecular analysis are difficult to explain. Different gene or chromosomal location might be responsible for the observed fragrance in 13259 and Kai Noi Leuang, or there would be the presence of another major aromatic compound analogous to 2-AP. Further investigations are required to elucidate this question. |
This fragment was found in the remaining 36 accessions of BGR and a local variety (1655 or Hom Nang Neuan). These genotypes shared a common fragment to that of the standard fragrant variety KDML 105. All of the accessions tested were either homozygous fragrant or homozygous non-fragrant (Table 1). The results confirmed the previous findings of Bradbury et al. (2005a) who demonstrated that the flavour or fragrance of basmati or jasmine rice is associated with the presence of 2-AP. A recessive gene (fgr) on chromosome 8 of rice has been linked to this important trait. Fragrance in domesticated rice has apparently originated from a common ancestor and may have evolved in a genetically isolated population, or may be the outcome of a separate domestication event. This is an example of effective human selection for a recessive trait during domestication. However, although 2-AP was found in two accessions (13259 and Kai Noi Leuang), the 257 bp fragment was not present in these accessions. This might be the second event of domestication of aromatic rice.
In conclusion, significant variation in 2-AP associated with fragrance in rice was found in the lowland black glutinous rice varieties of the Lao PDR. Most accessions with high 2-AP had a 257 bp fragment in common except for 13259 and Kai Noi Leuang. From our evidence, single event of domestication of fragrance in rice is in question and further investigations are required.
ACKNOWLEDGMENTS
We thank Mr. Phoumi INTHAPANYA Director of Agricultural Research Center of Lao PDR, for seed samples of Lao accessions. Financial support from IRRI is gratefully acknowledged. Grateful acknowledgment has been made to Mr. Thawan KESMALA for his critical editing during manuscript preparation. Special thanks to M. NAVARRO and V. BUTARDO Jr. for their good assist me during I conducted my research at IRRI (Grain Quality, Nutrition and Postharvest Centre).
REFERENCES
- Appa, R.S., C. Bounphanousay, J.M. Schiller and M.T. Jackson, 2002. Collection, classification and conservation of cultivated and wild rices of the Lao PDR. Genet. Res. Crop Evol., 49: 75-81.
CrossRef - Appa, R.S., C. Bounphanousay, J.M. Schiller, A.P. Alcantara and M.T. Jackson, 2002. Naming of traditional rice varieties by farmers in the Lao PDR. Genet. Res. Crop Evol., 49: 83-88.
CrossRef - Bergman, C.J., J.T. Delgado, R. Bryant, C. Grimm and K.R. Cadwallader et al., 2000. Rapid gas chromatographic technique for quantifying 2-acetyl-1-pyrroline and hexanal in rice (Oryza sativa L.). Cereal Chem., 77: 454-458.
CrossRef - Bradbury, L.M.T., R.J. Henry, Q.S. Jin, R.F. Reinke and D.L.E. Waters, 2005. A perfect marker for fragrance genotyping in rice. Mol. Breed., 16: 279-283.
CrossRefDirect Link - Buttery, R.G., L.C. Ling, B.O. Juliano and J.G. Turnbaugh, 1983. Cooked rice aroma and 2-acetyl-1-pyrroline. J. Agric. Food Chem., 31: 823-826.
CrossRefDirect Link - Buttery, R.G., J.G. Turnbaugh and L.C. Ling, 1988. Contribution of volatiles to rice aroma. J. Agric. Food Chem., 36: 1006-1009.
CrossRefDirect Link - Chen, S., J. Wu, Y. Yang, W. Shi and M. Xu, 2006. The fgr gene responsible for rice fragrance was restricted within 69 kb. Plant Sci., 171: 505-514.
CrossRefDirect Link - Cordeiro, G.M., M.J. Christorpher, R.J. Henry and R.F. Reinke, 2002. Identification of microsatellite markers for fragrance in rice by analysis of the rice genome sequence. Mol. Breed., 9: 245-250.
CrossRef - Fulton, T.M., J. Chunwongse and S.D. Tanksley, 1995. Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep., 13: 207-209.
CrossRefDirect Link - Goodwin, H.L., M.E. Rister, L.L. Koop, A.M. McClung and R.K. Miller et al., 1994. Impacts of various cultural, harvest and postharvest handling practices on rice quality attributes of Jasmine 85. Proceedings of the 26th Rice Tech. Work Group. Texas Agric. Exp. Stn., March 6-9, 1994, Texas A and M University, College Station, TX., pp: 124-125.
- Grimm, C.C., C. Bergman, J.T. Delgado and R. Bryant, 2001. Screening for 2-acetyl-1-pyrroline in the headspace of rice using SPME/GC-MS. J. Agric. Food Chem., 49: 245-249.
CrossRef - Jain, N., S. Jain, N. Saini and R.K. Jain, 2006. SSR analysis of chromosome 8 regions associated with aromaand cooked kernel elongation in Basmati rice. Euphytica, 152: 259-273.
CrossRef - Jin, Q., D. Waters, G.M. Cordeiro, R.J. Henry and R.F. Reinke, 2003. A Single Nucleotide Polymorphism (SNP) marker linked to the fragrance gene in rice (Oryza sativa L.). Plant Sci., 165: 359-364.
CrossRefDirect Link - Khush, G.S., 1997. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol., 35: 25-34.
CrossRef - Murray, S.S., 2004. Searching for the origins of African rice domestication. Antiquity, Vol. 78, No. 300.
Direct Link - Nadaf, A.B., S. Krimshnan and K.V. Wakte, 2006. Histochemical and biochemical analysis of major aroma compound (2-acetyl-1-pyrroline) in basmati and other scented rice (Oryza sativa L.). Curr. Sci., 91: 1533-1536.
Direct Link - Pinson, S.R.M., 1994. Inheritance of aroma in six rice cultivars. Crop Sci., 34: 1151-1157.
CrossRefDirect Link - Widjaja, R., J.D. Craske and M. Wootton, 1996. Comparative studies on volatile components of non-fragrant and fragrant rice. J. Sci. Food Agric., 70: 151-161.
CrossRefDirect Link - Zhang, W. and J. Yuan, 1998. A preliminary study of ancient excavated rice from Yuchanyan site, Dao County, Hunan province, PR China. Acta Agron. Sin., 24: 416-420.
Direct Link - Bradbury, L.M.T., T.L. Fitzgerald, R.J. Henry, Q. Jin and D.L.E. Waters, 2005. The gene for fragrance genotyping in rice. Plant Biotechnol. J., 3: 363-370.
CrossRefDirect Link