
Wen Teng
Jiangsu Key Laboratory of Environmental Change and Ecological Construction, College of Geographical Science, Nanjing Normal University, Nanjing 210097, China
Wang Guoxiang
Jiangsu Key Laboratory of Environmental Change and Ecological Construction, College of Geographical Science, Nanjing Normal University, Nanjing 210097, China
Li Qiang
Jiangsu Key Laboratory of Environmental Change and Ecological Construction, College of Geographical Science, Nanjing Normal University, Nanjing 210097, China
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 5 | Page No.: 773-780







ABSTRACT
Mature Myriophyllum spicatum L. were planted in ponds with different turbidities (30 NTU, 60 NTU, 90 NTU), which were made by sand particles less than 100 μm. After two months growth, measurements of chlorophyll parameters were made on leaves near stems by a pulse-amplitude fluorometer (Diving-PAM) to investigate the effects of water turbidity on the photosynthetic characteristics of Myriophyllum spicatum L. The Fv/Fm and F0 of Myriophyllum spicatum L. in three turbid ponds were not significantly affected by water turbidity. The qP and ΔFv/Fm’ of plants in pond B and C (30 and 60 NTU) were not significantly decreased when compared with control, while in pond D (90 NTU) they were apparently reduced (p<0.05). Moreover, the values of qN in all four ponds were rather low (≤ 0.1). This indicates that water turbidity can’t obviously affect the function of PSII of Myriophyllum spicatum L. and most light energy are still used to drive photosynthesis while less are dissipated as heat. The diurnal variation extents of rETR in three turbid ponds were significantly smaller than that in control, indicating that the photosynthesis of plants in turbid ponds was significantly decreased with increasing water turbidity. The diurnal variation of Fv/Fm exhibited a gentle V-shape, which indicates Myriophyllum spicatum L. could endure higher irradiance. In ponds whose turbidity were higher than 60 NTU, the recovering speed of Fv/Fm was slower than that in control, but the difference wasn’t significant, which demonstrates that water turbidity showed some impact on the anti-photoinhibition ability and recovery capacity of Myriophyllum spicatum L., but the impact was not significant. The value of rETRmax was decreased with increasing water turbidity, showing that photosynthesis capacity of the plants was apparently decreased.
PDF Abstract XML References Citation
How to cite this article
Wen Teng, Wang Guoxiang and Li Qiang, 2007. Effects of Water Turbidity on the Photosynthetic Characteristics of Myriophyllum spicatum L. Asian Journal of Plant Sciences, 6: 773-780.
DOI: 10.3923/ajps.2007.773.780
URL: https://scialert.net/abstract/?doi=ajps.2007.773.780
DOI: 10.3923/ajps.2007.773.780
URL: https://scialert.net/abstract/?doi=ajps.2007.773.780
INTRODUCTION
Various suspended components of water turbidity can affect the productivity of submerged macrophytes (Korschgen et al., 1997). Suspended substances in water can physically block the penetration of irradiance through the water column and greatly lower light availability to submerged plants and inhibit their growth (You and Song, 1995; Noriko et al., 2003). Zhang et al. (2006) have found that suspended substances is one of the important factors controlling the reconstruction of submerged macraphytes. However, submerged macrophytes play an important role in forming a clear macrophyte-dominated state in shallow lakes instead of a turbid phytoplankton-dominated state (Scheffer, 1998) by several mechanisms, such as competing with algae for nutrients, reducing the resuspension of the sediment or providing refuge for zooplankton (Torben et al., 2003; Mariana et al., 2003). Therefore submerged macrophytes has been widely used in lake restoration research. Generally, the suspended substances in turbid water could be divided into two major groups: one is organic matter and algae debris which usually exist in the eutrophic lake; the other is inorganic suspended solids like sands which appear in some lakes connected to rivers. Recent research always focuses on the former one, while the latter one has not so far been studied, especially its effect on submerged macrophytes.
When investigating the effect of light intensity on submerged macrophytes, plant morphology and its biomass are two major values (Korschgen et al., 1997), although iodimetric analysis (Su et al., 2004) and oxygen measurement system (Ben et al., 2002) are also introduced in some articles. However, all these methods only can be used on the detached leaves and can’t directly and truly reflect the photosynthetic process of submerged macrophytes. In the recent years, the introduction of a number of highly user-friendly (and portable) chlorophyll fluorometer can well solve this problem. The DIVING-PAM Underwater Fluorometer is one of them, which has been widely used for studying in situ photosynthesis in underwater plants under natural conditions (Beer et al., 1998). The technique is rapid, extremely sensitive and non-intrusive and can be performed on intact, attached leaves (Anthony and Christa, 1999). The operation of the tool is very easy and valuable information may be obtained within seconds. For these reasons Diving-PAM fluorometer has become an important tool in study of photosynthesis, in particular the functioning of PSII (Schreiber et al., 1995).
In China, some lakes and streams connected with rivers have lost submerged vegetation and species diversity. There are only some emerging macrophytes like reeds and Zizania latifolia growing along the bank. The major reason is high water turbidity lowers light availability to submerged plants and reduces their growth. Therefore, the essential step to restore and revegetate the submerged plant is investigating the effect of water turbidity on its growth. Myriophyllum spicatum L., is a submerged aquatic herb with branching leafy shoots and finely dissected whorls of leaves (Aiken et al. 1979). It is popularly used in lake revegetation programs (Zhu et al., 2002; Keskinkan et al., 2003; Lauridsen et al., 2003; Sivaci et al., 2004) not only because it can grow well in eutrophicate lakes, but also because it is effective in reducing eutrophication (especially for N and P; Song et al., 1997; Zhou et al., 2000) and inhibiting blue-green algae (Nakai et al., 2000; He et al., 2002). However, its growth in turbid water rich in suspended solids has not been well studied. In this present study, the aim was to investigate the effect of water turbidity on the photosynthetic characteristics of Myriophyllum spicatum L. by the tool of Diving-PAM.
MATERIALS AND METHODS
Plant material: In this study, the plants of Myriophyllum spicatum L. were collected from ponds along Yangtse River located in the Liuhe area of Nanjing city. Twenty days before the experiment, plants in similar development level were selected and planted in the experimental ponds. The sediment in the ponds was 0.3 m in depth.
Experiments design: All experiments were conducted from July 11, 2005 to September 14, 2005 in four outdoor enclosure ponds, located in Jiangsu Key Lab of Environmental Change and Ecological Construction. All ponds were 9.4 m in length and 1.1 m in width. The depths of water were 1.3 m. According to the water turbidity in the lower region of Yangtse River, the turbidities in 3 ponds (B, C and D) were respectively adjusted to 30, 60 and 90 NTU (Nephelometric Turbidity Units), while one pond was left as control (A). During the experiment, a re-circulation system was implemented among 4 ponds by pumps and the velocity of water was set about 1.5 mm sec-1. Every pond was evenly settled into 50 plants of Myriophyllum spicatum L. Before the experiment started, the average height of plants is about 1.0 m. Two months later, 10 mature plants of Myriophyllum spicatum L. in every pond were randomly selected and measurements of fluorescence parameters were made.
Sands preparation: Sands used in the study were totally collected from lower region of Yangtse River, located in city Zhenjiang of Jingsu. Then sands were solved in water and filtered by an iron grid (mesh diameter 100 μm). After 2 h, the upper part of filtrate, the suspended sands used in our study, were isolated and analyzed. The turbidity of water was measured with a HACH 2100P Portable turbidimeter.
Chlorophyll fluorescence measurements: All chlorophyll fluorescence measurements were performed using a Diving-PAM (PAM 2000: H. Waltz GmbH, Effeltrich, Germany) and all data were collected using a WinControl program (Waltz GmbH, Effeltrich, Germany). Ten mature plants of Myriophyllum spicatum L. in each pond were randomly selected and measurements were made on upper (adaxial) surface of leaves, which had been pre-darkened for 10 min. All data were reported as mean ± SE. Then the data from three turbid ponds were respectively compared with the one from control. A software of SPSS 11.5 was used to examine significant difference among them and the criterion of significance was set at p<0.05. Measurements included: basic fluorescence upon exposure to weak light after dark adaptation (F0), basic fluorescence after light adaptation (F0’), maximum fluorescence (Fm), maximum fluorescence yield of a light adapted leaf exposed to a pulse of saturating light (Fm’) and steady state fluorescence of a light adapted leaf (Ft). These measurements were used to determine quenching and electron transport rate. The potential maximal efficiency of PSII (Fv/Fm) of dark adapted leaves was calculated as Fv/Fm = (Fm-Fo)/Fm. The effective quantum Yield of PSII (ΔFv/Fm’) was calculated as Yield (ΔFv/Fm’) = (Fm’-Ft)/Fm’ (Schreiber et al., 1997). Photochemical quenching (qP) was calculated as qP = (Fm’-Ft)/(Fm’-F0’). Nonphotochemical quenching (qN), which refers to the nonradiative dissipation of energy, was calculated as qN = (Fm-Fm’)/(Fm-F0). Relative electron transport rate (rETR) was calculated as rETR = YieldxPARx0.84x0.5 (Schreiber et al., 1997). The whole process of chlorophyll fluorescence measurements were carried out as described previously (Kate and Giles, 2000) from 7:00 to 8:00 am. Initially, a measuring light (0.15 μmol photon m-2 sec-1) was switched on, giving a measure of the F0 (minimal) level of fluorescence. A 0.8 sec saturating-pulse light (4000 μmol photon m-2 sec-1) was then applied, allowing the measurement of Fm. Following on this, a 10 sec actinic light (30 μmol photo m-2 sec-1) was applied and at appropriate interval, further saturating light was applied. From each of these, Fm’ and Ft, can be measured.
Diurnal variation of photosynthetic rate measurements: The experiment was carried out every two hours during the day (from 7:00 am to 17:00 pm). Ten mature plants of Myriophyllum spicatum L. were randomly selected and the measurements were performed by the Diving-PAM (PAM 2000: H. Waltz GmbH, Effeltrich, Germany). Initially, the measures of Fo and Fm were carried out as described previously. Then a 0.8 sec saturating-pulse light (4000 μmol photon m-2 sec-1) was applied to give measurements of Fm’ and Ft. Upon this, the value of rETR and Fv/Fm could be calculated.
Rapid Light Curving (RLC) measurement: 10 mature plants of Myriophyllum spicatum L. were randomly selected in every pond and measurements of RLCs were performed by the Diving-PAM (PAM 2000: H. Waltz GmbH, Effeltrich, Germany). The whole process of RLCs measurements were carried out as described previously from 7:00 to 8:00 am. Initially, the leaves were pre-darkened for 10 sec. Then the leaf clip was removed and a measuring light (0.15 μmol photon m-2 sec-1) was switched on, giving a measure of the F0 (minimal) level of fluorescence. A 0.8 sec saturating-pulse light (4000 μmol photon m-2 sec-1) was then applied, allowing the measurement of Fm. Following on this, a series of 10 sec actinic lights (17, 49, 104, 176, 248, 342, 506, 684 μmol photo m-2 sec-1) was applied and at appropriate interval, further saturating light was applied. From each of these, Fm’ and Ft, can be measured. According to ETR = YieldxPARx0.84x0.5, weight mean values of ETR were obtained and then the RLCs could be visualized (Peter and Rolf, 2005).
RESULTS
Response of Fv/Fm and Fo to increasing water turbidity: Fv/Fm is a parameter, which measures the intrinsic (or maximum) efficiency of PSII (i.e., the quantum efficiency if PSII centres were essentially all open). Dark-adapted values of Fv/Fm reflect the potential quantum efficiency of PSII and are used as a sensitive indicator of plant photosynthetic performance (Björkman and Demmig, 1987; Johnson et al., 1993). After dark-adaptation, the values of Fv/Fm leaves in pond B, C and D (30, 60 and 90 NTU) were measured by Diving-PAM. They were respectively decreased by 99.95, 97.69 and 97.23% compared with control (Fig. 1), but the decreases were not significant (p>0.05).
Fo, the yield of fluorescence in the absence of an actinic (photosynthetic) light when PSII reaction centres are totally open, gives information about the efficiency of photochemical quenching and by extension, the performance of PSII. The elevated level of non-photochemical energy dissipation within the antennae matrix can possibly result in the decrease of F0, while the photodamage and reversible inactivation to PSII reaction center can cause the increase of F0. The value of F0 in pond B, C and D (30, 60 and 90) NTU, were respectively increased by 11.3, 15.8 and 21.0%, when compared with the control (Fig. 1), but the increases were not significant (p>0.05).
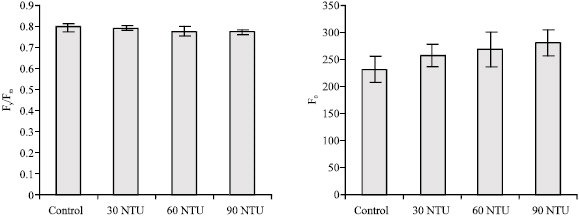 | |
| Fig. 1: | Fv/Fm and F0 of Myriophyllum spicatum L. in different turbidity ponds. Values are mean ± SE (n = 10), n is number of plants |
Response of ΔFv/Fm’, qP, qN and ETR to increasing water turbidity: The photosynthetic performance of a plant under light exposing can be reflected by some fluorescence parameters, such as effective quantum Yield of PSII (ΔFv/Fm’), photochemical quenching (qP), non-photochemical quenching (qN) and relative electron transport rate (rETR). Since the light intensity is greatly diminished by the suspended solids in turbid water, the real irradiance the underwater leaf can get is usually lower than the irradiance on the surface water. Therefore, because the surface water irradiance at 7:00 am was about 30 μmol photo m-2 sec-1, the leaf of Myriophyllum spicatum L. was exposed to the actinic light (30 μmol photo m-2 sec-1) for 10 sec in our experiment and then the measurements of fluorescence parameters were made.
ΔFv/Fm’, this parameter measures the photochemical efficiency of PS II, which refers to the proportion of light absorbed by chlorophyll associated with PS II that is used in photochemistry. Compared with the control pond, the values of ΔFv/Fm’ in pond B, C and D (30, 60 and 90 NTU) were respectively decreased by 0.3, 3.5 and 4.1% (Fig. 2A), but the decreases were not significant (p>0.05).
qP, the parameter of photochemical quenching, gives an indication of the proportion of PSII reaction centers that are open (Kate and Giles, 2000). A decrease in qP is due to closure of reaction centers and less quantum yield of carbon fixation, resulting from a saturation of photosynthesis by light. The values of qP in pond B, C and D (30, 60 and 90 NTU) were reduced by 3.0, 4.1 and 5.3% when compared with control (Fig. 2B). The decline achieved significant level in pond D (90 NTU) (p<0.05).
qN, linearly related to heat dissipation, can measure changes in heat dissipation. Any change in qN gives information about change in the efficiency of heat dissipation, which occur as a result either of processes that protect the leaf from light-induced damage or of the damage itself (Kate and Giles, 2000). In all four ponds, although the values of qN were increased with increasing water turbidity (control<30<60<90 NTU), they were all lower than 0.1, which implies that all plants showed limited development of non-photochemical quenching (Fig. 2C).
The Electron Transport Rate (ETR) was found to be closely related to the photosynthetic activity when measured by oxygen evolution or CO2 uptake (Beer et al., 1998), which is expressed as μmols of electrons per m-2 s-1. Relative ETR is an approximation of the rate of electrons pumped through the photosynthetic chain (Beer et al., 2001), which is calculated as rETR = YieldxPARx0.84x0.5 (Schreiber et al., 1997).
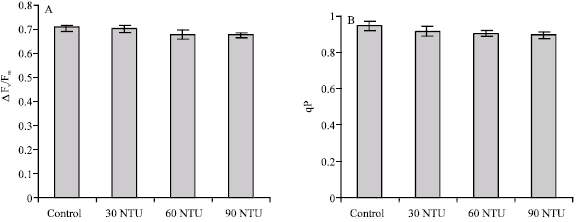 | |
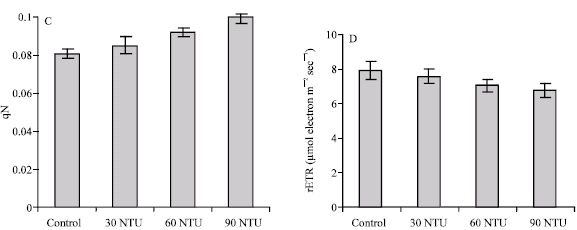 | |
| Fig. 2: | Yield, qN, qP and rETR of Myriophyllum spicatum L. in different turbidity ponds. Values are mean ± SE (n = 10), n is number of plants |
In the experiment, the values of rETR in turbid ponds were all lower than the one in control. Statistical analysis demonstrates that water turbidity had significant effect on the plants in pond D (90 NTU) (p<0.05), but no significant effect on the other two turbid ponds ( 30 and 60 NTU) was observed (Fig. 2D).
Response of diurnal variation of rETR and Fv/Fm to increasing water turbidity: To further investigate the effect of water turbidity on the PSII properties of Myriophyllum spicatum L., the diurnal variation of rETR and Fv/Fm in all four ponds were determined with Diving-PAM. The diurnal variation of irradiance, rETR and Fv/Fm were plotted as dotted curves. At 7:00 am, the irradiance on the surface water gradually increased, reached an elevated level at 11:00 am, achieved a zenith at 13:00 pm and then gradually decreased (Fig. 3). The curves of diurnal rETR variation exhibited similar shapes to the curves of diurnal irradiance variation. During the period from 9:00 am to 15:00 pm, the values of rETR were decreased with the increasing water turbidity (control>30> 60>90 NTU) and the differences of rETR between the three turbid ponds and the control pond were significant (p<0.05).
In contrast, the diurnal Fv/Fm variation exhibited flat V-shaped curves, which were almost opposite shapes to the curves of diurnal irradiance variation (Fig. 4). The diurnal variation of Fv/Fm in pond B (30 NTU) was not significant different with the one in control pond (p>0.05). Similar to control, its lowest value of Fv/Fm occurred at 13:00 pm, but at 17:00 pm its Fv/Fm could recover to 96.5% of the one at 7:00 am, showing a higher capacity for Fv/Fm recovery. In pond C and D (60 and 90 NTU), the differences between diurnal Fv/Fm variation and the control were enlarged when compared with pond B (30 NTU), but still not significant (p>0.05). Moreover, the recovery of Fv/Fm in the two turbid ponds was slower than that in pond B (30 NTU), that is, at 17:00 pm the value of Fv/Fm respectively recovered to 95.0 and 92.9% when compared with their own initial value at 7:00 am.
Response of Rapid Light Curves (RLCs) to increasing water turbidity: Light curves is another useful parameter to measure and describe the acclimation of the photosynthetic apparatus PSII to a range of water turbidity. In comparison to traditional light curves, a Rapid Light Curve (RLC) measures the effective quantum yield as a function of irradiance. It can provide a reliable assessment of photosynthetic activity, by integrating the leaf’s ability to tolerate light fluctuation, as well as reflecting its immediate short-term light history (Schreiber et al., 1997).
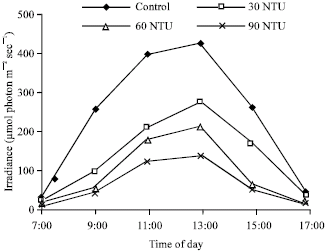 | |
| Fig. 3: | Diurnal variation of irradiance in different turbidity ponds |
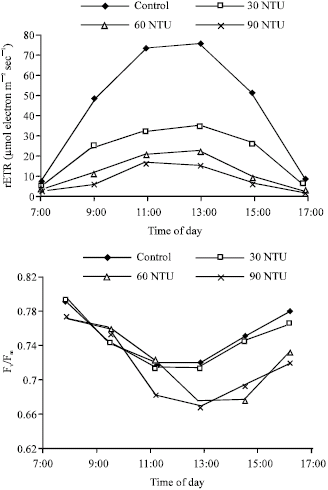 | |
| Fig. 4: | Diurnal variation of rETR and Fv/Fm of Myriophyllum spicatum L. in different turbidity ponds. Values are mean ± SE (n = 10), n is number of plants |
Figure 5 showed four average RLCs from four experimental ponds. Increasing water turbidity can not affect the minimum saturation irradiance (Ek) (Fig. 5).
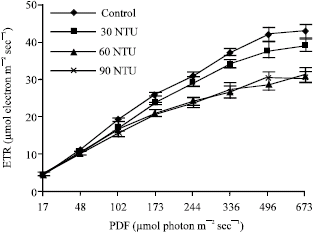 | |
| Fig. 5: | Rapid-light curves (RLCs) of Myriophyllum spicatum L. in different turbidity ponds. Values are mean ± SE (n = 10), n is number of plants |
In four ponds (from A to D), the values of Ek were all 673 μmol photon m-2 sec-1. The value of rETRmax, however, was affected by water turbidity. In control pond, the value of rETRmax was 42.08 μmol electron m-2 sec-1, significantly higher than those in three turbid ponds (30, 60 and 90 NTU), whose rETRmax were 37.71 μmol electron m-2 sec-1, 28.75 μmol electron m-2 sec-1 and 30.51 μmol electron m-2 sec-1, respectively. And with the increase of irradiance, the values of rETR in three turbid ponds became more significant different with control. Additionally, the difference of rETR between pond C and D (60 and 90 NTU) was not apparent, but the values of rETR in the two ponds were profoundly lower than that in pond B (30 NTU).
DISCUSSION
Water turbidity caused by suspended particulates not only can physically block the penetration of irradiance through the water column, but also may be harmful to submerged macrophytes when deposited on leaf surfaces by reducing light transmission to photosynthetically active leaf surfaces and possibly altering gas and nutrient exchange (Korschgen et al., 1997), which finally would lead to the decrease of photosynthetic capability and the damage to photosynthetic apparatus. After planted in different turbidity ponds for two-month, there were many suspended sands deposited on the leaf of Myriophyllum spicatum L. However, present study revealed that water turbidity may have caused a degree of photodamage to the leaf of Myriophyllum spicatum L., but can’t apparently affect its photosynthetic apparatus-PSII, because the F0 and Fv/Fm of dark-adapted leaf in the three turbid ponds didn’t show significant difference with control.
Over the past 17 years, the measurement of the chlorophyll a fluorescence has proven to be a powerful method of assessing the properties of the PSII which is the most sensitive component of photosynthetic apparatus within intact leaves (Schreiber, 2004). Light energy absorbed by chlorophyll molecules in a leaf can undergo one of three fates: it can be used to drive photosynthesis (photochemistry), excess energy can be dissipated as heat or it can be re-emitted as light-chlorophyll fluorescence (Kate and Giles, 2000). Yet, the total amount of chlorophyll fluorescence is very small (only 1 or 2% of total light absorbed) and most energy are used in photochemistry process and heat dissipation. The effective quantum yield of PSII (ΔFv/Fm’), photochemical quenching (qP) and non-photochemical quenching parameters (qN and NPQ) are some coefficients, describing the relative influence of the energy dissipation pathways. qP and ΔFv/Fm’ can give a measure of photochemistry, while qN and NPQ are both measures of the energy flow into heat (Peter and Rolf, 2005). An increase of qP and ΔFv/Fm’ indicates more light energy are used in photochemical process, while an increase of qN and NPQ indicates more energy are dissipated as heat. In our study, the fluorescence parameters of Myriophyllum spicatum L. were measured under a 30 μmol photo m-2 sec-1 actinic illumination. Results showed that in pond B and C (30 and 60 NTU), the value of qP, ΔFv/Fm’ and ETR were not significantly decreased, but the values of qN were rather low. This indicates that water turbidity can’t obviously affect the function of PSII of Myriophyllum spicatum L. and most light energy are still used to drive photosynthesis while less are dissipated as heat.
Generally, the photosynthetic process of plant is closely related with the diurnal variation of irradiance. In turbid water rich in suspended sands, the irradiance under water is clearly decreased with increasing water turbidities. In the experiment, the diurnal variation curves of rETR showed that the value of rETR in all four ponds were initially increased with the increasing irradiance, reached a maximum at noon and then gradually decreased. However, compared with control, the values of rETR in three turbid ponds were significantly decreased and were decreased with increasing water turbidity. This indicates that water turbidity showed significant effect on the photosynthetic process of Myriophyllum spicatum L. and the diurnal photosynthetic capacity of plants was decreased with increasing water turbidities. In contrast, the diurnal variation of Fv/Fm displayed almost opposite behavior to the diurnal variation of rETR. But the slope of the V-shape diurnal Fv/Fm variation curves was quite gentle, indicating that although high irradiance did induce photoinhibition on the leaves of Myriophyllum spicatum L., but the plant’s tolerant ability and recovery capacity to photoinhibition were not significant affected.
Rapid light curves (RLCs) can provide detailed information on the saturation characteristics of electron transport, as well as the overall photosynthetic performance of a plant. Results of RLCs demonstrate that under high irradiance (higher than 100 μmol photo m-2 sec-1), the relative electron transport rate (rETR) of Myriophyllum spicatum L., including the value of rETRmax was significantly decreased with increasing water turbidity, while its minimum saturation irradiance (Ek) was not affected. This implies that water turbidity can profoundly decreased the photosynthetic acclimation ability of Myriophyllum spicatum L. to light fluctuation and under higher irradiance the effect became more apparently with increasing water turbidity. However, its capacity to resist photoinhibition was not significantly affected.
In conclusion, results presented here show that mature Myriophyllum spicatum L. is a submerged species which could tolerate high water turbidity caused by suspended solids. It can grow well in turbid water whose turbidity is lower than 30 NTU (including 30 NTU). However, growing in water whose turbidity is higher than 60 NTU (including 60 NTU), its whole photosynthesis is dramatically reduced with increasing water turbidity, but no significant impact on its capacity to resist photoinhibition is observed under high irradiance. Therefore, Myriophyllum spicatum L. is a good aquatic species when applying revegetation and restoration in some lakes or streams connected with rivers. Obviously, further studies are needed to demonstrate the effect of water turbidity on the growth and development of Myriophyllum spicatum L. at a larger time and special scale.
REFERENCES
- Anthony, J.W. and C. Christa, 1999. Rapid light curves: A new fluorescence method to assess the state of the photosynthetic apparatus. Photosynthetic Res., 59: 63-72.
Direct Link - Beer, S., M. Bjork, R. Gademann and P.J. Ralph, 2001. Measurement of Photosynthesis in Seagrasses. In: Global Seagrass Research Methods, Short, F.T. and R. Coles (Eds.), Elsevier Publishers, The Netherlands, pp: 183-198.
Direct Link - Ben, J.L., K. Tim and W.R. John et al., 2002. An in situ study of photosynthetic oxygen exchange and electron transport rate in the marine macroalga Ulva lactuca (Chlorophyta). Photosynthesis Res., 74: 281-293.
PubMedDirect Link - Bjorkman, O. and B. Demmig, 1987. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta, 170: 489-504.
CrossRefDirect Link - Johnson, G.N., A.J. Young, J.D. Scholes and P. Horton, 1993. The dissipation of excess excitation energy in British plant species. Plant Cell Environ., 16: 673-679.
CrossRef - Maxwell, K. and G.N. Johnson, 2000. Chlorophyll fluorescence-A practical guide. J. Exp. Bot., 51: 659-668.
Direct Link - Keskinkan, O., M.Z.L. Goksu, A. Yuceer, M. Basibuyuk and C.F. Forster, 2003. Heavy metal adsorption characteristics of a submerged aquatic plant (Myriophyllum spicatum). Process Biochem., 39: 179-183.
CrossRefDirect Link - Korschgen, C.E., W.L. Green and K.P. Kenow, 1997. Effects of irradiance on growth and winter bud production by Vallisneria americana and consequences to its abundance and distribution. Aqua. Bot., 58: 1-9.
CrossRef - Lauridsen, T.L., H. Sandsten and P.H. Moller, 2003. The Restoration of a shallow lake by introducing Potamogeton spp. : The impact of waterfowl grazing. Lakes Reservoirs Res. Manage., 8: 177-187.
CrossRefDirect Link - Mariana, M., M. Nestor, M. Brian and R. Lorena, 2003. The structuring role of free-floating versus submerged plants in a subtropical shallow lake. Aqua. Ecol., 37: 377-391.
CrossRefDirect Link - Nakai, S., Y. Inoue, M. Osomi and A. Murakami, 2000. Myriophyllum spicatum released allelopathic polyphenols inhibiting growth of blue-green algae Microcystis aeruginosa. Water Res., 34: 3026-3032.
CrossRefDirect Link - Noriko, T., K. Yasuro, F. Michio, N. Megumi and B.H.O. Kim, 2003. Effects of aquatic macrophytes on water quality and phytoplankton communities in shallow lakes. Ecol. Res., 18: 381-395.
CrossRefDirect Link - Peter, J.R. and G. Rolf, 2005. Rapid light curves: A powerful tool to assess photosynthetic activity. Aqua. Bot., 82: 222-237.
Direct Link - Schreiber, U., H. Hormann, C. Neubauer and C. Klughammer, 1995. Assessment of photosystem II photochemical quantum yield by chlorophyll fluorescence quenching analysis. Aust. J. Plant Physiol., 22: 209-220.
CrossRefDirect Link - Schreiber, U., R. Gademann, P.J. Ralph and A.W.D. Larkum, 1997. Assessment of photosynthetic performance of prochloron in Lissoclinum patella in hospite by chlorophyll fluorescence measurements. Plant Cell Physiol. Tokyo, 38: 945-951.
Direct Link - Schreiber, U., 2004. PAM Pulse-amplitude Fluorometry and Saturation Pulse Method. In: Chlorophyll Fluorescence: A Signature of Photosynthesis, Advances in Photosynthesis and Respiration Series, Papageorgiou, G. and Govindjee (Eds.). Kluwer Academic Publishers, Dordrecht, The Netherlands, pp: 279-319.
- Sivaci, E.R., A. Sivaci and M. Sokmen, 2004. Biosorption of Cadimium by Myriophyllum spicatum L. and Myriophyllum triphyllum orchard. Chemosphere, 56: 1043-1048.
Direct Link - Su, W.H., G.F. Zhang, Y.S. Zhang, Y.S. Xiao and H.F. Xia et al., 2004. The photosynthetic characteristics of five submerged aquatic plants. Acta Hydrobiol. Sin., 28: 391-395.
Direct Link - Torben L.L., P.J. Jens J. Erik and S. Martin, 2003. Response of submerged macrophytes in danish lakes to nutrient loading reductions and biomanipulation. Hydrobiologia, 506-509: 641-649.
CrossRefDirect Link - Zhou, Y., J. Li and Y. Fu, 2000. Effects of submerged macrophytes on kinetics of alkaline phosphatase in lake donghu-I. Unfiltered water and sediments. Water Res., 34: 3737-3742.
Direct Link