
M. Motalebi
Department of Agriculture Science and Research Branch, Islamic Azad University, Tehran, Iran
M. Keshavarzi
Seed and Plant Improvement Institute, Karaj, Iran
M.R. Naghavi
Departmaent of Plant Breeding, Agricultural College, The University of Tehran, Karaj, Iran
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 399-402







ABSTRACT
Twenty-five accessions of Triticum durum originating from different geographical areas of Iran and 10 durum cultivars from European countries were evaluated for high molecular weight glutenin subunit (HMW-GS) composition using SDS-PAGE. The data indicated the prevalence of the null allele (52%) and 2* subunit (48%) at the Glu-A1 and four alleles, namely 14+15 (32%), 13+16 (28%) and 7+20 (24%) represented about 80% of the alleles at Glu-B1 locus in the Iranian durum compared to European durum. This information can be a valuable reference for designing breeding program for the improvement of breed and pasta making quality of bread and durum wheat, respectively in Iran.
PDF Abstract XML References Citation
How to cite this article
M. Motalebi, M. Keshavarzi and M.R. Naghavi, 2007. Glutenin Subunit Composition in Durum (Triticum durum) Landraces and Cultivars. Asian Journal of Plant Sciences, 6: 399-402.
DOI: 10.3923/ajps.2007.399.402
URL: https://scialert.net/abstract/?doi=ajps.2007.399.402
DOI: 10.3923/ajps.2007.399.402
URL: https://scialert.net/abstract/?doi=ajps.2007.399.402
INTRODUCTION
Wheat gluten proteins are classified into two broad groups on the basis of their aggregation and functional properties. These are the gliadins which are present as monomers which interact by mono-covalent forces and the glutenins which form polymers stabilized by inter-chain disulphide bonds. Glutenins fall into two basic classes according to the molecular weight of their subunits: HMW (High Molecular Weight) and LMW (Low Molecular Weight) glutenin subunits (GS).
The HMW subunits account for about 25-35% of the total glutenins (Seilmeier et al., 1991) and have been studied extensively. High molecular weight glutenin subunits are mainly responsible for dough strength in wheat. Dough strength determines the quality of bread and pasta made from bread and durum wheat, respectively (Ram, 2003).
HMW glutenin subunits arise from three loci (Glu-A1, Glu-B1, Glu-D1) located on the long arms of wheat chromosomes 1A, 1B and 1D. Several alleles, responsible for the production of diverse subunits, have been described for each locus. Variation in HMW glutenin subunit composition (e.g., allelic composition) contributes to genetic differences in bread making quality observed among varieties.
Durum wheat (Triticum turgidum L. var. durum Desf.) is widely known to be the best type of wheat for pasta products because of its kernel size, hardiness and golden amber color. Pasta is the general term for foods such as macaroni, spaghetti and noodles. Cooked pasta made from durum wheat semolina retains good firmness and elasticity and is resistant to surface disintegration and stickiness. These characteristics depend more or less on the cultivars that are processed (Dexter and Matsuo, 1980; Autran et al., 1986; Feillet et al., 1989; Bechere et al., 2002).
In durum wheat, genetic studies have revealed that glutenins are encoded at several, complex and highly polymorphic loci (Branlard et al., 1989) and quality differences between cultivars are strongly dependent upon their allelic composition for endosperm storage proteins (Carrillo et al., 1990, 2000). Contradictory results have been reported for the association between high molecular weight (HMW) glutenin subunits and pasta quality (Autran and Feillet, 1987; Autran and Galterio, 1989; Kaan et al., 1993). Consequently, wheat breeders use the natural genetic variation found at the Glu-A1 and Glu-B1 loci encoding HMW glutenin subunits, to improve durum wheat quality (Liu et al., 1996).
Due to this specific nature HMW-GS genes can be utilized as markers of some important characters as well as for characterization of cultivars and lines in wheat („erný and Šašek, 1996). The aim of the present investigation was to analyze the protein characteristics of 25 wild emmer wheat, Triticum turgidum L. var.
dicoccoides, accessions originating from Iran and to compare with 10 exotic durum wheat genotypes from European country for possible differences in their allelic composition at loci encoding HMW glutenin subunits. Suitable accessions can be used for the crossing programs, to introgress positive quality features of exotic germplasm into the existing genetic base.
MATERIALS AND METHODS
Plant materials: Seeds of 25 accessions of Triticum durum and 10 exotic durum wheat genotypes from European country were used. An Iranian cultivar (Falat) and catalogue of alleles for the complex gene loci Glu-A1, Glu-B1 and Glu-D1 (Payne et al., 1979) were applied to identify unknown loci bands and further comparisons.
Protein extraction and electrophoresis: Sodium Dodecyl Sulfate (SDS)-PAGE was carried out according to the Laemmli (1970) procedure as modified by Singh et al. (1991). Gels were prepared 0.75 mm thick, with the separating gel consisting of either 8 or 12% total acrylamide and the stacking gel layer at 4%. The glutenin extracts were characterized using modified method (Singh et al., 1991). The 40 mg of flour was extracted with 60% ethanol two times at room temperature. The high molecular weight glutenin subunits were isolated from the dried pellets using extraction buffer (0.08 M Tris HCl, pH 8.0) with 50% propanol. The supernatant was transferred to new tubes and alkylated with 4-vinylpyridine at 65°C for 30 min. The alkylated glutenins were precipitated with acetone and re-dissolved in sample buffer containing 2% SDS, 40% glycerol, 0.02% bromophenol blue and 0.08 M Tris-HCl 8.0. The glutenins were separated in SDS-PAGE and each gel lane was loaded with a 5 mL aliquot. After electrophoresis, the SDS-PAGE gels were stained with Coomassie Brilliant blue solution and destained with de-ionized distilled water. The HMW subunits of glutenin were designated according to the numbering system of Payne et al. (1979) and the alleles found at the two HMW loci have been identified.
RESULTS AND DISCUSSION
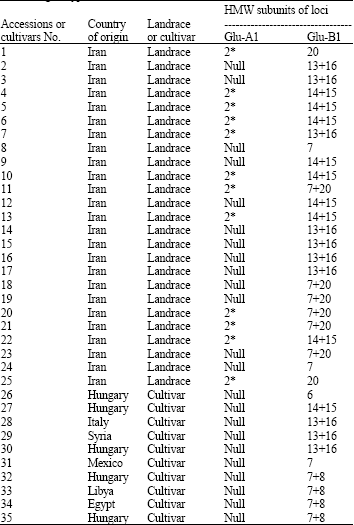
The variation in high-molecular-weight glutenin subunit composition has been reported to account for up to 70% of the variation in bread making quality of European wheats (Branlard and Dardevet, 1985). Twenty-five durum wheat accessions were evaluated for high molecular weight glutenin subunits and 8 different alleles were identified (Table 1). Two of them belonged to the Glu-A1 locus and 6 to Glu-B1. The more frequent allele in Gl-A1 were null (52%), 2*(48 %) and in the Glu-B1 were the bands 14+15 (32%), 13+16 (28%), 7+20 (24%).
| Table 1: | Distribution of high molecular weight glutenin subunits in studied genotypes of durum wheat |
 | |
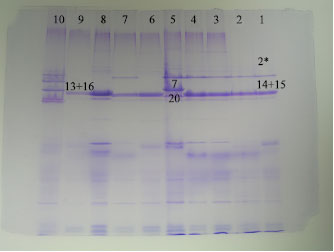 | |
| Fig. 1: | HMW-GS of Iranian durum |
The subunit 2* allele was observed 48% accessions of T. durum of which 50% had the 14+15 allele in Glu-B1 subunit. Alleles, respectively. The composition of High-Molecular Weight (HMW) glutenin subunit of European durum (Fig. 2) wheat is also indicated in Table 1.
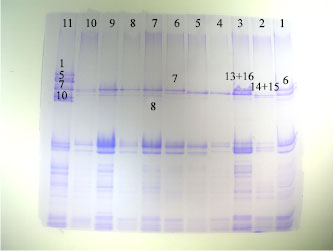 | |
| Fig. 2: | HMW-GS of European durum |
All allele in Glu-A1 were the Null allele and in the Glu-BI were the bands 7+8 in (40%) samples and 13+16 in (30%) samples.
The relationship between HMW-GS and durum wheat quality has been analyzed in several studies. Earlier studies showed no clear relationship between HMW-GS and durum quality (Du cros, 1987) but other studies didn’t showed. (Boggini and Pogna, 1989; Carrillo et al., 1990). Ram (2003) and Aghaei (1995) showed that protein subunit 2* and 1 were found correlated positively with improved dough strength as compared to subunit null. The Glu-B1 subunit 6+8 was associated with slightly stronger gluten type than 7+8 and 13+16, while subunit 20 was associated with weak gluten properties. In fact subunit 6+8 showed superior gluten strength and 7+8, 13+16 showed medium to strong gluten characters. Subunit 20 possesses weak gluten strength and consequently poor pasta- and bread-making quality.
The Glu-B1 subunit 14-15 was observed in 30% Iranian samples and Aghaei (1996) reported that this band increases SDS sedimentation. The result presented indicated that HMW-GS in Iranian durum wheat showed little diversity (Fig. 1) and 28% of them were medium gluten strong that to have subunit 13+16 and 32% that have subunit 20 were weak gluten strength and consequently poor pasta- and bread-making quality.
T. durum accessions from Iran belong to the old world crop, so they are in the position of the source of important agricultural properties for wheat breeding and preservation of genotype variability at the Genbanks (Galova et al., 2002). These results could provide a more complete understanding of the studied collections diversity on high molecular subunits and it will be useful to breeders who now possess a tool to formulate crosses by choosing landraces with appropriate characters. As previously reported, among glu-B1 subunit alleles 14+15, 13+16, 7+8 had positive effects on SDS sedimentation and these alleles should be considered in breeding program. Selection based on alleles at Glu-3 associated with positive effects can improved durum gluten quality. The knowledge of the different association of Glu-3 alleles with quality could help plant breeders to select better genotypes for crossing.
REFERENCES
- Autran, J.C. and G. Galterio, 1989. Associations between electrophoretic composition of proteins, quality characteristics and agronomic attributes of durum wheat. II. Protein-quality associations. J. Cereal Sci., 9: 195-215.
CrossRef - Bechere, E., R.J. Pena-Bautista and D. Mitiku, 2002. Glutenin composition, quality characteristics and agronomic attributes of durum wheat cultivars released in Ethiopia. Afr. Crop Sci. J., 10: 173-182.
Direct Link - Branlard, G., J.C. Aulran and P. Monneveux, 1989. High molecular weight glutenin subunit in durum wheat (T. durum). Theor. Applied Genet., 78: 353-358.
CrossRef - Carrillo, J.M., J.F. Vazquez and J. Orellana, 1990. Relationship between gluten strength and glutenin proteins in durum wheat cultivars. Plant Breed., 104: 325-333.
Direct Link - Carrillo, J.M., M.C. Martnez, C. Moita, M.T. Brites, M.T. Nieto Taladriz and J.F. Vazquez, 2000. Relationship between endosperm proteins and quality in durum wheat (Triticum turgidum L. var. durum). Options Mediterranees, 40: 463-467.
Direct Link - Feillet, P., O. Ait-Mouh, K. Kobrehel and J.C. Autran, 1989. The role of low molecular weight glutenin proteins in the determination of gluten quality of pasta products: An overview. Cereal Chem., 66: 26-30.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Liu, C.Y., K.W. Sheperd and A.J. Rathjen, 1996. Improvement of durum wheat pasta making and breadmaking qualities. Cereal Chem., 73: 155-166.
Direct Link - Seilmeier, W., H.D. Belitz and H. Wieser, 1991. Separation and quantitative determination of high-molecular weight subunits of glutenin from different wheat varieties and genetic variants of the variety Sicco. Z. Lebensm. Unters. Forsch. A, 192: 124-129.
Direct Link - Singh, N.K., K.W. Shepherd and G.B. Cornish, 1991. A simplified SDS-PAGE procedure for separating LMW subunits of glutenin. J. Cereal Sci., 14: 203-208.
Direct Link