
Rafia Azmat
Department of Chemistry, Jinnah University for Women, 5C Nazimabad, 74600-Karachi, Pakistan
Sehrish Hasan
Department of Chemistry, Jinnah University for Women, 5C Nazimabad, 74600-Karachi, Pakistan
Fahim Uddin
Department of Chemistry, University of Karachi, Karachi, Pakistan
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 8 | Page No.: 1246-1250







ABSTRACT
The present study was conducted to investigate the growth, synthesis of amino acids and proteins of Lens culinaris under aluminium (Al) stress. Results showed that nutrients disorder were observed with the loss of apical dominance and both lateral and primary roots were stunted and swollen with increasing concentration of Al. All levels of Al reduced the size and weight of roots, shoots and leaves. Al altered both root and leaf architecture. At low concentration of Al, size of leaf was approximately equal to that of control plant whereas at high dose, reduced leaf expansion were observed. Investigations suggest that a number of physiological and biochemical processes in the plant cell have been affected before growth inhibition occurs. It was concluded that Lens culinaris is an aluminium sensitive species.
PDF Abstract XML References Citation
How to cite this article
Rafia Azmat, Sehrish Hasan and Fahim Uddin, 2007. Aluminium Stress Induced Alteration in Seedling Growth and Alleviation in Protein and Amino Acid Contents of Lens culinaris. Asian Journal of Plant Sciences, 6: 1246-1250.
DOI: 10.3923/ajps.2007.1246.1250
URL: https://scialert.net/abstract/?doi=ajps.2007.1246.1250
DOI: 10.3923/ajps.2007.1246.1250
URL: https://scialert.net/abstract/?doi=ajps.2007.1246.1250
INTRODUCTION
Aluminium (Al) bioavailability has raised much interest in the last two decades because Al is the most abundant metal and third most common element in the earth‘s crust. Development of acid soils that limits crop production is an increasing problem worldwide. Because approximately 50% of the world‘s potentially arable soil are acidic (Mistrik et al., 2000). Acid soils are such an important constraint to agriculture, under mechanism. The target of Al toxicity is the root tips, in which Al exposure causes inhibition of cell elongation and cell division, leading to root stunting accompanied by reduced water and nutrient up take (Petra and Proctor, 2000). Morphological stress was also noticed on depression in root and shoots system, chlorosis of leaves etc., stunted growth and reduced yield were other symptoms (Oleksyn et al., 1996). Pan et al. (1989) suggests that one mode of action by which Al may affect shoot growth is by inhibiting the synthesis and subsequent translocation of cytokinin to the meristematic regions of the shoot. However, the inability of applied cytokinin to counter the restriction imposed by Al on total shoot dry matter production implies the impairment by Al toxicity of other root functions, such as ion and water transport, also played an important role in altering shoot morphology (William et al., 1989). Neogy et al. (2002) reported different concentrations of Al2(SO4)3 were found to have significant effect both on shoot and root length. Leaf area, fresh and dry weight was significantly reduced. Shamsi et al. (2007) found that low pH (4.0) and Al treatments caused marked reduction in root length, shoot height, dry weight, chlorophyll content, (SPAD value) and photosynthetic rate. Karina et al. (2006) reported no significant changes in the ammonium and protein contents in the nodules or root when the plant were treated with 50 μM but at higher concentration like 500 μM. Al-treated seedlings show identical or increased protein level compared to the control (Pietraszewska et al., 2001). In the nitrate added samples the effects of Al/P were less characteristic. The total amount of secreted amino acids has a specific correlation with the external Al3+ concentration. At first, the amino acids secrete normally, but when Al3+ concentration is over 10 mg L-1, the amino acid constitution varies obviously (Wang et al., 2006). Ismail (2005) reported that Al significantly decrease in root length and dry matter yield in the shoots and roots of carrot (Daucus carota L.) and radish (Raphanus sativus L.) plants which leads to decrease in sugars and total amino acids, whereas a significant increase in the proline content of the shoots and roots was detected.
The present study focuses on an integrated assessment of Al toxicity on the weight, length, morphological and physiological changes in the Lens culinaris.
MATERIALS AND METHODS
Ten to fifteen seeds of Lens culinaris were surface sterilized with 0.1% mercuric chloride and germinated at natural environment in petri dishes in the darkness containing Whatman No. 1 filter paper moistened with Hoagland nutrient solution. After 48 h of germination, seeds were transferred to pots containing Hoagland nutrient solution. Aluminium was given in form of aluminium chloride (AlCl3) at increasing concentration (20, 30, 60, 90, 100 and 150 mg L-1). Experiment was conducted during winter in January 2007 in the Department of Chemistry, Jinnah University for Women.
Five uniform plants were selected and dissected in roots and shoots. Root length was recorded individually for each plant by measuring total length of all the roots from root base to maximum of a root and shoot length were measured using standard centimeter scale an interval of 48 h. The plant material was dried at 60°C to achieve a constant weight. Ratio of root length to weight was calculated for each plant and averaged for pot and replications of a treatment (Akmal et al., 2005).
Protein contents in root and shoot extract in water, were treated with Folin cicalteau phenol reagent (half diluted). The extract was left for 30 minutes at room temperature. A blue colored complex was developed. Absorbance of the complex was observed at 650 nm on Schimadzo 160 A UV- Visible spectrophotometer. Amino acids were determined by treating the extract of root and shoot with ninhydrin solution in 10% ethanol and heated to 50 to 70°C for few minutes till purple color appeared. Optical density was recorded at 566 nm.
RESULTS AND DISCUSSION
The toxicity of Al depends upon a number of factor, including, pH and organic matter content. Results showed that (Al) treatment caused a significant decrease in root and shoot length and weight (Fig. 1) (Loboda and Wolejko, 2006). Compared to shoot growth, an adverse effect of the increasing metal concentration to the plant root has already been observed (Ayala-Silva and Al-Hamdani, 1997).
Statistically significant changes in the root biomass, shoot biomass and total plant mass were observed. Root biomass after 2 weeks was reduced by 11% compared with plants grown without Al (Fig. 2-3).
Root W = 7x10-5 Al +0.0252 (R2 = 0.0166) |
| Shoot W = –7x10-5 Al+ 0.0294 (R2 = 0.6132) |
Total Plant Dry Mass (TPDM) = –7x10-6 Al+0.556 (R2 = 0.0002) |
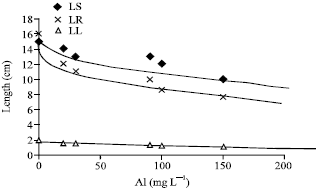 | |
| Fig. 1: | Length of shoot, root and length of Lens culinaris under Al stress, LS = Length of Shoot, LR = Length of Root, LL = Length of Leaf |
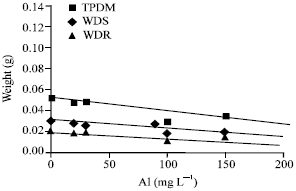 | |
| Fig. 2: | Total plant dry mass, weight of dry shoot and weight of dry root of Lens culinaris, TPDM = Total Plant Dry Mass, WDS = Weight of Dry Shoot, WDR = Weight of Dry Root |
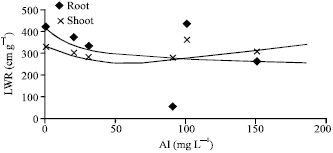 | |
| Fig. 3: | Length weight ratio of root and shoot of Lens culinaris under Al stress |
Leaf Weight Ratio (LWR) for all dose showed a reduction with plant age (Akmal et al., 2005). Moreover, a drastic reduction in the root density was recorded with stunted, compact and swollen structure (Ayala-Silva and Al-Hamdani; 1997; Goransson and Eldhuset, 1991; Clark, 1977). Root length to weight ratio of Lens culinaris treated with different concentration of Al showed reduction. LWR is the ratio of dry mass and average maximum length of all roots and shoots of a plant (Fig. 3).
| LWR = –1.0359 Al+385.21 (R2 = 0.179, root) |
LWR = 0.0512 Al+305.36 (R2 = 0.0076, shoot) |
The values correspond to R2 indicates that aluminum toxicity is the primary factor limiting crop productivity in acidic soils, which comprise large areas of the worlds’ land. The reduction in root and shoot elongation with an increasing concentration of Al has also been observed for many other crops, as the first sign of Al toxicity appears in the root system which becomes stubby as a results of inhibition of elongation of root main axis (Pan et al., 1989).
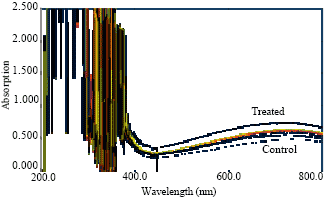
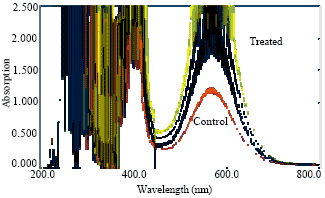
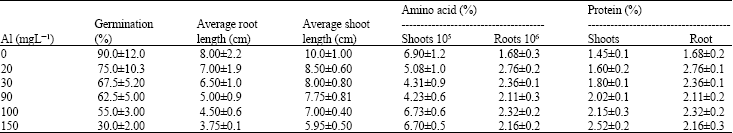
Aluminum (Al) is widely known to inhibit root growth, however it also affects many physiological processes like synthesis of amino acids and protein (Clark, 1977). The concentration of these metabolites increased in roots and shoots of the seedlings in the nutrient solution with added Al. The absorption spectrum of total amino acids and protein contents were shown in Fig. 4-7 and results were reported in Table 1 where a uniform increase in total protein contents was observed with the increase concentration of Al as compared to control plant. These results suggest that soluble protein contents in plant are an important indicator of physiological state as reported earlier that time course experiments for Arabidopsis plants, respond to aluminum toxicity by altering their protein expression and also Al-treated seedlings show identical or increased protein level compared to the control (Pietraszewska, 2001).
The total amino acids contents found increased in roots and shoots with plant exposure to Al. The concentration of these metabolites indicates that Lens culinaris accumulated more metabolites under Al stress than in control.
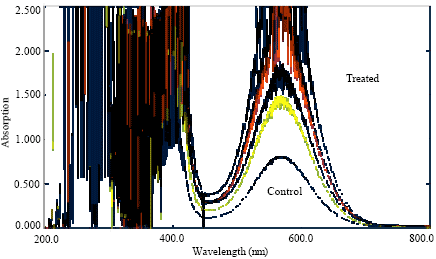
A board range absorption spectrum of amino acid were observed as compared to non treated plant (Fig. 6 and 7) which may be due to affinity of amino acids towards Al. The rhizotoxic Al+3 ions, prevailing at lower pH often enhanced growth at low concentration under acidic conditions that reduced root elongation whereas with regarding protein and amino acids in Al treated plants, it may be related to the different changes in cytoplasmic and membrane proteins which is an agreement with the reported data of different plants (Walter et al., 1999; William et al., 1989; Hu et al., 1995).
Mechanism: The exact mechanism of root growth inhibition is uncertain but two main mechanisms to tolerate high soil Al-including the soil solution Al and inactivating absorbed Al. Recent observation of a plant disorder in Lens culinaris on acid soils, was associated with low leaf that plant communities on very acid soils tend to be slow growing and relatively unproductive, even if they do tolerate the conditions.
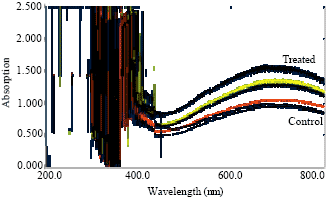 | |
| Fig. 4: | Absorption spectrum of proteins of shoot of Lens culinaris under Al stress |
 | |
| Fig. 5: | Absorption spectrum of proteins of root of Lens culinaris under Al stress |
 | |
| Fig. 6: | Absorption spectrum of amino acids of root of Lens culinaris under Al stress |
Current evidence indicates the tolerance mechanisms have a cost to the plant. This cost can eventually be expected to show up as reduced yield potentials. In the second tolerance mechanism the plant inactivates the absorbed Al, by forming organic complexes with the damaging aluminum ions.
 | |
| Fig. 7: | Absorption spectrum of amino acids of shoots of Lens culinaris under Al stress |
| Table 1: | Some morphological and physiological parameters of Lens culinaris under Al stress |
 | |
It is worth to note that both the tolerance mechanisms seem to involve compromises. A different explaination for the stimulation of plant growth by low Al dose may be based on speciation effects related to the complex solution chemistry of Al that may influence the bio availability of other ions in a way that stimulate plant growth. Decrease in plant growth may be the results of alleviating of proton (H+) toxicity by Al+3. Absorption spectrum of protein and amino acids offered an in increase concentration of protein and amino acid contents in the presence of Al, which may be due to the affinity of protein and amino acids towards Al. Al tends to bind to the phosphate or carboxyl groups rather than to SH groups characteristic for chelatins (Gunse et al., 1997; Khan et al., 2000). However, Snowden et al. (1995) and Wu et al. (2000) suggested that plant metallo-thionein-like protein and phytochelatins may play a role in Al tolerance this may be attributed with the increase in concentration of proteins in leguminous plant under investigation. An Al-induced polypeptide (TA1-18) was identified in wheat that shows homology to a pathogenesis associated (PR) protein (Cruz-Ortega and Ownby, 1993) due to strong interaction of Al+3, the main Al toxic form, with oxygen donor ligands (proteins, nucleic acids, polysaccharides) results in the inhibition of cell division, cell extension and transport of water. The response of the studied material at low pH (4.5) and different Al concentration may be in accordance with Kinraide (1993, 1997) hypothesis visualized Al+3 and H+ competing for common apoplast binding sites. Slaski (1989) reported an increase in protein NAD-kinase activity in root apical meristems of various crops under Al stress while decreased in some most sensitive plants.
CONCLUSIONS
The above investigation revealed that Al inhibits plant growth by interfering with many physiological processes prevailing at low pH, would be capable of alleviating proton toxicity results in the increase in protein and amino acids content of Lens culinaris.
REFERENCES
- Gunse, B., C. Poschenrieder and J. Barcelo, 1997. Water transport properties of roots and root cortical cells in proton and Al-stressed maize varieties. Plant Physiol., 113: 595-602.
Direct Link - Goransson, A. and T.D. Eldhuset, 1991. Effects of aluminium on growth and nutrients uptake of small Picea abies and Pinus sylvestris. Plants J. Trees Str. Fun., 5: 136-142.
Direct Link - Ismail, M., 2005. Aluminium-phosphorus interactions on growth and some physiological traits of carrot and radish plants. J. Acta Agron. Hungarica, 53: 293-301.
Direct Link - Karina, B., G.M. Susana and M.L. Tomaro, 2006. Aluminium stress affects nitrogen fixation and assimilation in soya bean. Plant Growth Regul., 48: 271-281.
Direct Link - Khan, A.A., T. McNeilly and J.C. Collins, 2000. Accumulation of amino acids, proline and carbohydrates in response to aluminum and manganese stress in maize. J. Plant Nutr., 23: 1303-1314.
Direct Link - Mistrik, I., L. Tamas and J. Huttova, 2000. Quantitative changes in maiz membrane induced by aluminium. Biol. Plant, 43: 85-91.
Direct Link - Neogy, M., J. Datta, A.K. Roy and S. Mukheri, 2002. Studies on phytotoxic effect of aluminium on growth and some morphological parameters of Vigna radiata L. Wilczek. J. Environ. Biol., 23: 411-416.
Direct Link - Petra, S.K. and J. Proctor, 2000. Effects of aluminium on the growth and mineral composition of Betula pendula Roth. J. Exp. Bot., 51: 1057-1066.
Direct Link - Mossor-Pietraszewska, T., 2001. Effect of aluminium on plant growth and metabolism. Acta Biochem. Polonica, 48: 673-686.
Direct Link - Shamsi, I.H., K. Wei, G. Jilani and G. Zhang, 2007. Interactions of cadmium and aluminum toxicity in their effect on growth and physiological parameters in soybean. Ziran Kexueban, 8: 181-188.
Direct Link - Snowden, K.C., K.D. Richards and R.C. Gardner, 1995. Aluminum-induced genes. Introduction of toxic metals, low calcium and wounding and pattern of expression in root tips. Plant Physiol., 107: 341-348.
Direct Link - Walter, J.H., N. Schmohl, M. Kollmeier, Franti, ek Balu ka and M. Sivagurul, 1999. Does aluminium affect root growth of maize through interaction with the cell wall-plasma membrane-cytoskeleton continuum. J. Plant Soil, 215: 163-174.
Direct Link - Wang, P., B. Shuping, W. Shi and D. Qiuyan, 2006. Variation of wheat root exudates under aluminum. J. Agric. Food Chem., 54: 10040-10046.
Direct Link - Wu, P., C.Y. Liao, B. Hu, K.K. Yi, W.Z. Jin, J.J. Ni and C. He, 2000. QTLs and Epistasis foraluminum tolerance in rice (Oryza sativa L.) at different seedling stages. Theor. Applied Genet., 100: 1295-1303.
Direct Link