
M. Ramdani
Laboratory of Natural Resources Valorisation, Ferhat Abbas University, 19000 Setif, Algeria
O. Rached
Laboratory of Biology and Envoronnement, Mentouri University, 25000 Constantine, Algeria
T. Lograda
Laboratory of Natural Resources Valorisation, Ferhat Abbas University, 19000 Setif, Algeria
A. Aggoun
Laboratory of Biostatistics, Ferhat Abbas University, 19000 Setif, Algeria
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 8 | Page No.: 1211-1216







ABSTRACT
Genetic variation in monoterpenes foliage was determined in natural populations of Cupressus dupreziana A. Camus. Samples were obtained from 12 populations in 12 regions from Tassili n’Ajjer (Central Sahara of Algeria). A total 20 monoterpenes were detected, with a particular abundance of α-pinene and Δ3-carene. Terpenoids markers allowed the detection of individual chemotypic variability. The high variability found in the monoterpene composition suggested that genetic basis was not the main factor contributing to the extinction of this species. It seems most probable that the reduced population size of this species is related to the effect of Tassili n’Ajjer desertification.
PDF Abstract XML References Citation
How to cite this article
M. Ramdani, O. Rached, T. Lograda and A. Aggoun, 2007. Genetic Diversity in Foliar Terpinoids Among Natural Populations of Cupressus dupreziana in Tassili n`Ajjer (Algeria). Asian Journal of Plant Sciences, 6: 1211-1216.
DOI: 10.3923/ajps.2007.1211.1216
URL: https://scialert.net/abstract/?doi=ajps.2007.1211.1216
DOI: 10.3923/ajps.2007.1211.1216
URL: https://scialert.net/abstract/?doi=ajps.2007.1211.1216
INTRODUCTION
The Cupressus genus belongs to the Cupressaceae family which is represented in Algeria by one widely endemic species found in Tassili n’Ajjer. Cupressus dupreziana is a rare plant and listed by (IUCN, 2006) as an endangered species with only 230 plants (Abdoun and Beddiaf, 2002).
Climatic conditions as well as human activities seem to be the main factors reducing the diversity of this biologic heritage. The two hundred and thirty recorded specimens are relics and aged having more than ten centuries (Leredde, 1957). The ancient cypress of Tassili is likely to die without regenerating themselves in situ. It resembles C. sempervirens L., but differs from that species in its distinctly compressed twigs, the small leaves being usually furnished with glandular pits in the part adnate to the branchlets and especially by the much smaller and longer cones, the seeds, when developed, being of regular form, flattened, sub-orbicular and with wider, thinner wings. There is a possibility that C. dupreziana may be a geographical form of C. sempervirens (Silba, 1983). The differences between the two species are due to the environment (Ferrandes, 1979), other authors support the recognition of C. dupreziana at the specific level (Farjon, 1998; Rushforth et al., 2003; Barrero et al., 2005).
Little information about natural populations of C. dupreziana is available and most studies reported on this cypress tree were done from arboreta. A critical botanical review of this species has been made by Barry et al. (1970).
Phytochemical investigations on Cupressus dupreziana were restricted to isolate and identify terpenoids components and fats (Pauly et al., 1983; Piovetti et al., 1982). Studies related to the determination of essential oils identified 18 terpenoids, six of them were considered as artefacts (Piovetti and Diara, 1977). Some work has been done on the oil isolated from the leaves essential oil (Pauly et al., 1983; Piovetti et al., 1981a; Chanegriha et al., 1997), the Leaves hydrocarbons (Piovetti et al., 1981b), the Oxygenated mono and sesquiterpenes of wood (Piovetti and Diara, 1977; Piovetti et al., 1980a) and diterpenes of wood (Piovetti et al., 1980b). Terpene composition gives valuable information on the in situ population’s variability.
Several genetic studies on the species are currently conducted using vegetative material from specimen planted in botanical gardens (Pichot et al., 1998; Pichot and El-Maataoui, 2000; Pichot et al., 2000, 2001). Terpene variability is determined by genetic as well as ecological factors (Hanover, 1966; Squillace, 1976; Yazdani et al., 1985; Baradat and Yazdani, 1988; Loveless and Hamrick, 1984). Terpenes play an important role in defence mechanisms against pathogens (Rishbeth, 1972; Schuck, 1980), insects attacks (Hanover, 1974) and in the determination of seeds origin (Esteban et al., 1978; Squillace et al., 1980; Yazdani et al., 1982).
We have been working on variation in natural populations of Cupressus dupreziana using foliage monoterpenes as genetic markers. The efficiency of terpenes as genetic marker has been discussed by Strauss et al. (1992). Their utility in studies of evolution and speciation has been demonstrated by Von Rudloff (1975) and Zavarin et al. (1989, 1990).
The present study aims to determine the terpenoids components of Cupressus dupreziana, their distribution, and their qualitative and quantitative variability expression as affected by genetic and ecological parameters in the Tassili n’Ajjer.
MATERIALS AND METHODS
Plant material: The present study is based on the aerial parts analysis of 160 trees sampled in 12 populations from twelve sites in Tassili n’Ajjer as mentioned in Fig. 1. Sampling was non random, the green branchlets of different branches around the crow from natural populations of C. dupreziana and from a planted stand in Djanet, were collected in April 2002-2003. The air dried aerial parts of the plants were subjected to hydrodistillaton for 3 h using a Clevenger-type apparatus. The characteristics of C. dupreziana populations sampled are shown in Table 1.
The plants were authenticated by the staff of the laboratory of natural resources valorisation by comparison with the herbarium specimens. Voucher specimens are deposed in herbarium of the institute of Biology, University Ferhat Abbas Setif, Algeria.
GC analysis: The oils were analyzed by GC on a Hewlett-Packard 5890 GC serial II, equipped with FID, fitted with a SE-54 capillary column, 25 m x 0.25 mm; 0.25 μm film thickness). The column temperature was programmed from 55°C (5mn) to 210°C (4 mn) at the rate of 6°C min-1. The injector and detector temperatures were programmed at 220°C.
Helium was used as carrier gas at a flow rate of 0.6 mL min-1, split ratio 1:50. The analysis by GC-MS performed an a Hewlett-Parckard GC-MS system (5890, series 2; MSD5971A, Hewlett-Parckard), equipped with SE-54 column, under the following conditions splitless (0.7 min), injector 220°C, colum 35°C (0.7 min), 6°C min-1 program to 250°C and the same other GC conditions.
Kovat’s indices were determined by r.t; comparison with n alkanes analyzed with the same temperature program. The compounds were identified by comparing their Retention Indices (RI) (Massada, 1976) and mass spectra with those of authentic samples and/or the NIST/NBS, NIST02, wiley 575 libraries spectra and those of literature (Adams, 2001). The percentage composition of the essential oil is based on peak areas obtained with FID conditions.
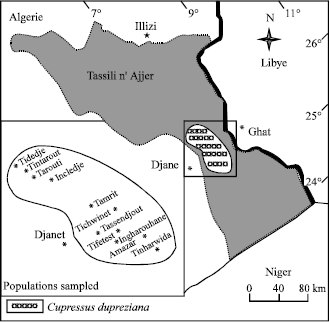 | |
| Fig. 1: | Distribution of Cupressus dupreziana and sampling sites in Tassili n’Ajjers (Central Sahara of Algeria) |
| Table 1: | Description of Cupressus dupreziana populations sampled |
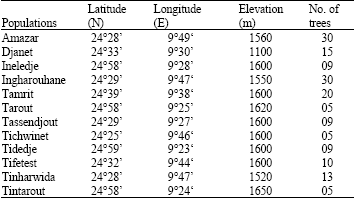 | |
Statistical analysis: Statistical analyses, including multivariate analyse of variance, Principal Components Analysis (PCA) and Cluster analysis, the Unweighted Pair Group Method with Arithmetic mean (UPGMA), were carried out using Statistica 7 software. Six monoterpenes, α-pinene, β-pinene, myrcene, Δ3-carene, limonene and terpinolene, were found in sufficient quantity to be considered in statistical analyses (Hanover, 1966; Baradat and Yazdani, 1988).
RESULTS AND DISCUSSION
The composition of the oils differed quantitatively for wild and cultivated plants (Table 2).
The identified monoterpenoids represent 95% on average of the total monoterpenoid weight analysed. They are common compounds such as α-pinene (54.3%), Δ3-carene (31.2%), limonene (3.1%), myrcene (3.1%), terpinolene (2.9%) and β-pinene (1.7%).
| Table 2: | Means of foliar monoterpenes in populations of Cupressus dupreziana in percent of total monoterpenes |
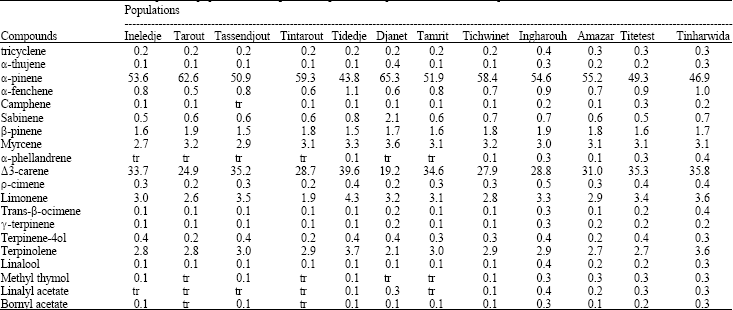 | |
| tr = Trace | |
| Table 3: | Pair wise difference at level 0.05 of Cupressus dupreziana populations with d.f (11 and 145) |
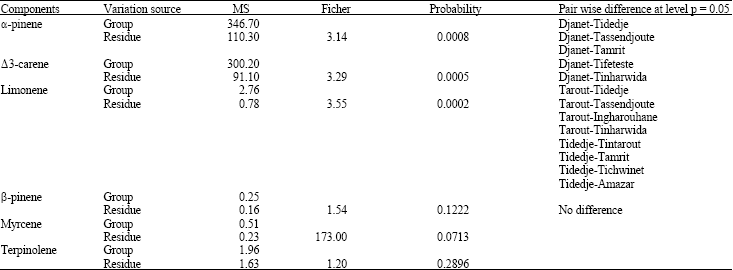 | |
Further analyses revealed 14 monoterpenoids with lower rate (<1%) indicating that within and between populations variability was high. Terpenoids markers allowed detecting the presence of chemotypic variations (Δ3-carene, camphene, α-phellandrene, trans-β-ocimene, γ-terpinolene, linaleol acetate).
Variance analysis: The variance analysis presents the same results, with the HSD test (Honestly significant difference) and Bonferronni test. The Cupressus dupreziana populations are discriminated better by (α-pinene, Δ3-carene and limonene), the pair wise difference at level 0.05 are shoed in Table 3.
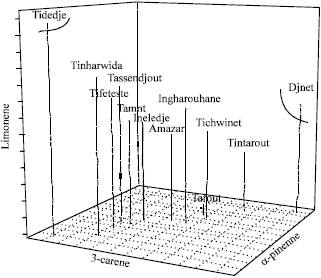
The α-pinene and Δ3-careens it allowed the separation of Djenet population of the natural Tassili n’Ajjer populations, whereas limonene separates the natural populations from the plate of Tassili Ajjer. The highest cophenitic coefficient (0.92) was obtained by using the Mahalonobis distance. These parameters enabled us to produce the Hierarchical tree.
Projection of these populations along the three axes based on correlation of 3 monoterpenes, separated Djanet population and plate populations (Fig. 2), as well as separation within the natural populations of Tarout and Tidedje. This overlapping is related to the elevated rate of the α-pinene and the low variation of β-pinene and myrcene components. The significant monoterpenoids variability found is an indication of the heterogeneity of the genetic structure of the C. dupreziana populations.
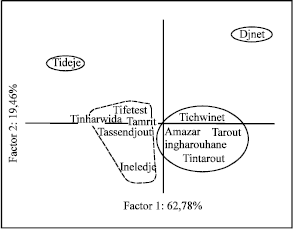
Results of the Principal Components Analysis (PCA): Principal components analysis of six variables resulted in three eigenvectors accounting for 82.3% of the total variation presents in the original data. The first two components explained respectively 62.8 and 19.5% of the total variation. The ordination of population’s means along these two vectors is shown in (Fig. 3).
 | |
| Fig. 2: | Projection of Cupressus dupreziana populations, based on three terpenes |
 | |
| Fig. 3: | Ordination of the first two principal axes of Cupressus dupreziana populations |
This analysis clustered populations in four groups, (Ineledje, Tamrit, Tassendjout, Tinharwida, Tifetest) are characterised by a high rate of Δ3-carene and terpinolene. These populations are found in open craggy Oueds with frequent temporary gueltas (temporary reserve water).
While in the dry gritty Oueds, populations of Tichwinet, Amazar, Ingharouhane, Tarout and Tintarout are found as a specific group characterised by α-pinene, β-pinene and with low contain of myrcene. These populations are localised in rocky and dry rivers. Tidedje with the highest rate of Δ3-carene, limonene and terpinolene, is insulated on dry sandy grounds. Whereas Djanet-planted stand population, localised in humid gritty soils, is characterised by limonene and myrcene.
Results of cluster analysis: Genetic analyses were carried out using 6 terpenes including some compounds that have been shown in other species of Cupressus to be under the control of a single locus with two alleles.
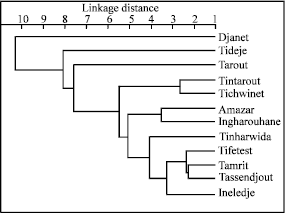 | |
| Fig. 4: | Dendrogram based on Mahalanobis distance |
The dendrogram based on a UPGMA clustering is shown in Fig. 4, confirms presence of the same three groups obtained from ACP analyses and emphasizes the distinctiveness of Djanet Population. Distinctive leaf oil monoterpene patterns can be recognized in Djanet population. These are characterized by major amounts of α-pinene (65.3%), sabinene (2.1%) and myrcene (3.6%). Djanet populations were also characteristic for their lowest rate of Δ3-carene (19.2%) and terpinolene (2.1%). In the group of the natural populations, Tidedje and Tarout are isolated. The remainder of the populations is separated into several groups with high degrees of similarity.
Based on the rate of the Δ3-carene, Cupressus dupreziana populations are divided into two main groups, This component translates the effect of the environmental factors on populations behaviour, as well as the response of individuals to the ecological pressure (Tobolski and Hanover, 1971; Baradat et al., 1975).
Djanet population is more divergent, with a low similarity, from the other populations. This population contains the lowest rate of Δ3-carene (18.13%) and 3 monoterpenes with maximal values. Populations of Tassili n’Ajjer grouped apart into two sub-groups. These populations grow under very harsh ecological conditions revealed by increases in the rate of the Δ3-carene. The separation of the Tidedje population is due to the presence of a high rate of Δ3-carene.
Aggregation of Tassili n’Ajjer populations into small groups is an indication of monoterpenoids variability in this species. High amount of monoterpenes is an indicator of genetic variability (Baradat et al., 1975; Forrest, 1980).
Monoterpenoids analysis of wild cypress populations is a useful complement to morphological studies. Integrating measurements of genetic variability based on monoterpenes with information on ecological requirements and micro evolutionary mechanisms is important for both preserving adaptability and managing natural populations.
Analyses of C. dupreziana endemic species of the Tassili n’Ajjer showed a monoterpenes variability within populations. The UPGMA analysis of terpene traits, confirm the presence of this variability, but no clear link between geographical location and genetic structure can be identified.
C. dupreziana endemic to the Tassili n’Ajjer region are believed to have originated from a Mediterranean variety of C. sempervirens (Silba, 1983). Present results show that the C. dupreziana terpenic composition differs from other species of cypress.
The study of Barrero et al. (2005) show that the composition of C. atlantica essential oil of Morocco different from that of C. dupreziana. Rushforth (2003) suggested that C. dupreziana is closer genetically to C. atlantica than of C. semperviren and it is essentially identical to the cultivated forms of C. dupreziana throughout the Mediterranean area. This difference can be the manifestation of the very hard ecological conditions of Tassili n’Ajjer and the insulation of this species.
The restricted localization of C. dupreziana, probably led to genotypes reduction. For maintain the population of the genetic diversity require bigger than 500 individuals Raddi and Sümer (1999) and all loss of the genetic diversity can only be restored with a big number of generations (Nei et al., 1975).
The populations of Tassili n’Ajjer are an important reservoir of genetic diversity. Cypress population located at Djanet is the most differentiated and have an important regeneration capacity. Therefore the conservation of this species outside of its natural territory is possible. In the Tassili n’Ajjer, under present climatic conditions preservation of specimens is possible, provided that anthropic effects could be seriously controlled.
ACKNOWLEDGMENT
The authors acknowledge the Tergui’s guides (Lahbib. Wendoken and Ahmed Agha).
REFERENCES
- Abdoun, F. and B. Beddiaf, 2002. Cupressus dupreziana A. Camus: Distribution, decline and regeneration on the Tassili n'Ajjer, Central Sahara. Comptes Rendus Biologies, 325: 617-627.
Direct Link - Barrero, A.F., M.M. Herrador, P. Arteaga, J. Quilez and M. Piedra et al., 2005. Chemical composition of the essential oils of Cupressus atlantica Gaussen. J. Essential Oil Res., 17: 437-439.
Direct Link - Barry, J.P., B. Belin, J.C. Celles, D. Dubost, I. Faurel and P. Hethener, 1970. Essai de monographie du Cupressus dupreziana A. Camus, cypres endemique du Tassili des Ajjers (Sahara central). Bul. Soc. Hist. Nat. Afr. Nord, 61: 95-176.
Direct Link - Hanover, J.W., 1966. Environmental variation in the monoterpenes of Pinus monticola Dougl. Phytochemistry, 5: 713-717.
Direct Link - Loveless, M.D. and J.L. Hamrick, 1984. Ecological determinants of genetic structure in plant populations. Annu. Rev. Ecol. Syst., 15: 65-95.
CrossRefDirect Link - Pauly, G., A. Yani, L. Piovetti and C. Bernard-Dagan, 1983. Volatile constituents of the leaves of Cupressus dupreziana and Cupressus sempervirens. Phytochemistry, 22: 957-959.
Direct Link - Pichot, C., A. Borrut and M. El-Maataoui, 1998. Unexpected DNA content in the end osperm of Cupressus dupreziana A. Camus seeds and its implications in the reproductive process. Sex. Plant Reprod., 11: 148-152.
Direct Link - Pichot, C. and M. El-Maataoui, 2000. Unreduced diploid nuclei in Cupressus dupreziana A. Camus pollen. Theor. Applied Genet., 101: 574-579.
Direct Link - Pichot, C., B. Fady and I. Hochu, 2000. Lack of mother tree alleles in zymograms of Cupressus duprezianaA. Camus embryos. Ann. For. Sci., 57: 17-22.
Direct Link - Pichot, C., M. El-Maataoui, S. Raddi and P. Raddi, 2001. Surrogate mother for endangered Cupressus. Nature, 412: 39-39.
PubMed - Piovetti, L. and A. Diara, 1977. Sesquiterpenes from Cupressus dupreziana. Phytochemistry, 16: 103-106.
Direct Link - Piovetti, L., C. Francisco, G. Pauly, O. Benchabane and A. Diara, 1981. Volatile constituents of Cupressus dupreziana and the sesquiterpenes of Cupressus sempervirens. Phytochemistry, 20: 1299-1302.
Direct Link - Raddi, S. and S. Sumer, 1999. Genetic diversity in naturel Cupressus sempervirens L. Populations in turkey. Biochem. Syst. Ecol., 27: 799-814.
Direct Link - Rushforth, K., R.P. Adams, M. Zhong and Ma. X-qiang, 2003. Variation among Cupressus species from the eastern hemisphere based on Random Amplified Polymorphic DNAs (RAPDs). Biochem. Syst. Ecol., 31: 17-24.
Direct Link - Strauss, S.H., J. Bousquet, V.D. Hipkins and Y.P. Hong, 1992. Biochemical and molecular genetic markers in biosystematic studies of forest trees. New Forest, 6: 125-158.
Direct Link - Yazdani, R., J.E. Nilson and T. Ericsson, 1985. Geographical variation in the relative proportion of monoterpenes in cortical oleoresin of Pinus sylvestris in Swiden. Silvae Genet., 34: 201-208.
Direct Link