
L. Zarei
Collage of Agricultural, Razi University of Kermanshah, Iran
E. Farshadfar
Collage of Agricultural, Razi University of Kermanshah, Iran
R. Haghparast
Dryland Agricultural Research Institution, Sararood, Iran
R. Rajabi
Dryland Agricultural Research Institution, Sararood, Iran
M. Mohammadi Sarab Badieh
Collage of Agricultural, Razi University of Kermanshah, Iran
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 8 | Page No.: 1204-1210







ABSTRACT
The objective of this study was to evaluate the ability of several selection indices to identify drought resistant cultivars under field and laboratory conditions in Kermanshah, Iran. Twenty bread wheat genotypes differing in yield performances were evaluated using a RCBD design with three replications under two different environments (irrigated and rein-fed) during 2004-2005 growing season. Grain yield of the genotype in the both conditions, Stress Tolerance Index (STI), was calculated for each genotype. Moreover Proline amino acid concentration (PC) in flag leaf of each genotype was also measured in grain filling stage in the stressed condition. Cell Membrane Stability (CMS) of leaf tissues in seedling stage was calculated for each genotype grown in the pots in greenhouse. Germination Stress Index (GSI), coleoptile length and root length, number of roots, Promptness Index in Stress (PIS) and non-stress (PINS) condition were recorded in the controlled condition of laboratory by using a 2x20 factorial experiment within completely randomized design with three replications under two different stress and non-stress (normal) water regimes. The result of analysis of variance exhibited highly significant difference among the genotypes for the most of the traits. Drought stress induced by PEG (polyethylene glycol) decreased all of the traits. Root length was the most sensitive to drought stress. Correlation analysis indicated strong association between STI and CMS and PC. It was inferred from the study that the greater drought tolerance corresponded to the greater cell membrane stability and proline accumulation. Also, highly significant correlation was observed between GSI, STI and PIS. The results imply that these criteria may be screened for indirect selection of drought tolerance in the initial stages of the bread wheat growth.
PDF Abstract XML References Citation
How to cite this article
L. Zarei, E. Farshadfar, R. Haghparast, R. Rajabi and M. Mohammadi Sarab Badieh, 2007. Evaluation of Some Indirect Traits and Indices to Identify Drought Tolerance in Bread Wheat (Triticum aestivum L.)
. Asian Journal of Plant Sciences, 6: 1204-1210.
DOI: 10.3923/ajps.2007.1204.1210
URL: https://scialert.net/abstract/?doi=ajps.2007.1204.1210
DOI: 10.3923/ajps.2007.1204.1210
URL: https://scialert.net/abstract/?doi=ajps.2007.1204.1210
INTRODUCTION
Drought stress particularly at seedling stage is a major determinant of wheat production in many parts of the world (Dhana et al., 2002). The research into drought stress has until now diverted towards whole plant with particular emphasis on the exploitation of high yield potential under normal and drought stress conditions or selecting the genotypes for morphological and physiological characters responsible for drought resistance (Blum, 1988). The genotype may possess high potential yield under water non-limiting conditions, or may have escaped from drought by reaching maturity before drought develops. Yield in drought-prone environments may be considered to be affected by three components, viz. yield potential, appropriate phenology and drought tolerance. Thus, if drought screening is based on grain yield, genotypes selected may have high potential yield or appropriate phenology, but not drought tolerance (Ouk et al., 2006). A wide range of putative selection criteria that could be used to increase drought tolerance in plants is available. There are, however, very few examples of success obtained using physiological traits in breeding programs. The main reason for this is that few of these traits have been studied in terms of their functional significance to seed yield. In addition, screening techniques using these traits have usually proved to be laborious and costly (Turner et al., 2001).
The term, osmotic stress, is commonly used to refer to situations where insufficient water availability limits plant growth and development. It can result from drought as well as from excessive salinity (Zhu et al., 1997). Polyethylene Glycols (PEGs) are a group of neutral osmotically active polymers with a certain molecular weight. PEG widely used to induce water stress, is a nonionic water polymer, which is not expected to penetrate into plant tissue rapidly (Nepomuceno et al., 1998). PEG is most frequently used in plant water deficit studies to induce dehydration by decreasing the water potential of the nutrient solution (Krizek, 1985; Jacomini et al., 1988). The measurement of solute leakage from plant tissue is a long-standing method for estimating membrane integrity in relation to environmental stresses, growth and development and genotypic variation (Blum and Ebercon, 1981; Bandurska, 2000). In this regard, the degree of cell membrane stability is considered to be one of the best physiological indicators of drought stress tolerance (Kocheva et al., 2004).
The accumulation of free proline in response to osmotic stress is a widely distributed adaptive reaction not only in plants but in other organisms as well (McCue and Hanson, 1990; Delauney and Verma, 1993). This amino acid plays a number of important functions (Hare and Cress, 1997). Of considerable importance among them is its interaction with membrane proteins which is probably involved in the maintenance of the cell membrane structure (Bohnert and Jensen, 1996).
Growing of seedling and studying their responses to environmental stress is an efficient process of screening the genotypes for drought resistance (Winter et al., 1988). The seedling grown under laboratory condition can be altered simultaneously for the entire population. Moreover, the biochemical and physiological changes related to water stress will be more uniform in laboratory conditions in contrast to the intact plant system which has comparatively more heterogeneous environment (Dhana et al., 2002). The present investigation was therefore undertaken to study some promising characters and indices related to drought resistance/tolerance.
MATERIALS AND METHODS
The experiment was conducted during 2004-2005 growing season at the experimental farm, laboratory and greenhouse of dry land agricultural research institution Sararood station, Kermanshah, Iran (47°20’ N latitiude, 34°20’ E longitude and 1351 m altitude). Climate in this region is classified as semi-arid with mean annual rainfall of 478 mm.
Stress Tolerance Index (STI): In the field, twenty genotypes of bread wheat were evaluated using a RCBD design with three replications under two different environments (irrigated and rein-fed) during 2004-2005 growing season. Each plot consisted of 6 rows with 3 m in length and spaced by 20 cm. Seeds were planted with 400 seeds density m-2. After separation of border effects from each plots, yield potential (Yp) and stress yield (Ys) were measured. Stress Tolerance Index (STI) of the grain yield of the genotype in both conditions was calculated using the formula suggested by Fernandez (1992):
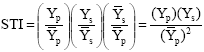 |
Where, Ys, Yp and ![]() represent yield under stress condition, yield under non-stress condition and overall mean of the entries in non-stress condition, respectively.
represent yield under stress condition, yield under non-stress condition and overall mean of the entries in non-stress condition, respectively.
Free proline content: Free proline content was measured according to Bates et al. (1973).
Leaf membrane stability: For determination of Cell Membrane Stability (CMS), 20 leaf pieces (2 cm each) were taken from 15 day old seedlings grown in the pots in green house based on RCBD design with three replications. Distilled and deionised water was used for washing of samples as treatments. For desiccation treatment, leaf sections were submerged in 20 mL of 30% polyethyleneglycol-6000 (PEG-6000) solution in test tubes and allowed to stand in the solution for 24 h at 10°C. Control samples were submerged in water. The samples were then washed rapidly three times with deionised water. For both desiccated and control samples, 20 mL of water was added and leaf discs were maintained for 24 h at 10°C. Then the tubes were warmed to 45°C for 1 h, shaken well and electrical conductivity (C), was measured using an electrical conductivity meter. The leaf tissue were killed by boiling in a water bath for 30 min, then cooled to room temperature and electrical conductivity (C2) was measured, cell membrane stability of leaf tissues was calculated as the percentage injury using the following equation:
CMS (%) = 1-I (also in %) |
| T1 and T2 | = | First and second conductivity measurement of desiccation treatment, respectively |
| C1 and C2 | = | First and second conductivity measurement of control, respectively |
Germination and seedling characters: Seeds were initially treated with 1.5% sodium hypochlorite for 15 min. Residual chlorine was eliminated by through washing of seeds with distilled water. Twenty seeds were then germinated on filter paper in Petri-dishes of 25 mm diameter in an incubator at 22±2°C. The experiment was conducted under normal (0 bar) and drought stress (-0.8 MPa) created with the help of polyethyleneglycol-6000 (PEG-6000) by the method suggested by Michel and Kauffman (1972). The experiment was a 2x20 factorial within completely randomized design with three replications under two different stress and non-stress (normal) water regimes described above. The first factor (A) which had two levels (stress and non-stress treatments) and the second factor (B) which had 20 levels (genotypes) were compared. In the stress and normal treatments 6 mL of PEG solution and distilled water added to the each Petri dish respectively, in the 1 day and 4 mL added in 6 day to compensate the losses due to evaporation. The emergence of 2 mm of radical and plumule was taken as the criterion for germination.
Germination stress index: After 10 days the number of germinated seeds was recorded and the Promptness Index (PI) and Germination Stress Index (GSI) were calculated using the formula proposed by Bouslama and Schapaugh (1984):
PI = nd2 (1.0) + nd4 (0.8) + nd 6 (0.6) + nd8 (0.4) + nd10 (0.2) |
In which nd2, nd4, nd6, nd8, nd10 represent the percentage of germinated seeds after 2, 4, 6, 8 and 10 days after sowing, respectively.
Where, PIS is PI under drought stress condition and PINS is PI under normal condition.
The data for germination percentage, root length (cm), coleoptile length (cm) and number of primary roots were recorded on the 10th day after sowing.
Statistical analyses were performed using SPSS and MSTAT-C software.
RESULTS AND DISCUSSION
The results of analysis of variance of the yield in both conditions showed highly significant differences between genotypes (Table 1). The yield of the genotypes in stress and non-stress conditions in field study was used to calculate Stress Tolerance Index (STI). There were significant differences between genotypes for PC. On the basis of analysis of variance, there were no significant differences between the genotypes for CMS.
The results indicated that the highest values of grain yield under stress condition (Ys) and STI belonged to genotype 8.
| Table 1: | ANOVA based on RCBD design for the traits of bread wheat in the field and green house |
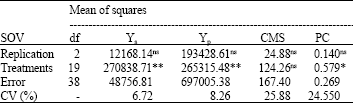 | |
| **: Significant at 1% level, ns: not significant, CV: Coefficient of variation (%) | |
| Table 2: | Mean comparison between bread wheat genotypes for the characteristics under investigation |
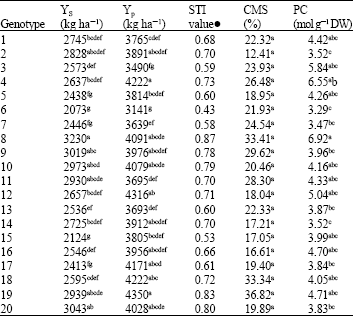 | |
| ! No statistical analysis performed; Means followed by the same letter are not significantly different according Duncan’s test. Yp: Yield potential (kg ha-1), Ys: stress yield (kg ha-1), STI: Stress Tolerance Index, PC: Proline amino acid concentration mol g-1 dry weight, CMS: Cell Membrane Stability (%) | |
| Table 3: | ANOVA based on factorial experiment for germination characters of bread wheat in the laboratory |
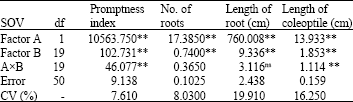 | |
| **: Significant at 1% level, ns: not significant, CV: Coefficient of Variation | |
Also, this genotype had the high yield in non-stress condition (Table 2). The genotypes 8 and 4 had the highest amount of proline content.
The results of analysis of variance of the traits measured in laboratory are given in Table 3. Highly significant differences were observed among the levels of the first factor (A) for all the traits and among the levels of the second factor (B) for all the traits. Drought stress caused by PEG decreased all the traits significantly (factor A). It has been found that decrease in water potential gradient between seeds and their surrounding media adversely affect seed germination and subsequent growth process.
| Table 4: | ANOVA based on CRD design for germination characters of bread wheat in drought stress condition in the laboratory |
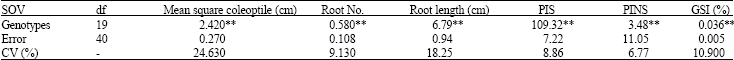 | |
| **: Significant at 1% level, CV: coefficient of variation (%) | |
| Table 5: | Mean comparison between bread wheat genotypes for the characteristics under investigation |
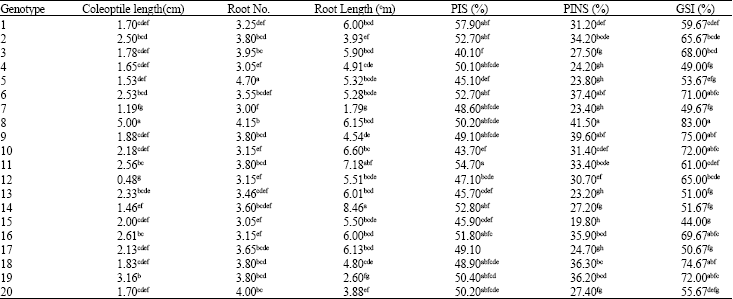 | |
| Means followed by the same letter are not significantly different according Duncan’s test; PIS: Promptness index in stress condition, PINS: Promptness index in non-stress Condition, GSI: Germination Stress Index (%) | |
| Table 6: | Simple correlation coefficient of the traits |
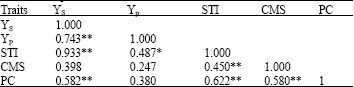 | |
| ** and *: Significant at 1 and 5% level, respectively. Yp: Yield potential (kg ha-1), Ys: Stress yield (kg ha-1), STI: Stress Tolerance Index, PC: Proline amino acid concentration (mol g-1 dry weight), CMS: Cell Membrane Stability (%) | |
Reduced fresh weight in elevated solution of PEG (data not presented) indicated reduced water uptake by the germinating seeds is supported by the work of Alam et al. (2002). The effect of drought stress was more noticeable on root length. Solution of PEG (-0.8 MPa) reduced root length by 49%. Alam et al. (2002) reported that the plumule length of rice was more sensitive to PEG than either germination or germination rate. Xiping (1999) showed that plumule elongation of the cell expanding growth was the most sensitive stage to water stress.
Analysis of variance in both of stress and normal conditions was done separately. Table 4 shows analysis of variance in the drought stress condition based on CRD design. There were significant differences among all of traits in drought stress imposed by PEG. Comparison of means grouped the entries in different classes (Table 5). The longest coleoptile belonged to the genotypes 8, 19 and 16. The primary roots were longer in the genotypes 10, 11, 14 and 8. Genotypes 8, 5 and 20 had the most number of roots.
The highest value of PI in stressed condition belonged to genotypes 8, 6 and 9. Using PI in stressed and non-stress condition value, the GSI were calculated for all the genotypes. Genotypes 8, 9 and 18 had the highest value of GSI, respectively. Sapara et al. (1991) reported that in wheat and triticale, genotypes with higher GSI had also higher drought tolerance. This is agreement with result of Mohammadi et al. (2003).
For a trait to be considered as a selection criterion for plant breeding, it must associate with yield. It is therefore essential to determine whether or not yield was correlated with a particular physiological trait. However, because the relationship of yield to physiological attributes is not clearly understood (Gupta et al., 2001), an essential step would consist of searching for simple correlations between variable traits and yield under stress or drought-response indices. To study the relationship, simple correlation between each pair of the traits was calculated (Table 6). Grain yield under irrigated condition was correlated with rein-fed condition suggesting that a high potential yield under optimum condition may result in improved yield under stressed condition. Thus indirect selection for a drought-prone environment based on the results of optimum condition will be efficient. These results are not in agreement with those of Sio-Se Marde et al. (2006) who found that cultivars of wheat with low yield potential were more productive under stress condition.
| Table 7: | Simple correlation coefficient of the traits of bread wheat genotypes |
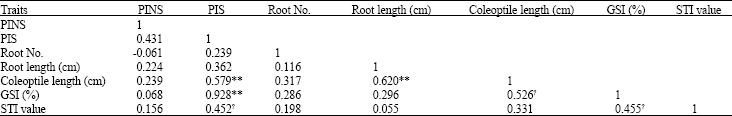 | |
| ** and *: Significant at 1 and 5% level, respectively, STI: Stress Tolerance Index, PIS: Promptness index in stress condition, PINS: Promptness index in non-stress condition, GSI: Germination Stress Index | |
STI was positively correlated with Yp and Ys. With selection for higher STI (Fernandez, 1992) genotypes with high potential under both conditions are selected. In other word, the higher the value of STI for a genotype, the higher its stress tolerance and yield potential. PC was positively correlated with STI (Table 6). Mohsenzadeh et al. (2006) reported that when the drought condition was extended to 18 days, free proline amount was increased for 30 times. Proline accumulation in great amounts is an established fact for many plant and animal species which most probably is not occasional (Kocheva et al., 2003). However, the exact mechanisms of its action in the adaptation to stress are not completely understood yet. Nevertheless, the accumulation of proline could be regarded as an indicative reaction in response to stress at the cellular level (Delauney and Verma, 1993). A possible association of proline accumulation with the drought resistance of different genotypes will obviously have a major practical importance: It would provide an easy screening criterion to plant breeders and help agronomists to assess the field performance of genotypes. The main problem, however, lies in the ambiguity of the term drought resistance for field crops (Karamanos, 1995). The criteria change among investigators: Singh et al. (1972) used the yield stability index over a range of water stress to associate proline accumulation with the drought resistance of barley varieties. We tried to examine proline accumulation in wheat genotypes using STI and CMS to associate proline accumulation with the drought resistance.
A significant positive correlation was found between CMS and STI indicating that the higher the CMS, the higher is drought stress (Fig. 1). The Cell Membrane Stability (CMS) is a measurement of resistance induced in plants that are exposed to desiccation created artificially by polyethylene glycol (Sullivan, 1971). In the present study the CMS technique was used to screen for drought tolerance of different bread wheat genotypes. Kocheva et al. (2004) showed that greater water loss corresponded to greater membrane damage. This is in agreement with our experiment. The results showed that the use of laboratory test systems simulating drought by application of agents such as PEG can reveal character of the resistance of the leaf cells to dehydration induced by drought stress.
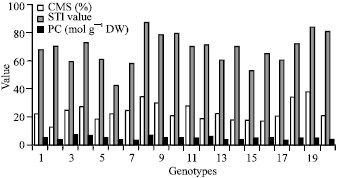 | |
| Fig. 1: | Relation of CMS, PC and STI |
The highly positive correlation found between CMS and PC. The more drought tolerant genotypes accumulate more proline when subjected to dehydration, at the same time less damage of the cell membranes is observed.
The opinion shared by many authors that free proline might be involved in the stabilization of the membranes during water stress is confirmed in our experiments (Kocheva et al., 2003).
It was not a significant correlation between CMS and yield stress.
The PIS and PINS showed high and positive correlation with GSI (Table 7). Farshadfar et al. (2002) found high correlation coefficient between PI and GSI. There was a significant correlation between coleoptile length and GSI indicating that the longer the coleoptile, the higher is drought tolerance. Coleoptile length was also screened as index of drought tolerance (Bouslama and Schapaugh, 1984; Farshadfar et al., 1993). Root length and root number were not associated with GSI, neither are they related to drought response indices in this study. This is not in agreement with Mohammadi et al. (2003) who found high and positive correlation between primary root number, root length and GSI. The GSI and PIS showed significant positive correlation with STI. Tikhonov (1973) found high correlation coefficients between PI, STI and GSI. It is concluded that tolerance to water stress in growing seedlings can be screened for by using PEG-containing nutrient solutions.
In conclusion therefore, in a breeding program they can be used as a selection criterion for drought tolerance, under controlled conditions at earlier stage of the plant growth. These criteria may be screened for indirect selection of drought tolerance in the initial stages of the crop growth.
REFERENCES
- Alam, M.Z., T. Stuchbury and R.E.L. Naylor, 2002. Effects of NaCl and PEG induced osmotic potentials on germination and early seeding growth of rice cultivars differing in salt tolerance. Pak. J. Biol. Sci., 5: 1207-1210.
Direct Link - Bandurska, H., 2000. Does proline accumulated in leaves of water deficit stressed barley plants confine cell membrane injury? I. Free proline accumulation and membrane injury index in drought and osmotically stressed plants. Acta Physiologiae Plantarum 22: 409-415.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Blum, A., 1988. Plant Breeding for Stress Environments. CRC Press, Boca Raton, USA., ISBN-10: 0849363888, Pages: 223.
Direct Link - Bohnert, H.J. and R.G. Jensen, 1996. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol., 14: 89-97.
CrossRefDirect Link - Bouslama, M. and W.T. Schapaugh, 1984. Stress tolerance in soybeans. I. Evaluation of three screening techniques for heat and drought tolerance. Crop Sci., 24: 933-937.
CrossRefDirect Link - Delauney, A.J. and D.P.S. Verma, 1993. Proline biosynthesis and osmoregulation in plants. Plant J., 4: 215-223.
CrossRef - Dhana, S.S., G.S. Sethi and R.K. Behel, 2002. Inheritance of seedling traits under drought stress conditions in bread wheat. Cereal Res. Commun., 30: 293-300.
Direct Link - Farshadfar, E., R. Mohammadi and J. Sutka, 2002. Association between field and laboratory predictors of drought tolerance in wheat disomic addition lines. Acta Agron. Hung., 50: 377-381.
Direct Link - Fernandez, G.C.J., 1992. Effective selection criteria for assessing plant stress tolerance. Proceedings of the International Symposium on Adaptation of Vegetables and other Food Crops in Temperature and Water Stress, August 13-16, 1992, Shanhua, Taiwan, pp: 257-270.
Direct Link - Gupta, N.K., S. Gupta and A. Kumar, 2001. Effect of water stress on physiological attributes and their relationship with growth and yield of wheat cultivars at different stages. J. Agron. Crop Sci., 186: 55-62.
CrossRef - Hare, P.D. and W.A. Cress, 1997. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul., 21: 79-102.
CrossRefDirect Link - Jacomini, E., A. Bertani and S. Mapelli, 1988. Accumulation of PEG 6000 and its effects on water content and carbohydrate level in water-stressed tomato plants. Can. J. Bot., 66: 970-973.
Direct Link - McCue, K.F. and A.D. Hanson, 1990. Drought and salt tolerance: Towards understanding and application. Trends Biotechnol., 8: 358-362.
CrossRef - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Mohammadi, R., E. Farshadfar, M. Aghaee-Sarbarzeh and J. Sutka, 2003. Locating QTLs controlling drought tolerance criteria in rye using disomic addition lines. Cereal Res. Commun., 31: 257-264.
Direct Link - Mohsenzadeh, S., M.A. Malboobi, K. Razavi and S. Farrahi-Aschtiani, 2006. Physiological and molecular responses of Aeluropus lagopoides (Poaceae) to water deficit. Environ. Exp. Bot., 56: 314-322.
Direct Link - Nepomuceno, A.L., D.M. Oostrerhuis and J.M. Stewart, 1998. Physiological responses of cotton leaves and roots to water deficit induced by Polythylene. Glycol. Environ. Exp. Bot., 40: 29-41.
Direct Link - Ouk, M., J. Basnayake, M. Tsubo, S. Fukai, K.S. Fischer, M. Cooper and H. Nesbitt, 2006. Use of drought response index for identification of drought tolerant genotypes in rein-fed lowland rice. Field Crop Res., 99: 48-58.
Direct Link - Sapra, V.T., E. Savage, A.O. Anaele and C.A. Beyl, 1991. Varietal differences of wheat and triticale to water stress. J. Agron. Crop Sci., 167: 23-28.
CrossRefDirect Link - Singh, T.N., D. Aspinall and L.G. Paleg, 1972. Proline accumulation and varietal adaptability to drought in barley: A potential metabolic measure of drought resistance. Nat. New Biol., 236: 188-190.
PubMedDirect Link - Mardeh, A.S.S., A. Ahmadi, K. Poustini and V. Mohammadi, 2006. Evaluation of drought resistance indices under various environmental conditions. Field Crop Res., 98: 222-229.
CrossRefDirect Link - Turner, N.C., G.C. Wright and K.H.M. Siddique, 2001. Adaptation of grain legume (pulses) to water-limited environments. Adv. Agron., 71: 193-231.
CrossRef - Zhu, J.K., P.M. Hasegawa, R.A. Bressan and H.J. Bohnert, 1997. Molecular aspects of osmotic stress in plants. Crit. Rev. Plant Sci., 16: 253-277.
CrossRefDirect Link