
M.A. Rahman
Department of Crop Science, Faculty of Agriculture, University Putra Malaysia, Malaysia
J. Kadir
Department of Plant Protection, Faculty of Agriculture, University Putra Malaysia, Malaysia
T.M.M. Mahmud
Department of Crop Science, Faculty of Agriculture, University Putra Malaysia, Malaysia
R. Abdul Rahman
Department of Process and Food Engineering, Faculty of Engineering, University Putra Malaysia, Malaysia
M.M. Begum
Department of Plant Protection, Faculty of Agriculture, University Putra Malaysia, Malaysia
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 1 | Page No.: 12-20







ABSTRACT
Of the 27 antagonistic bacteria isolated from the fructosphere of papaya and screened by dual and concomitant test, four isolates of bacteria (B23, B19, B04 and B15) had high antagonistic activities against Colletotrichum gloeosporioides of papaya. Using the Biolog system, isolates B23 and B19 were identified as Burkholderia cepacia and B04 and B15 as Pseudomonas aeruginosa. Both B. cepacia and P. aeruginosa strongly inhibited the fungal growth by an average of 74.13 and 68.45%, respectively during in vitro screening on PDA medium. The bacteria also produced volatile as well as diffusible substances. Malformation of hyphae occurred in the presence of both bacteria. Hyphae were thickened, vacuolar and many swellings occurred in them or at the tips of hyphal strand. However, B. cepacia was found most efficacious biocontrol agent in this study. Total inhibition (100%) in spore germination was noted in presence of B. cepacia at 24 h after treatment. Filter sterilized culture filtrate of B. cepacia also significantly inhibited the mycelial growth (59.2%) and spore germination (100%) of the test fungus, thus suggesting that an antibiotic substance (s) may be produced by the bacterium. Therefore, in vitro activities of the B. cepacia against C. gloeosporioides of papaya in this study suggested that the bacterium can be an effective biological control agent.
PDF Abstract XML References Citation
How to cite this article
M.A. Rahman, J. Kadir, T.M.M. Mahmud, R. Abdul Rahman and M.M. Begum, 2007. Screening of Antagonistic Bacteria for Biocontrol Activities on Colletotrichum gloeosporioides in Papaya
. Asian Journal of Plant Sciences, 6: 12-20.
DOI: 10.3923/ajps.2007.12.20
URL: https://scialert.net/abstract/?doi=ajps.2007.12.20
DOI: 10.3923/ajps.2007.12.20
URL: https://scialert.net/abstract/?doi=ajps.2007.12.20
INTRODUCTION
Papaya (Carica papaya L.) is one of the most popular tropical fruit in Malaysia and has an excellent potential as an export crop. In 2002, Malaysia occupied the top position for export papaya to Hong Kong and ranked second in the world among the papaya exporter countries after Mexico. Malaysia’s export of papaya was accounted for 28.5% of the total world exports in 2002 (FAMA, 2006). However, during storage and transportation, postharvest diseases may significantly lower the quality and value of this commodity (Gamagae et al., 2004). Among the postharvest diseases, anthracnose caused by Colletotrichum gloeosporioides Penz. Sacc., is a major postharvest disease in tropical countries (Paull et al., 1997; Snowdon, 1990). In Sri Lanka, the disease restricts the export of papaya to overseas markets (Gamagae et al., 2003). Anthracnose in papaya can be controlled by using postharvest application of prochloraz or propiconazole (Sepiah, 1993), hot water treatment (HWT) at 43-49°C for 20 min (Couey et al., 1984) and heat treatment or HWT in combination with fungicides (Couey and Farias, 1979). However, heat treatment leads to enhance softening of papaya fruit. The response of papaya fruit to heat treatments varies with maturity and season, with fruit being most susceptible to injury during cooler periods of storage (Paull, 1995). HWT affect the ripening process in papaya (Paull, 1990) and the use of fungicides for extended periods may cause emergence of strains of fungus resistance to these fungicides. The use of fungicides is becoming more restrictive due to health concerns (Ragsdale and Sisler, 1994) and consumers are demanding less chemical residue on produce. For these reasons, elucidating non-chemical control methods to reduce postharvest decay is becoming increasingly important. It is therefore, necessary to develop safer alternatives to synthetic fungicides that are effective and economically feasible as well as environmental friendly. An alternative that continues to show promising results is biological control of postharvest diseases (Zhou et al., 2001; Janisiewicz and Jeffers, 1997; Wilson and Wisniewski, 1989).
However, there has been no report on the successful biological control of Colletotrichum gloeosporioides, the causal agent of anthracnose disease of papaya using antagonistic bacteria. Therefore the objective of this study is to isolate, identify and evaluate bacterial antagonists against C. gloeosporioides.
MATERIALS AND METHODS
The experiment was conducted at Plant Protection Laboratory, Faculty of Agriculture, University Putra Malaysia, Serdang, Selangor, Malaysia in 2005.
Isolation of C. gloeosporioides: Colletotrichum gloeosporioides were isolated from infected papaya collected from different locations in Selangor, Malaysia. The isolation of C. gloeosporioides was according to procedures outlined by Bautista-Banos et al. (2003) and Gamagae et al. (2003). Identifications of the C. gloeosporioides were done by comparing its cultural and morphological characteristics with descriptions published in specialized literature (Sutton, 1992; Barnett and Hunter, 1972). The isolate was maintained on PDA slant at 28±2°C. Continuous re-inoculations and re-isolations on papaya fruit were carried out to maintain pathogenicity of the inoculum.
Preparation of conidial suspension of C. gloeosporioides: Isolate of C. gloeosporioides was grown on PDA agar at 28±2°C for 7 days. Spores were subsequently harvested by flooding the surface of the media with distilled water and gently agitating the plate with a bent glass rod to dislodge the spores. The resulting suspension was filtered through two layers of sterile muslin cloth. The concentration of conidia in the filtered suspension was adjusted to 5x105 conidia per mL with sterile distilled water using a haemacytometer (Sariah, 1994; Obagwu and Korsten, 2003).
Isolation of antagonistic bacteria from the surface of papaya fruit: Both infected and non-infected papaya fruits were used to isolate antagonistic bacteria. Pieces of fruit tissues, 5 mm2, were cut from the healthy and lesion area and then surface disinfected with 1% sodium hypochlorite. Fifty grams of sliced tissues are then soaked in 10 mL of sterilized water and rotary shaking for 24 h at room temperature (28±2°C). Fold serial dilutions (10-1-10-6) of suspension was made in test tube. Then 0.5 mL suspension from each test tube was spread on nutrient agar (NA; Difco, Detriot MI) medium with a sterile bent glass rod and incubated for 24-48 h at 28±2°C. Single colonies of bacteria were then transferred onto petri dishes containing NA to obtain pure culture (Dikin et al., 2002; Monteallegre et al., 2003).
Screening, selection and identification of bacteria: Seventy six isolates of bacteria were obtained by the dilution method from fructosphere of papaya collected from different locations in Selangor, Malaysia. These isolates were first dually cultured with C. gloeosporioides on PDA medium. Isolates with inhibitory characteristics against C. gloeosporioides were selected for further screening by means of dual and concomitant test (Wu and Chou, 1995).
The effective antagonistic bacteria were then identified by using BIOLOG (Biolog Inc., Hayward, CA, USA) identification system with the software Microstation System Release Version 4.20 as described by Chiou and Wu (2001). This test was repeated once.
Preparation of aqueous antagonist suspension: Isolates of Burkhoderia cepacia and Pseudomonas aeruginosa were selected based on their antagonistic activity. In preparing aqueous antagonist suspension, isolates were grown on NA at 28±2°C for 24 h. A loop of each culture was then transferred to a 250 mL conical flask containing 50 mL of nutrient broth (NB, Difco, Detroit, MI) and incubated on a rotary shaker at 150 rpm for 48 h at 28±2°C. The isolates were re-cultured in fresh nutrient broth for another 72 h before used (Smilanick et al., 1993). To enumerate, Colony Forming Units (CFU), cultures were serially diluted in 1% bacto-peptone (Difco, Detroit, MI) and plated on nutrient agar. The number of cfu counted after 48h incubation at 28±2°C. At the time of use, the suspension was adjusted to approximately 109 cfu mL-1.
Preparation of filter sterilized culture filtrate: Culture filtrates of both isolates were prepared using NB medium following the procedure as described by Yoshida et al. (2001) and used in this experiment to determine their effect on fungal growth.
Antagonism
Dual culture assay: For this test dual culture plates were prepared as described by Sariah (1994). Plates were incubated for seven days at room temperature (28±2°C).
After the incubation period, percent inhibition of radial growth (PIRG) was recorded based on the following formula (Sivakumar, 2002; Sariah, 1994):
Where,
| R1 | = | Radial growth of C. gloeosporioides in control plate |
| R2 | = | Radial growth of C. gloeosporioides interacting with antagonistic bacteria |
Furthermore, random plating of PDA plugs on fresh PDA plates taken from the interaction zone was done to assessed viability of the fungus in the zone of interaction in dual culture test. Each treatment was carried out with five replicates. The experiment was arranged according to completely randomized design.
Production of diffusible antifungal substances: In this experiment, diffusible antifungal substance (s) was produced in sandwich agar plates, which was prepared according to the procedure as described by Dikin et al. (2002). The experiment was conducted with five replications and repeated thrice. Results were expressed as means of %inhibition of growth of C. gloeosporioides in the presence and absence of bacterial isolate.
Production of volatile antibiotics: The ability of the bacterial isolate to produce volatile antibiotics was evaluated using published procedure as described by Montealegre et al. (2003). Results were expressed as means of % inhibition of growth of C. gloeosporioides in the presence and absence of bacterial isolate.
Mycelial growth test: Six mm agar plugs of C. gloeosporioides were taken by cork borer and placed into a sterilized glass test tube. Mycelial agar plugs were then dipped into selected antagonistic bacterial suspension (109 cfu mL-1) for 30 min and air dried. Treated mycelial plugs were transferred on petri dishes containing PDA medium. Fungal plugs dipped in sterile distilled water served as control. Each plate of a treatment was replicated 5 times. The plates were then incubated at 28±2°C for 7 days. The observation of radial inhibition of mycelial growth was carried out 7 days after incubation.
In another experiment, filter sterilized culture filtrate of B. cepacia and P. aeruginosa grown in nutrient broth (NB) were also tested for their antagonistic activity to C. gloeosporioides. The culture filtrates of each bacterium and a control of NB were mixed separately with PDA (2:3v/v) cooled to 45°C. Radial growth of the C. gloeosporioides on these amended media was recorded after 7 days of incubation at 28±2°C. Both of the experiments repeated twice.
Study on hyphal morphology: A 6 mm diameter mycelial plug from four days old PDA culture of C. gloeosporioides was transferred to the center of a petri dish, containing 15 mL of PD broth and incubated at 28±2°C for 2 days. When mycelial growth on the surface of the liquid reached a diameter about 1.5 cm, 1 mm bacterial suspension (109 cfu mL-1) of either B. cepacia or P. aeruginosa was transferred to the plate and the culture was further incubated at 28±2°C for 3 days. Hyphal strands at the end of the fungal colony were removed and examined under a microscope for abnormalities (Sariah, 1994). Cultures of C. gloeosporioides in PD broth without bacterial inoculation served as control. The experiment was arranged in a completely randomized design, with five replicates each. The experiment was repeated once.
Spore germination test: Two experiments were conducted to determine the effect of antagonistic bacteria on the spore germination of C. gloeosporioides. Bacterial suspensions (109 cfu mL-1) prepared as described above were used in this experiment. Spore suspension of C. gloeosporioides was also prepared with 5x105 spores/mL. Then 100 μL of cell suspension of selected bacteria were placed into a sterile test tube and added 100 μL of spore suspension of the test fungus. The mixed suspension was then incubated for 30 min at 28±2°C. For the control plate sterilized water was added instead of bacterial suspension. After completion of incubation time, 100 μL of mixed suspension was spread over the PDA plates with a sterile bent glass rod. The plates were then incubated for 24 h at room temperature (28±2°C). Spores were examined under a microscope for germination. A spore was considered to have germinated when the germ tube length was half of the length of the spore (Sariah, 1994). The experiment was repeated two times and plates were arranged in a completely randomized design with five replicates.
In the second experiment, filter-sterilized culture filtrates of selected bacteria were used instead of bacterial suspension to determine their effect on the spore germination and sterilized NB served as the control.
In another test, 100 μL of spore suspension of C. gloeosporioides was spread over the PDA plate with a sterile bent glass rod. Two sterilized filter paper discs (1 cm diameter), were placed 3 cm apart on the agar and 50 μL of bacterial suspension was pipetted onto each of the paper disc. In control plate, discs received 50 μL of sterilized NB only. After 24 h of incubation at 28±2°C, spore occurring within 10 mm around each disc were examined under a microscope for germination.
Statistical analysis: Data were statistically analyzed using SAS software (SAS, Institute Cary, NC, 1987), by analysis of variance and the significance of the treatments was determined using LSD (p<0.05). The percentage data were transformed into Arcsine transformation before subjected to ANOVA (Gomez and Gomez, 1984).
RESULTS
Isolation of antagonistic bacteria from the surface of papaya fruit: A total of 76 bacterial isolates were isolated based on liquid assay. Pure culture of each isolate was maintained for screening test against C. gloeosporioides.
Screening, selection and identification of antagonistic bacteria: Isolates with inhibitory characteristics were selected and screened by means of dual culture and concomitant tests. Of the 76 isolated bacteria, 27 were inhibitory to C. gloeosporioides on PDA (Table 1). Out of 27, four isolates namely B23, B19, B04 and B15 had a significantly (p<0.05) higher inhibitory effect than the others. The PIRG of B23, B19, B04 and B15 were 74.2, 74.06, 68.78 and 68.21%, respectively, with respect to the control after seven days of incubation ( Fig. 1).
Four selected antagonistic isolates were gram-negative, aerobic rod-shaped bacteria. Using the Biolog system, isolates B23 and B19 were identified as Burkholderia cepacia and B4 and B15 as Pseudomonas aeruginosa.
| Table 1: | Screening for antagonistic bacteria against Colletotrichum gloeosporioides of papaya. |
 | |
| Means in the column followed by same letter(s) are not different significantly at p<0.05 according to LSD.,* Percentage inhibition of radial growth of Colletotrichum gloeosporioides at 7 days after incubation | |
Both B. cepacia and P. aeruginosa strongly inhibited the mycelial growth of C. gloeosporioides on PDA medium. No mycelial growth was observed after four days of incubation, when PDA plugs from interaction zone were re-cultured on fresh PDA plates.
Production of diffusible and volatile antibiotics: Both B. cepacia and P. aruginosa were found to produce diffusible as well as volatile antibiotics, which showed significant difference with control (p<0.05) in respect of mycelial growth inhibition (Table 2). Mycelial growth of C. gloeosporioides was completely inhibited by diffusible antibiotics produced by both of the bacterial isolates after 7 days of incubation. B. cepacia showed the highest inhibitory effect on the growth of C. gloeosporioides by producing volatile antibiotics. Inhibition of mycelial growth was 26.6% at 7 days after incubation with respect to the control.
Mycelial growth test: Both B. cepacia and P. aeruginosa completely inhibited the mycelial growth of C. gloeosporioides on PDA medium after 7 days of incubation with respect to the control (Table 3). Filter sterilized culture filtrate of both bacteria also significantly reduced the growth of C. gloeosporioides.
| Table 2: | Growth inhibition of C. gloeosporioides by diffusible and volatile antibiotics produced by antagonistic bacteria at 7 days after incubation |
 | |
Means in a column with the same letter(s) do not different significantly at 0.05 levels according to LSD | |
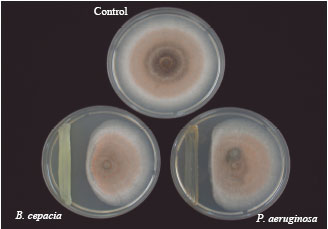 | |
| Fig. 1: | Inhibition of Colletotrichum gloeosporioides by B. cepacia and P. aeruginosa 7 days after incubation at 28±2°C |
| Table 3: | Effect of antagonistic bacteria on the growth and spore germination of C. gloeosporioides |
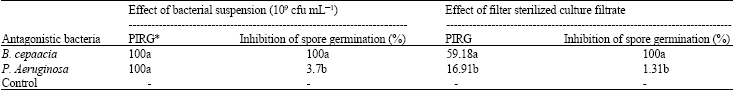 | |
| Means in a column with the same letter(s) do not different significantly at 0.05 levels according to LSD, *Percentage inhibition of radial growth of Colletotrichum gloeosporioides at 7 days after | |
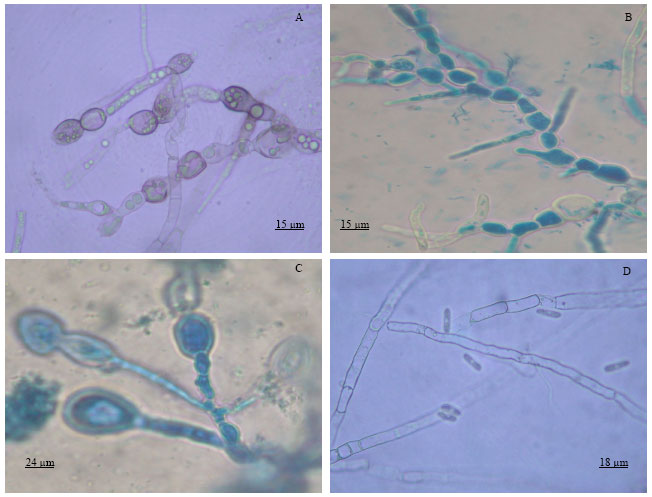 | |
| Fig. 2: | Hyphal morphology of C. gloeosporioides as affected by B. cepacia and P. aeruginosa. Occurrence of bubbles, vacuoles and swelling in hyphae (A); Thickened and swelled hyphae and hyphal tips (B-C) and D = Normal hyphae |
Inhibition of mycelial growth was recorded as 59.2 and 16.9% by the culture filtrate of B. cepacia and P. aeruginosa, respectively with respect to the control.
Study on hyphal morphology: A little additional mycelial growth of C. gloeosporioides was observed after introduction of both B. cepacia and P. aeruginosa to PDB. Hyphal tips of the fungus became malformed and hyphae were thickened and vacuolar compared with hyphae in the control plate. Many swellings occurred in the hyphae or at the tips of hyphal strands, whereas normal hyphal walls were smooth with no swellings or vacuolation ( Fig. 2).
Spore germination test: Spore of C. gloeosporioides did not germinate in the presence of cell suspension or the culture filtrate of B. cepacia after 24 h of incubation (Table 3). Whereas, only 3.7 and 1.31% of spore germination was inhibited in presence of cell suspension and culture filtrate of P. aeruginosa, respectively. In the control plates, 94.43% spore germination was recorded after seven h of incubation. The germinated spores were characteristics by the production of germ tubes and sometimes with the development of globose appressoria ( Fig. 3).
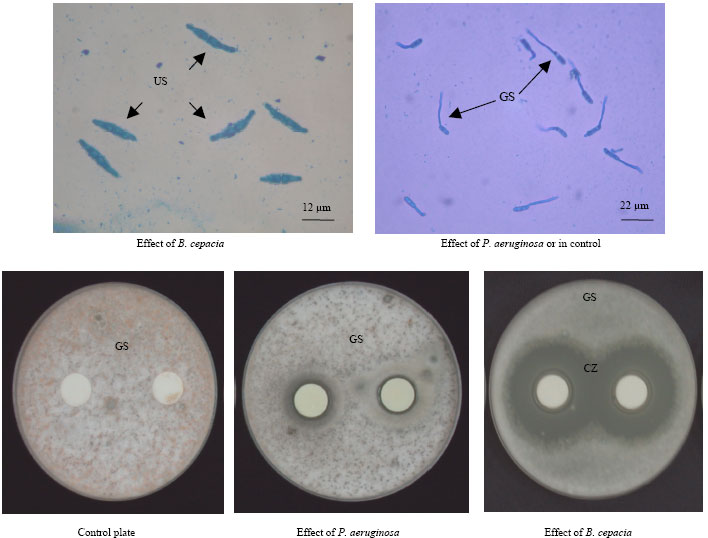 | |
| Fig. 3: | Un-germinated spores of C. gloeosporioides in the presence of B. cepacia (US); Germinated spores in the presence of P. aeruginosa or in control plate (GS); A clear zone (CZ) of un-germinated spores around the paper discs in the presence of B. cepacia |
DISCUSSION
Results of the present study clearly showed the potential of using a fructosphere-associated antagonistic bacterium to control anthracnose disease of papaya, caused by Colletotrichum gloeosporioides. These bacterial isolates were identified as Burkholderia cepacia and Pseudomonas aeruginosa by the BIOLOG system. The most efficacious biocontrol agent identified in this present research was B. cepacia. It is a gram-negative, nonfluorescent pseudomonad of extreme genetic diversity (Ramette et al., 2005; Schroth et al., 1983). It is a ubiquitous, primarily soil-inhabiting organism with little tolerance to desiccation that can only survive in low populations on leaf surfaces (Armstrong et al., 1987; Leben et al., 1965). Many members of B. cepacia complex have been isolated from environmental and plant samples. Among them, candidates suitable for biopesticide action, plant growth promotion and bioremediation have been found (Parke and Gurian-Sherman, 2001). Moreover, isolates similar to B. cepacia genomovar III, have been found as a plant associated bacterium from soil and inner tissues of crop plants such as maize, wheat and lupin (Balandreau et al., 2001). This indicates the close association between B. cepacia and crop plant surfaces. Furthermore, commercial products containing B. cepacia are available in the market for control of many soil-borne pathogens (Fravel et al., 1998).
The results of present study demonstrated that the isolate of B. cepacia produced antibiotic substances in vitro that inhibited mycelial growth and spore germination of C. gloeosporioides. Mycelial malformation observed was probably due to the toxic effect of antibiotic substances interfering with normal growth processes (Sariah, 1994). B. cepacia is known to produce a wide range of secondary metabolites such as pyrrolnitrin, phenazine and cepabactin, which can influence the growth and morphology of fungal mycelium (Cartwright et al., 1995; Meyer et al., 1989). Pyrrolnitrin is a broad-spectrum antifungal antibiotic. Antibiotic production by P. cepacia has been suggested to be the mode of disease control (Homma et al., 1989; Janisiewicz and Roitman, 1988). The inhibition of radial growth of mycelium in this study is considered to be antibiosis, in which the antibiotic metabolites may be penetrate in cell and inhibited its activity by chemical toxicity. The vacuolar appearance of the mycelium also probably may be due to the antibiotic metabolites produced by the bacteria, which may penetrate and caused protoplasmic dissolution and disintegration (Huber et al., 1966).
The failure of spores of C. gloeosporioides to germinate after 24 h exposure either to the bacterial cell suspension or culture filtrate of B. cepacia indicated that the antibiotic metabolite substances produced by the bacterium was not only fungistatic but also fungicidal to the spore of the test fungus. Suppression of spore germination of several fungi by B. cepacia has been reported (De Costa and Erabadupitiya, 2005; Smilanick and Denis-Arrue, 1992; Janisiewicz and Roitman, 1988).
In present study we also found that both B. cepacia and P. aeruginosa produced volatile substance as well as diffusible substances that significantly inhibited the mycelial growth of C. gloeosporioides on PDA. Many workers reported that the production of volatile ammonia by Pseudomonas spp. has been indicated as a possible mechanism to control soilborne pathogens (Baligh et al., 1996, 1991; Conway et al., 1989).
Therefore, in vitro activities of the B. cepacia against C. gloeosporioides of papaya in this study suggested that the bacterium can be an effective biological control agent. The use of B. cepacia as a biocontrol agent against C. gloeosporioides of papaya may be an economical as well as environmental safe way to suppress the disease. However, the form in which the bacterium can be applied on fruits needed further investigation.
ACKNOWLEDGMENT
The authors wish to thank the Government of Malaysia for financing this research through IRPA Grant 01-02-04-0061 EA001.
REFERENCES
- Armstrong, J.L., G.R. Knudsen and R.J. Seidler, 1987. Microcosm method to assess survival of recombinant bacteria associated with plants and herbivorous insects. Curr. Microbiol., 15: 229-232.
CrossRef - Balandreau, J., V. Viallard, B. Cournoyer, T. Conenye, S. Laevena and P. Vandamme, 2001. Burkholderia cepacia genomovar III is a common plant associated bacterium. Applied Environ. Microbiol., 67: 982-985.
Direct Link - Bautista-Banos, S., M. Hernandez-Lopez, E. Bosquez-Molina and C.L. Wilson, 2003. Effects of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose levels and quality of papaya fruit. Crop Protect., 22: 1087-1092.
CrossRefDirect Link - Cartwright, D.K., C. Chilton and D.M. Benson, 1995. Pyrrolnitrin and phenazine production by Pseudomonas cepacia, strain 5.5B, a biological agent of Rhizoctonia solani. Applied Microbiol. Biotechnol, 43: 211-216.
Direct Link - Chiou, A.L. and W.S. Wu, 2001. Isolation, identification and evaluation of bacterial antagonists against Botrytis elliptica on Lily. J. Phytopathol., 149: 319-324.
Direct Link - Couey, H.M., A.M. Alvarez and M.G. Nelson, 1984. Comparison of hot water spray and immersion treatment for control of postharvest decay of papaya. Plant Dis., 68: 436-437.
Direct Link - Costa, D.M. and H.R.U.T. Erabadupitiya, 2005. An integrated method to control postharvest diseases of banana using a member of the Burkholderia cepacia complex. Posthar. Biol. Tec., 36: 31-39.
CrossRef - Dikin, A., M.A. Sijam, Z. Abidin, H.M. Zakaria and A.S. Idris, 2002. Interaction between Schizophyllum commune and antagonistic bacteria isolated from rotted fruits of oil palm. Proceedings of 25th Malaysian Microbiology Symposium and 5th UNESCO National Workshop for the Promotion of Microbiology in Malaysia, Towards Commercialization of Microbiology Research, Sept. 8-9, Kota Bahru, Kelantan Darul Naim, Malaysia.
- Gamagae, S.U., D. Sivakumar, R.S.W. Wijeratnam and R.L.C. Wijesundera, 2003. Use of sodium bicarbonate and Candida oleophila to control anthracnose in papaya during storage. Crop Prot., 22: 775-779.
CrossRefDirect Link - Gamagae, S.U., D. Sivakumar and Wijesundera, 2004. Evaluation of post-harvest application of sodium bicarbonate-incorporated wax formulation and Candida oleophila for the control of anthracnose of papaya. Crop Prot., 23: 575-579.
Direct Link - Yoshihisa, H., S. Zenji, H. Fukushi, K. Katsuhiro, S. Haruhisa and S. Takahito, 1989. Production of antibiotics by Pseudomonas cepacia as an agent for biological control of soilborne Plant pathogens. Soil Biol. Biochem., 21: 723-728.
CrossRef - Huber, D.M. A.L. Andersen and A.M. Finley, 1966. Mechanisms of biological control in a bean root rot soil. Phytopathology, 56: 953-956.
Direct Link - Janisiewicz, W.J. and J. Roitman, 1988. Biological control of blue mold and gray mold on apple and pear with Pseudomonas cepacia. Phytopathology, 78: 1697-1700.
Direct Link - Janisiewicz, W.J. and S.N. Jeffers, 1997. Efficacy of commercial formulation of two biofungicides for control of blue mould and gray mould of apples in cold storage. Crop Prot., 16: 629-633.
Direct Link - Meyer, J.M., D. Hohnadel and F. Halle, 1989. Cepabactin from Pseudomonas cepacia, a new type of siderophore. J. Gen. Microbiol., 135: 1479-1487.
PubMed - Obagwu, J. and L. Korsten, 2003. Integrated control of citrus green and blue molds using Bacillus subtilis in combination with sodium bicarbonate or hot water. Postharvest Biol. Technol., 28: 187-194.
CrossRefDirect Link - Parke, J.L. and D. Gurian-Sherman, 2001. Diversity of the Burkholderia cepacia complex and implications for risk assessment of biological control strains. Ann. Rev. Phytopathol., 39: 225-258.
Direct Link - Paull, R.E., 1995. Preharvest factors and the heat sensitivity of field-grown ripening papaya fruit. Postharvest Biol. Technol., 6: 167-175.
CrossRefDirect Link - Ragsdale, N.N. and H.D. Sisler, 1994. Social and political implications of managing plant diseases with decreased availability of fungicides in the United States. Ann. Rev. Phytopathol., 32: 545-557.
PubMed - Ramette, A., J.J. LiPuma and J.M. Tiedje, 2005. Species abundance and diversity of Burkholderia cepacia complex in the environment. Applied Environ. Microbiol., 71: 1193-1201.
Direct Link - Sariah, M., 1994. Potential of Bacillus spp. as a biocontrol agent for anthracnose fruit rot of chilli. Malays. Applied Biol., 23: 53-60.
Direct Link - Smilanick, J.L. and R. Dennis-Arrue, 1992. Control of green mold of lemons with Pseudomonas species. Plant Dis., 76: 481-485.
Direct Link - Smilanick, J.L., R. Denis-Arrue, J.R. Bosch, A.R. Gonzalez, D. Henson and W.J. Janisiewicz, 1993. Control of postharvest brown rot of nectarines and peaches by Pseudomonas species. Crop Prot., 12: 513-520.
Direct Link - Wilson, C.L. and M.E. Wisniewski, 1989. Biological control of postharvest diseases of fruits and vegetables: An emerging technology. Annu. Rev. Phytopathol., 27: 425-441.
CrossRefDirect Link - Wu, W.S. and J.K. Chou, 1995. Chemical and biological control of Alternaria carthami on zinnia. Seed Sci. Technol., 23: 193-200.
Direct Link - Yoshida, S., S. Hiradate, T. Tsukamoto, K. Hatakeda and A. Shirata, 2001. Antimicrobial activity of culture filtrate of Bacillus amyloliquefaciens RC-2 isolated from mulberry leaves. Phytopathology, 91: 181-187.
CrossRefDirect Link - Zhou, T., C.L. Chu, W.T. Liu and K.E. Schaneider, 2001. Postharvest control of blue mould and gray mould on apples using isolates of Pseudomonas syringae. Can. J. Plant Pathol., 23: 246-252.
Direct Link