
W.M. Muiru
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, Nairobi, Kenya
E.W. Mutitu
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, Nairobi, Kenya
J.W. Kimenju
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, Nairobi, Kenya
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 8 | Page No.: 1190-1196







ABSTRACT
Fifty seven maize lines comprising 23 open pollinated, 30 hybrids and 4 inbreed lines were evaluated for resistance to Turcicum leaf blight both under greenhouse and field conditions. For greenhouse evaluation, the genotypes were inoculated with mixed inoculum derived from seven pathogenic isolates of Exserohilum turcicum whereas in the field trial, natural inoculum was relied upon. The parameters assessed were incubation period, disease incidence, disease severity, lesion density and grain yield. The scale used for disease severity in the greenhouse ranged from 0-6 whereas in the field evaluation, assessment was done using a 0-5 scale based on the proportionate leaf area affected by the disease. The maize genotypes evaluated showed a great variation in terms of lesion density, disease incidence and disease severity. Four lines namely SC Duma 41, H614D, H625, HR-KIB-04A-1-5 consistently rated as resistant with disease severity scores of less than 1, while IR-KIB-03B-19-4, PH1, IR-KIB-03B-19-2, PH4, ZIMLINE/KAT BG-25 and SYNIRO 1 showed a high level of susceptibility with severity scores of higher than 3. The maize genotypes were categorized into three groups namely tolerant, moderately tolerant and susceptible. Fifty three percent of the lines were moderately tolerant while 17 and 30% were tolerant and susceptible, respectively. The susceptible lines produced elliptical gray necrotic sporulating lesions while the lesions were chlorotic and small to medium in size in tolerant genotypes. This study is a clear demonstration of the existence of levels of tolerance in maize genotypes which can be utilized in the management of northern leaf blight.
PDF Abstract XML References Citation
How to cite this article
W.M. Muiru, E.W. Mutitu and J.W. Kimenju, 2007. Reaction of Some Kenyan Maize Genotypes to Turcicum Leaf Blight Under Greenhouse and Field Conditions. Asian Journal of Plant Sciences, 6: 1190-1196.
DOI: 10.3923/ajps.2007.1190.1196
URL: https://scialert.net/abstract/?doi=ajps.2007.1190.1196
DOI: 10.3923/ajps.2007.1190.1196
URL: https://scialert.net/abstract/?doi=ajps.2007.1190.1196
INTRODUCTION
Turcium leaf blight of maize (Zea mays) also known as northern corn leaf blight caused by Exserohilum turcicum (Pass.) teleomorph Setosphaeria turcica (Lutrell Leonard and Suggs) is a widespread disease of maize which can cause yield losses of up to 70% (Yeshitila, 2003). Apart from yield loss, the disease causes qualitative changes in the seed resulting to decreased sugar content, germination capacity and severely infected plants are predisposed to stalk rot (Gowda et al., 1992; Cardwell et al., 1997). Although the disease has a worldwide distribution, it is more severe in cool humid regions characterized by heavy dew (Dorothea et al., 1998; Juliana et al., 2005).
The disease is mainly controlled through the use of resistant varieties derived from qualitative and quantitative genes acting together or separately (Juliana et al., 2005). Qualitative resistance is typically race-specific and controlled by single genes whereas quantitative resistance is race-non specific and controlled by many genes (Welz and Geiger, 2000; Singh et al., 2004). Most of the qualitative genes such as Ht1, Ht2, Ht3, Htn and Htm are dominant or partially dominant and have a great phenotypic effect but might be overcome by virulent genes present in specific races of the pathogen (Mwangi, 1998; Juliana et al., 2005). Disease severity, disease incidence, lesion density, lesion sizes and area under disease progress curve are the common parameters used in the evaluation of maize germplasm to turcicum leaf blight (Adipala, 1994).
Resistance conferred by Ht genes is characterized by chlorotic and necrotic lesions without spore formation thereby limiting the spread of the disease Welz and Geiger, 2000; Singh et al., 2004). Resistance conferred by minor genes is not absolute as in qualitative resistance but chances of new biotypes overcoming this resistance are considered to be minimal (Ojulong et al., 1995). Current breeding programmes rely predominantly on qualitative resistance conferred by Ht genes but emergence of new races of the pathogen is a constant threat (Freymark et al., 1994; Juliana et al., 2005). E. turcicum exhibits a wide pathogenic variability (Yeshitila, 2003) and reports of new races overcoming previously resistant varieties have been documented. Although many maize genotypes have been released from breeding programmes, their reaction to the northern leaf blight pathogen remains largely unknown. This study was undertaken with the aim of determining the reaction of 57 maize genotypes to northern leaf blight pathogen under greenhouse and field conditions.
MATERIALS AND METHODS
The study was carried out at the University of Nairobi field station during the months of March to August 2006 and the repeat trials done between September 2006 to February 2007. Fifty seven maize lines comprising 23 open pollinated, 30 hybrids and 4 inbred lines were obtained from CIMMYT and from the local seed stockists. The genotypes were evaluated for resistance to northern corn leaf blight of maize under field and greenhouse conditions.
Inoculum preparation: Isolates of E. turcicum were obtained from diseased maize samples collected from different agro-ecological zones representing the main maize growing areas in Kenya. The infected leaves were cut into small sections (1 square centimetre) and surface sterilized using 2.5% Sodium hypochlorite for 2-3 min. The tissues were then rinsed with sterile distilled water, blot dried and plated on corn meal agar in petri-dishes and incubated at room temperature for 3-4 days. Pure cultures were prepared by sub-culturing from the isolation plates and incubating the cultures for 7-10 days to obtain sufficient growth. Inoculum was prepared by flooding the cultures with sterile distilled water, scrapping the surface with microscopic slides to dislodge the conidia and then filtering using cheese cloth. The concentration of the conidia suspension was then adjusted to 105 conidia per mL using a haemocytometer. Tween 20 (Polyoxythylene sorbitan monolaurate) was added at the rate of two drops per 10 mL of water.
Greenhouse evaluation: The potting medium was prepared by mixing soil, manure and sand at the ratio of 6:2:1, respectively and sterilized by autoclaving. The potting medium was put into 20 cm diameter polythene sleeves. Diamonium phosphate (DAP) fertilizer was added at the rate of 10 g per pot and maize seeds sowed into the potted media. Two seeds per genotype were sowed into each pot and the experimental plot comprised of 9 pots replicated three times. The treatments were arranged in a completely randomized design and watering was done lightly until the emergence of maize seedlings. Subsequent watering was done depending on the moisture status of the potting medium and this was maintained moist to feel in all the pots.
Inoculum was prepared from 7 isolates as described above and used to inoculate the maize seedlings. Inoculation was done at the fifth leaf stage and this was accomplished by spraying the maize plants with the conidia suspension until runoff using a hand atomizer. The maize seedlings were then covered with clear polythene bags moistened with sterile distilled water for 24 h to raise the relative humidity necessary for the infection to take place. The ground and other open surfaces were sprinkled with water to increase the relative humidity and lower the temperatures in the greenhouse.
Inoculated plants were monitored daily and parameters such as type of symptoms, incubation period, lesion sizes, disease incidence and disease severity recorded for each of the lines. The reaction was assessed based on a modified scale from Bigirwa et al. (1993), where 0 = no symptoms, 1 = chlorotic lesions with a few (1-2) lesions, 2 = chlorotic lesions with some necrosis and 3-5 lesions, 3 = moderate (6-9) lesions with chlorotic lesions and considerable necrosis, 4 = heavy infection (over 10 lesions) with wilting of some leaves, 5 = coalesced lesions and generalized wilting of the whole plant and 6 = plant completely dead.
Field evaluation: The field plots were established at Kabete Campus, University of Nairobi and the field was prepared by disc ploughing and harrowing to obtain a fine tilth. The different maize genotypes were planted and planting depth was maintained at two and a half times the length of the seed and DAP at the rate of 10 g hill-1 added. The trials were laid in a randomized complete block design. The rows comprised the experimental plots and these were replicated four times. Each row had 20 hills and the distance between rows was 75 cm whereas the distance between the hills was 30 cm. Rows and blocks were spaced 1 and 2 m, respectively. Two seeds were sowed per hill and thinning done later when the maize seedlings had attained a height of 15 cm.
Unlike in the greenhouse evaluation, the source of inoculum was natural infection. Watering was done to field capacity using overhead irrigation to promote conditions that are favourable for disease development. Top dressing was done using Calcium Ammonium Nitrate (CAN) at the rate of 10 g hill-1 when the plants were about 40 cm in height. The disease was monitored continuously until the crop reached physiological maturity.
The disease parameters assessed and scored were lesion density, lesion sizes, disease severity and yield data. The key used for scoring disease severity was a modification from Elliot and Jenkins (1946):
where:
| 0 | = | No symptoms, |
| 0.5 | = | Very slight infection one or two restricted lesions on lower leaves, |
| 1 | = | Slight infection, a few scattered lesions (3-8) on lower leaves, |
| 2 | = | Light infection, moderate number of lesions (9-15) on lower leaves, |
| 3 | = | Moderate infection, abundant lesions (>16) on lower leaves and a few on middle leaves, |
| 4 | = | Heavy infection, lesion abundant on lower and middle leaves and extending to the upper leaves, |
| 5 | = | Very heavy infection, lesions abundant on all leaves and plants may be killed prematurely. |
Yield data was taken at physiological maturity by picking 10 cobs per line, hand shelling and weighing. Genstat ver 8.0 was used to analyse the data and ANOVA was performed to show the significance of differences in the parameters tested. The means were separated using least significant difference.
RESULTS
The maize genotypes showed a significant variation in percentage of diseased leaf tissue and the lesion density (Table 1, 2). The incubation period was 4-5 days with no significant differences among the maize genotypes evaluated. Small pin-head size spots were observed on all the maize genotypes 48 h after inoculation. Initial symptoms appeared as small chlorotic flecks, which later enlarged and coalesced to form large chlorotic lesions. Susceptible genotypes produced small necrotic lesions which tended to enlarge and coalesce whereas in tolerant genotypes, the necrotic lesions remained localized. The characteristic Turcicum leaf blight symptoms of elliptical gray necrotic sporulating lesions were observed 7-10 days after inoculation and these lesions coalesced with time resulting to wilting of the leaves in most of the susceptible genotypes.
Tolerant genotypes formed chlorotic spots whereas in the susceptible ones, the small pin head spots developed into typical lesions. The genotypes which developed 3-9 small chlorotic lesions were considered moderately resistant, whereas those that developed more than 10 lesions characterized by considerable necrosis and wilting were considered susceptible.
| Table 1: | Disease severity, lesion sizes and type of symptoms on the various maize genotypes under green house conditions |
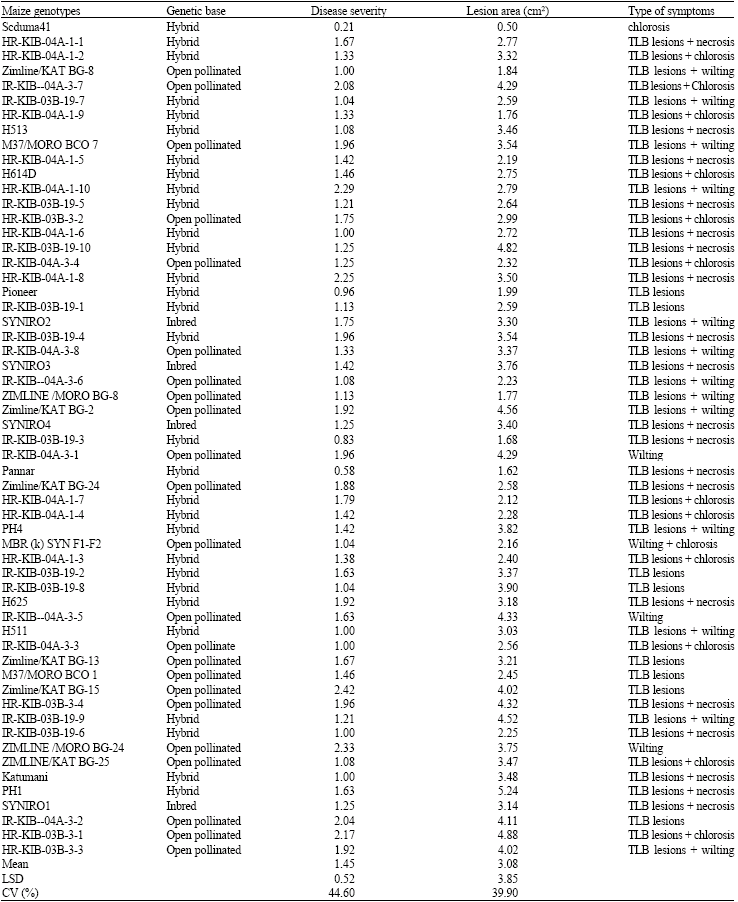 | |
| TLB = Turcicum Leaf Blight | |
| Table 2: | Area under disease progress curve, lesion density, lesion sizes, disease severity and yields from the various genotypes under field evaluation |
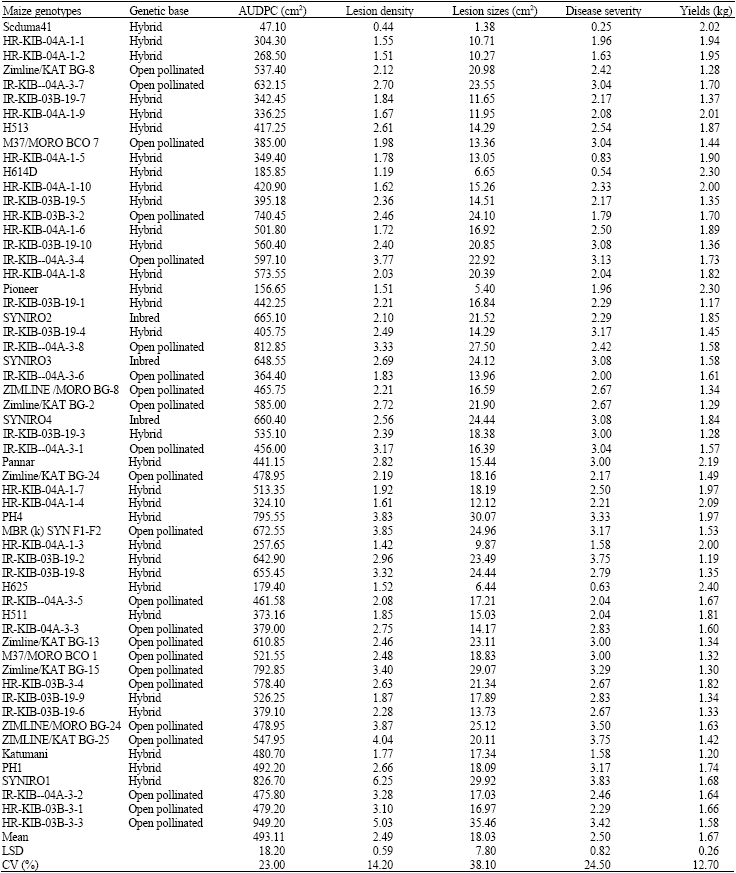 | |
| AUDPC is area under disease progress curve | |
Water-soaked lesions with dark margins and profuse sporulation developed on susceptible plants. Disease severity, lesion sizes, area under disease progress curve and lesion densities varied with the maize genotypes and reactions among the genotypes were consistent both under the greenhouse and field trials. Maize genotype SC Nduma had small (1.38 cm2) lesions and was rated as the most tolerant whereas HR-KIB-03B-3-3 was found to be highly susceptible with large lesions (35.46 cm2) and extensive chlorosis.
Out of the 57 genotypes assessed for reaction to northern leaf blight, 10 or 17% were found to be tolerant, 30 genotypes or 53% were found to be moderately tolerant, while 17 genotypes or 30% were found to be susceptible. The differences in disease severity were significant among the maize genotypes. Reactions to the disease ranged from tolerant (severity scores of 0 and 1) to highly susceptible (score 4-5) with some varieties showing moderate levels of tolerance (scores 2-3). Of the ten genotypes found to be tolerant, 9 were hybrids while 1 was an open pollinated variety. Of the 30 genotypes found to have moderate levels of tolerance 16 were hybrids, 13 were open pollinated varieties and 1 was an inbred line. Of the 17 genotypes found to be susceptible to northern leaf blight 5 were hybrids, 9 were open pollinated whereas 3 were inbred lines.
DISCUSSION
The effect of the disease was more severe in the greenhouse plants where apart from necrosis and chlorosis, wilting and death of the seedlings was quite common. This agrees with report by Agrios (2005) and this phenomenon can be explained by the fact that, young plants are normally more susceptible compared to older plants. Inoculation in the greenhouse had been done when the maize plants were 30 days old as opposed to field evaluation where natural inoculum was relied upon and the disease set in much later around the tasseling stage. Cumulatively, lesion sizes were much higher in the field evaluation since the plants were allowed to grow to maturity thereby offering more area for infection as opposed to greenhouse evaluation where the trials were terminated three months after inoculation.
Of the parameters used in the germplasm evaluation, counting lesions to determine lesion density and measuring lesion sizes was found to be tedious and cumbersome especially when dealing with large number of genotypes. Similar findings were reported by Adipala (1994) who suggested use of disease severity index as it is reliable and easy to score. The study identified a number of genotypes with appreciable levels of tolerance to Turcicum leaf blight. It is not possible to ascertain the reaction of theses genotypes in other localities with different climatic conditions since environmental factors such as temperature and light can modify resistance based on Ht genes and also disease pressure varies in different areas. Welz and Geiger (2000) reported differential expression of resistance by some varieties when tested at different places. There is therefore a need to test such varieties in different localities and including as many races of the pathogen to ensure that such varieties possess a durable resistance since E. turcicum is a highly variable pathogen. This will also give a better assessment of the reaction of such genotypes to northern leaf blight.
Majority of the genotypes were found to be moderately tolerant and susceptible to Turcicum leaf blight. Since E. turcicum is a highly variable pathogen, more sources of resistance need to be sought not only from other corn varieties but also from other close relatives. Welz et al. (1998) reported the existence of resistance genes in tripsacum and this has been introgressed into the corn plant to improve resistance. Since use of resistance varieties is the most feasible and practical measure of managing Turcicum leaf blight of maize, there is a need to intensify breeding programmes and incorporate both qualitative and quantitative resistance. Qualitative resistance controlled by Ht1, Ht2, Ht3 and Htn genes is limited in practical value because it is unstable and thus not durable (Welz, 1998). Quantitative resistance is insensitive to varying light and temperature conditions thus it is more stable and durable (Carson and Dyke, 1994).
With the use of host differentials bearing Ht genes, it is possible to characterize the lesion type into chlorotic lesions as was observed with inbreds bearing Ht genes and necrotic lesions on susceptible genotypes without Ht genes. In this study, most genotypes had a combination of chlorotic and necrotic lesions and this is evident that such genotypes had a combination of partial and complete resistance and this is corroborated by findings of Lipps et al. (1997). Although the number of inbreds included in the study were few, they were found to be more susceptible to Turcicum leaf blight compared to hybrids and open pollinated varieties and these findings are in agreements with those of Adipala et al. (1993). Differences among the genotypes were well represented by disease incidence, severity, lesion density and lesion area.
ACKNOWLEDGMENT
This research was funded by the German Academic Exchange Programme (DAAD).
REFERENCES
- Adipala, E., P.E. Lipps and L.V. Madden, 1993Adipala, E., P.E. Lipps and L.V. Madden, 1993. Reaction of maize cultivars from Uganda to Exserohilum turcicum. Phytopathology, 83: 217-233.
Direct Link - Agrios, G.N., 2005. Plant Pathology. 5th Edn., Elsevier, Cambridge, Massachusetts, ISBN-13: 9780080473789, Pages: 922.
Direct Link - Bigirwa, G., A.M. Julian and E. Adipala, 1993. Characterization of Ugandan isolates of E. turcicum from maize. Afr. Crop Sci. J., 1: 59-67.
Direct Link - Cardwell, K.F., F. Schulthess, R. Ndemah and Z. Ngoko, 1997. A systems approach to assess crop health and maize yield losses due to pests and diseases in Cameroon. Agric. Ecosyst. Environ., 65: 33-47.
CrossRefDirect Link - Carson, M.L. and C.G. Van Dyke, 1994. Effect of light and temperature on expression of partial resistance of maize to Exserohilum turcicum. Plant Dis., 78: 519-522.
Direct Link - Dorothea, S.B., H.W. Hunter and H.G. Hartwig, 1998. Molecular marker analysis of European Setosphaeria turcica populations. Eur. J. Plant Pathol., 104: 611-617.
Direct Link - Freymark, P.J., M. Lee, C.A. Martinson and W.L. Woodman, 1994. Molecular marker facilitated investigation of host plant response to E. turcicum in maize (Zea mays L.). Theor. Applied Genet., 88: 305-313.
Direct Link - Gowda, K.T.P., H.S. Shetty, B.J. Gowda, H.S. Prakash and L. Sangam, 1992. Comparison of two methods for assessment of yield loss due to Turcicum leaf blight of maize. Indian Phytopathol., 45: 316-320.
Direct Link - Juliana, B.O., A.G. Marco, O.G. Isaias and E.A.C. Luis, 2005. New resistance gene in Zea mays Exserohilum turcicum pathosystem. Genet. Mol. Biol., 28: 435-439.
Direct Link - Lipps, P.E., R.C. Pratt and J.J. Hakiza, 1997. Interaction of Ht and partial resistance to Exserohilum turcicum in maize. Plant Dis., 81: 277-282.
Direct Link - Ojulong, H.F., E. Adipala and P.R. Rubaihayo, 1995. Effects of concentrating resistance to Northern leaf blight of maize on agronomic traits. East. Afr. Agric. For. J., 61: 161-166.
Direct Link - Singh, R., V.P. Mani, K.S. Koranga, G.S. Bisht, R.S. Khandelwal, P. Bhandari and S.K. Pan, 2004. Identification of additional sources of resistance to Exserohilum turcicum in maize (Zea mays L.) Sabrao. J. Breed. Genet., 36: 45-47.
Direct Link - Welz, H.G., T. Lubberstedt, H.H. Geiger and A.E. Melchinger, 1998. Resistance of early maturing European maize inbred lines to Setosphaeria turcica. Maydica, 43: 27-33.
Direct Link - Welz, H.G. and H.H. Geiger, 2000. Genes for resistance to Northern corn leaf blight in diverse maize populations. Plant Breed., 119: 1-14.
Direct Link