
M. Kafi
Department of Agronomy, Ferdowsi University, P.O. Box 1163, Mashhad, Iran
H. Griffiths
Physiological Ecology Group, Department of Plant Science, University of Cambridge, UK
A. Nezami
Department of Agronomy, Ferdowsi University, P.O. Box 1163, Mashhad, Iran
H.R. Kazaie
Department of Agronomy, Ferdowsi University, P.O. Box 1163, Mashhad, Iran
A. Sharif
Department of Food Science, Ferdowsi University, P.O. Box 1163, Mashhad, Iran
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 8 | Page No.: 1166-1173







ABSTRACT
This study carried out in order to investigate the effects of salinity and time after salinisation on carbon isotope discrimination (Δ) of shoots and grains of three spring wheat cultivars (cv). Plants were grown in sand culture medium at four levels of salinity including 0 (control), 100, 200 and 300 mol m-3 of NaCl and CaCl2 from two leaves stage until ripening under controlled environment conditions. Carbon isotope discrimination decreased with increasing salinity but became significant only at the highest level of salinity. In all cvs ratio of intercellular to ambient partial pressure of CO2 (pi/pa) increased with increasing Δ. Although, the regression of Δ and pi/pa is positive, the slope of regression line is different from that predicted by theory, indicating an underestimation of Δ in these samples. Significant differences of Δ between shoot and grains were observed in all cvs and all levels of salinity. Relatively more 13C (lower Δ) was found in grains as compared with shoots. Therefore, Δ might be applicable, as a useful tool, for study of salinity tolerance of wheat genotypes.
PDF Abstract XML References Citation
How to cite this article
M. Kafi, H. Griffiths, A. Nezami, H.R. Kazaie and A. Sharif, 2007. Effects of Salinity on Carbon Isotope Discrimination of Shoot and Grain of Salt-Tolerant and Salt-Sensitive Wheat Cultivars. Asian Journal of Plant Sciences, 6: 1166-1173.
DOI: 10.3923/ajps.2007.1166.1173
URL: https://scialert.net/abstract/?doi=ajps.2007.1166.1173
DOI: 10.3923/ajps.2007.1166.1173
URL: https://scialert.net/abstract/?doi=ajps.2007.1166.1173
INTRODUCTION
The rate of photosynthesis is generally reduced by salinity. This reduction could be partly due to a reduced stomatal conductance and consequently restriction of the availability of CO2 for carboxylation. During the fixation of carbon by photosynthesis, the naturally occurring stable isotope 13C is discriminated against, because of fractionation of carbon stable isotopes (12C and 13C) mainly by Rubisco (Farquhar et al., 1989). Plants therefore, contain a lower ratio of 13C to 12C than the air that supplies them (Farquhar and Richards, 1984).
The most general model describing carbon isotope fractionation during photosynthesis in C3 plants assumes that the major components contributing to the overall fractionation are the differential diffusion of CO2 containing 12C and 13C across the stomatal pathway and the fractionation by rubisco. Farquhar et al. (1982b) reported that when stomatal conductance is small in relation to the capacity for CO2 fixation, intercellular partial pressure of CO2 (pi) is also small and carbon isotope discrimination (Δ) tends toward 4.4% (α). When conductance is comparatively large, pi approaches atmospheric CO2 partial pressure (pa) and Δ approaches net fractionation (b) between 27 to 30 ‰. Farquhar et al. (1982b) suggested the following expression for Δ in leaves of C3 plants:
(1) |
Farquhar et al. (1982a) suggested that there is a significant correlation between Δ in plant dry matter and pi/pa among different species over the maximum range of pi/pa 0.30-0.85. They plotted the carbon isotopes composition ![]() and pi/pa for Phaseolus vulgaris and Avicennia marina and found reasonable agreement between
and pi/pa for Phaseolus vulgaris and Avicennia marina and found reasonable agreement between ![]() and Ci/Ca.
and Ci/Ca.
Guy et al. (1986) found that increased salinity decreased Δ in Puccinellia maritima and pi/pa was low as expected from theory. This discrimination will be least in those plants that have the greatest water use efficiency. A linear increase in soil salinity produced significant (p<0.05) and linear decrease in grain yield and Δ, suggesting that Δ is a useful indicator of yield potential in barley (Isla et al., 1998; Qingzhen et al., 2006). Quarrie et al. (1999) postulated that Δ is an integral measure of plant water-use efficiency. Selecting for low Δ has also resulted in increased yield under drought conditions.
Farquhar et al. (1989) demonstrated that because of partial stomatal closure due to salinity, pi/pa might decrease with increasing stress. Therefore, based on Equation 1, the value of Δ decreases with increasing salinity. Brugnoli and Lauteri (1990), reported that in both a salt tolerant (Gossypium hirsutum L.) and a salt-sensitive species (Phaseolus vulgaris L.) Δ decreased with increasing salinity, however, Δ showed a more pronounced salt-induced decline in Phaseolus than Gossypium. The results obtained by Merah et al. (1999) confirm the validity of kernel Δ as a predictive criterion for grain yield under water stress in durum wheat and suggest the possible use of kernel ash content as an alternative criterion to select genotypes with higher water stress tolerance
The genetic control of Δ appears to be strong in wheat (Farquhar et al., 1989; Robina and Hood-Nowotny, 2005). Voltas et al. (1999) reported that genotypic Δ were more consistent across environment than the genotype yield of barley cvs. Condon et al. (1987) showed that the genotypic ranking was maintained at different sites and between wheat plants grown in pots and in the field.
Assessment of the possibility of using carbon isotope discrimination as an alternative method for selecting wheat material might be time saving and beneficial. Therefore, the aims of the present study were to investigate the effects of salinity and time after salinisation on Δ of shoots and grains of three wheat cvs showing marked differences in sensitivity to salinity from four hours to four weeks after salinisation and at ripening. Relationships between stomatal conductance, photosynthesis (A) and intercellular CO2 partial pressure in different salt treatments and cvs were also studied.
MATERIALS AND METHODS
Three wheat cvs differing in their salt sensitivity [CR (Iranian salt tolerant), Ghods (Iranian salt-sensitive) and Kharchia-65 (standard salt tolerant)] were grown in controlled environment conditions at the Department of Agricultural and Environmental Sciences, University of New Castle Upon Tyne, UK, Department of Agronomy, Ferdowsi University of Mashad, Iran. Four levels of salinity [0 (control), 100, 200 and 300 mol m -3 of NaCl and CaCl2] were imposed. All above ground plant parts were taken as shoot sample 4, 12, 24, 72 h after salinisation and 14, 21 and 28 days after the start of salt treatments (Two leaves growth stage to flag leaf emergence) and also in the time of ripening. Three plants in each cv. in each level of salinity and each replication were harvested and Δ was analysed in shoots and grain of different wheat cvs.
Shoots were oven-dried to a constant weight at 70°C, just after sampling. The dried leaves were ground in liquid nitrogen by mortar and pestle to pass a 2 mm sieve. Ground samples of 1.0 to 1.5 mg were placed in a tin foil cap, taking care not to touch the foil during preparation. Carbon and nitrogen isotope composition (δ15C and δ15N, respectively) of the leaf samples were then analysed in a Europa Automated Nitrogen Carbon Analysis-Solids/Liquids ANCA-SL System (Europa Ltd., Crewe, UK). Carbon isotope discrimination (Δ) was calculated as:
(2) |
Where, δa and δp are the carbon isotope composition of source air and plant material, respectively, relative to the international standard Pee Dee Belemnite (PDB) (Hubick et al., 1986). The values of δa and δp were measured as described by Hubick et al. (1986). δ13C values were obtained in parts per thousand (‰).
Total nitrogen concentration of shoots and grain was measured and calculated as μg mg -1 dry weight. Carbon nitrogen ratio (C:N) was also calculated by dividing the total amount of carbon to nitrogen in each sample.
The data were subjected to balanced analysis of variance by Minitab Statistic Software for Windows version 9.2 (Minitab Inc., 3081 Enterprise Drive, PA 16801-3008, USA). In order to show the result of all shoot samples in different times (from 4 to 672 h after salinisation) log of time in hour was used. All regression lines were fitted by using the Scientific Figure Processor software (Fig. P. Software Corporation, Durham, NC, USA).
RESULTS
Carbon isotope discrimination (Δ) decreased with increasing salinity, e.g., the average Δ of three cvs were 22.6x10-3 and 19.9x10-3 at control and 300 mol m-3 salinity, respectively (Fig 1a). The salt-sensitive cv, Ghods, showed significantly lower Δ than the other two cvs at all levels of salinity and the salt tolerant cv, Kharchia-65 had a higher Δ at 200 mol m-3 salinity (Fig. 1a). Δ declined also with age of the plants particularly in samples of the first three days after salinity imposed (Fig. 1b). Different genotypes showed significant differences in Δ between different times after salinisation and this difference was quite significant in the first 24 h after salinisation (Fig. 1).
Figure 2 shows the average Δ of cvs at high salinity and control conditions in different times after salinisation. Because there was no significant differences in Δ amongst wheat cvs at control and 300 mol m-3 salinity, data of three cvs were pooled in the Fig. 2a and b. As is shown in the Fig. 2, the trend of Δ at control and high level of salinity was quite different, i.e., Δ increased with time at control conditions while it decreased with time after salinisation at highest level of salinity.
Carbon isotope discrimination in shoots was compared with pi/pa, as obtained from gas exchange measurement during the lifespan of the plants Figure 3 shows relationships between Δ in shoots, photosynthesis and pi/pa.
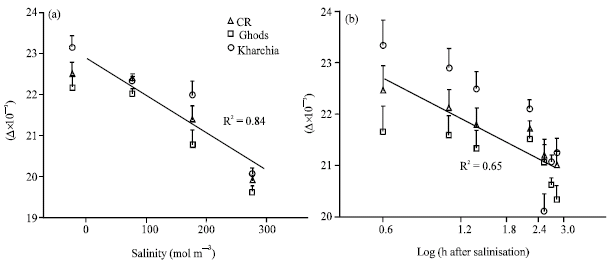 | |
| Fig. 1: | Salinity (a) and time after salinisation (b) on shoot carbon isotope discrimination (Δ). Each point is the average of four levels of salinity and two times measurements. Vertical lines are standard error of means for each point at 95% probability |
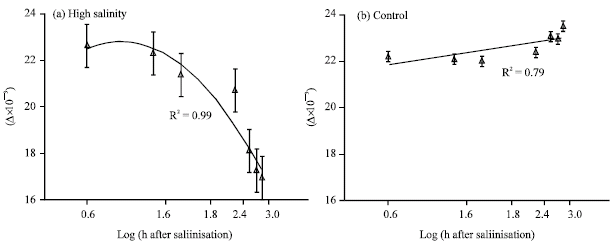 | |
| Fig. 2: | Shoot carbon isotope discrimination (Δ) at high salt concentration (a) and control conditions (b) in different age from 4 h to four weeks after salinisation. In Fig. 1b each point is the average of two times of measurement. Vertical lines are standard error of means for each point at 95% probability |
A positive correlation was observed between pi/pa and Δ in shoots. Δ values of samples were much higher than the theoretical values calculated from Equation 1, but the slope of the sample line was less than that the theoretical line (Fig 3a). The data of highest level of salinity showed a marked decline in Δ, while pi/pa at this level was not lower than those of 100 and 200 mol m-3.
The relationship between Δ and assimilation rate (A) was positive as it was for pi/pa and Δ (Fig 3a). A marked increasing between photosynthetic data of control and salinity was observed (Fig. 3). Control condition plants also showed a higher pi/pa which indicate more stomatal conductance in these plants (Fig. 3b).
Wheat grains in all cvs were always isotopically heavier with respect to shoots. Thus, the discrimination was less in the grain than the shoots. This difference between Δ of shoots and of grain increased with increasing salinity (Table 1). However, the sensitive wheat cv had no significant difference in Δ of grain with other cvs, in fact its Δ was even slightly more than that of the other two cvs at all levels of salinity, which was the opposite of Δ in the shoots (Table 1). Carbon isotope discrimination of grains was at least 5‰ lower than shoots in control and all levels of salinity and as salinity increased the difference between Δ in the shoots and grains also increased.
The best correlation, a parabolic regression line, was plotted between Δ in shoot and grain (Fig. 4). The results showed that at the highest level of salinity one is not a good predictor for another, but in other levels of salinity the relationship between Δ in shoot and grain is positively linear (R2 = 0.66) (Fig. 4a).
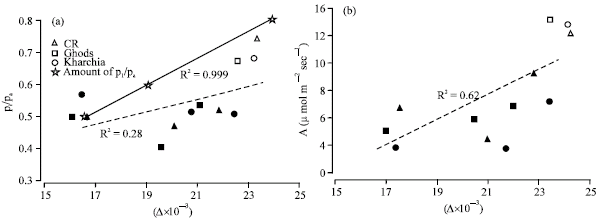 | |
| Fig. 3: | Relationship between shoot carbon isotope discrimination (Δ) pi/pa (a) and photosynthesis (A) (b) of wheat cvs in the presence of salinity. Each point is the average of seven replications. Open symbols are control and solids are salinity. Solid line in (a) is drawn from equation 1 with a = 4.4% and b = 29% |
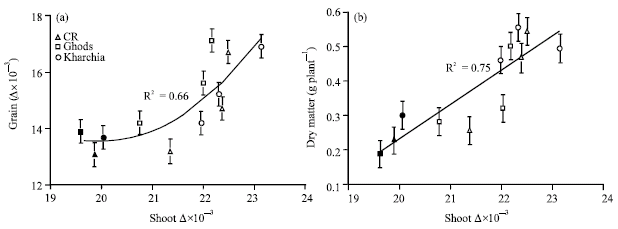 | |
| Fig. 4: | Best fit relationship between shoot and grain carbon isotope discrimination (Δ) (a) and shoot dry weight (b) of wheat cvs in the presence of salinity. Each point is the average of seven replications. Open symbols are control, 100 and 200 mol m-3 and solids are high level of salinity. Vertical lines are standard error of means for each point at 95% probability |
| Table 1: | Comparison of carbon isotope discrimination (Δx10-3) in shoot and grain of wheat cultivars in different levels of salinity. Each number is the average of seven measurements in shoots and two replications in grains |
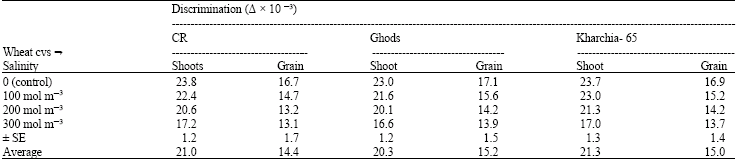 | |
There was a positive correlation between dry matter accumulation and Δ in the presence of salinity (Fig. 4b).
Salinity significantly reduced the shoot N concentration at the highest level of salinity, while grain N content of the same levels of salinity increased (Fig. 5). Although N content reduced at all levels of salinity, there were no significant differences between control, 100 and 200 mol m-3 in the shoot N content of average of three cvs. Amongst wheat cvs. CR had the highest N concentration in both control and salinity, but there were no significant differences amongst cvs. at different salinity levels.
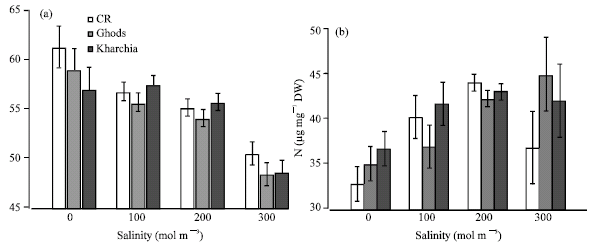 | |
| Fig. 5: | Nitrogen concentration in shoot (a) and grain (b) of wheat cvs in the presence of different salt concentration. Each point is the average of seven times measurements in shoots and two replications in grain. Vertical lines are standard error of means for each point at 95% probability |
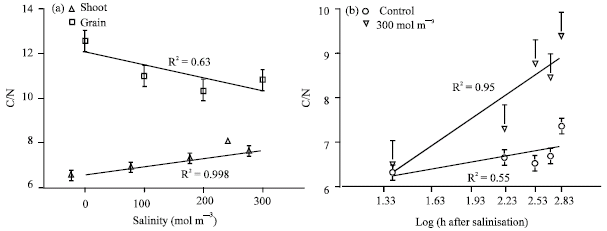 | |
| Fig. 6: | Best fit relationships between salinity (a) and time after salinisation (b) on carbon/nitrogen ratio (C/N) of shoot and grain of wheat cvs. Each point is the average of seven times measurements in part (a) and four levels of salinity in part (b). Vertical lines are standard error of means for each point at 95% probability |
Shoot nitrogen content at 300 mol m-3 was significantly lower than in other salt treatments (Fig. 5a). The overall N content of shoots was between 55.83 μg mg-1 DW in CR and 54.21 in Ghods. Nitrogen content of shoots declined with plant age (57.6 μg mg -1 DW at 4 h after salinisation and 42.7 μg mg -1 DW at four weeks after salt treatment overall salinity levels and cv). In the long-term exposure to salinity, 28 days, N concentrations of shoots were 50.6 μg mg -1 DW in the control conditions and 38.7 μg mg-1 DW at 300 mol m-3. In the grains, N concentration was significantly higher in salt stressed plants than in controls (Fig. 5b). Grain N concentration increased with increasing level of salinity in Ghods and Kharchia-65 but not in CR which had the higher N content in dry weight basis at 100 and 200 mol m-3 than control and 300 mol m-3 salinity (Fig. 5b).
Shoot C:N ratio increased with increasing salinity and time after salinisation (Fig 6). The overall C:N ratio increased from 6.38 at the time of salinisation to 9.1 at 28 days after salinity imposed. In different wheat cvs C:N ratio did not show significant differences either in control or in salt stressed plants. C:N ratio of grain had the opposite trend of C:N in the shoots (Fig. 6).
DISCUSSION
Ghods as a salt-sensitive cv showed consistently but not significantly lower Δ than CR and Kharchia-65 at all levels of salinity indicating that Δ could be used to select for salt tolerance in wheat genotypes. Salt tolerant cvs, CR and Kharchia-65, did not show significant changes at all levels of salinity (Fig. 1a). Although carbon isotope discrimination tended to be reduced by salinity, reduction in Δ was markedly greater at 300 mol m-3 salinity than the other salinity levels (Fig. 1, 2). The relationship of photosynthesis (A) and Δ in Fig. 3 shows that one of the main causes of reduction in A in saline conditions could be stomatal conductance (Kafi, 1996; Robina and Hood-Nowotny, 2005) and this led to reduced discrimination (Farquhar et al., 1989; Brugnoli et al., 1988). The lower values of Δ in stressed plants compared with control, indicate a decrease in CO2 partial pressure of the intercellular space resulting from reduced stomatal conductance (g), confirmed this assuming that there is a positive correlation between g and Δ (R2 = 0.77) (Araus et al., 1997; Fischer et al., 1998; Robina and Hood-Nowotny, 2005).
Figure 3 shows that Δ correlated with the average of pi/pa measured by gas exchange. Based on Equation 1, the expectation is that a positive correlation between Δ in the shoot and pi/pa calculated in gas exchange measurement should exist. In shoots the slope of the line based on Δ measured in samples was less than that of a line drawn based on Eq. 1, i.e., were the theoretical response. The scattering of data might be because the pi/pa measurements done for only a short period of photosynthesis, while the Δ values reflect carbon assimilation over the life of the leaf.
It can be concluded that pi/pa measured by gas exchange in salt stressed plants in this experiment was underestimated compared with theoretical equation except at high level of salinity. For instance, based on Eq. 1, one would expect pi/pa = 0.6 where, Δ= 19.6x 10-3, but pi/pa was 0.5 in this case. Brugnoli and Lauteri (1991) also reported that in bean leaves, although, Δ decreased with increasing salinity, pi/pa measured by gas exchange was unaffected or even increased. They found that in cotton leaves pi/pa had a good correlation with Δ (R2 = 0.87 for leaves). In this experiment, however, pi/pa decreased with increasing salinity up to 200 mol m-3 salinity, when pi/pa was increased at 300 mol m-3 in comparison with control and low levels of salinity.
Relatively more 13C in grain than shoots could be due to existence of more starch in the grains. Brugnoli et al. (1988) postulated that the amount of 13C in the starch was more than that in soluble carbohydrates. They suggested two possible reasons for this difference: Firstly, trios phosphate can either move out of the chloroplast on the phosphate translocator or continue in the Calvin cycle to form starch or RuBP. This could be an equilibrium effect in which 13C preferably concentrates in glyceraldehyde-3-phosphate (GAP) rather than in dihydroxyacetone phosphate (DHAP). DHAP is the preferred substrate for the translocation. Another possibility is that fructose-6-phosphate can take the metabolic pathway to starch or continue in the Calvin cycle to regenerate RuBP. Deleéns-Provent and Schwebel-Dugue (1987) reported that starch was enriched in 13C compared with other carbohydrates in maize seedlings. They argued that translocation between plant parts have different isotopic composition.
Another possible reason for declining Δ in grain compared with shoot could be the time of producing carbohydrate for storage in the kernels. The results showed that with passing time Δ decreased in the presence of salinity. Therefore because these carbohydrates accumulate in the kernels at the last days of the plant’s life, 13C is reduced in the kernels. Farquhar and Richards (1984) suggested that in wheat higher 13C/12C in grain compared to leaf might be partly due to the higher nitrogen content of the grain. In addition, carboxylation of phosphoenolpyruvate (PEP), which discriminates in favour of 13C, is needed for formation of carbon skeletons of some amino acids. Obviously, further information is needed in this area, for understanding this difference between leaf and grain.
Although many researchers found that nitrogen contents were reduced at low levels of salinity e.g., 50 mM NaCl in this experiment N content of shoots did not significantly decline up to 200 mol m-3. The possible reason for these results might be the presence of Ca2+ in the growth medium in the form of CaCl2 (Hawkins and Lewis, 1993).
The other reason could be age of the plants at the time of sampling; a maximum of four weeks after being first salt treated (Munns, 1993). Munns et al. (1995) reported that in wheat plants in the first four to five weeks after salt treatment the NaCl concentration of leaves was below the toxic level.
The nitrogen accumulation pattern in the wheat grains was completely different from shoots. As describes in Fig. 5b it increased with increasing salinity level. This might be because N was expressed in μg g-1 DW and salinity reduced dry weight yield. Thus if it expressed as μg N per kernel, per plant or per spike, the trend of N accumulation would not be as shown in Fig. 5b. In the wheat grain, carbohydrates, mainly starch, accumulate solely in the endosperm, while protein, consisting of nitrogen, is stored in the aleurone layer (up to 30% of total), scutellum and endosperm (Bewley and Black, 1985). Cereal protein content reaches its final level about 20 days after fertilisation, while carbohydrates reach this point only after four to six weeks after fertilisation (Copeland and McDonald, 1995). Therefore the acceleration of senescence in salt stressed plants limits the continuing of carbohydrate accumulation in the grain and it can be another reason for decreasing C:N in grain. Therefore in control and low levels of salinity where kernels are quite filled, the weight ratio of aleurone, scutellum and husk to endosperm is relatively lower than high salt treated wrinkled kernels.
C:N ratio in grain decreased with increasing salinity (Fig. 6). The main reason for this result is the increase in nitrogen content of the grain with salinity while the N concentration of shoots decreased with increasing salinity (Fig. 5). Nitrogen concentration of control, 100 and 200 mol m-3 was not significantly different, thus carbohydrate content of grains must be reduced to cause a decrease in the C:N ratio. In the shoot, C:N ratio increased with increasing salinity only at the most saline treatment. This increment was not only because of carbon accumulation in the shoot, but also of reduction in N content. Reduction in the N content could lead to a higher C:N ratio in this level of salinity. Plant age also caused an increment in C:N ratio in both control and salinity (Fig. 6b). One can say that accumulation of carbohydrates in older organs increased the C content of shoots in control conditions. In the presence of salinity, in addition to accumulation of carbohydrates in older organs, they also may accumulate in the cytoplasm of other tissues as osmotica. The difference between the two lines in Fig. 6b could be partly due to the acceleration of senescence in stressed plants and carbohydrate accumulation as a osmoticum. Low nitrogen absorption in high level of salinity is another reason for the difference between C:N ratio in control and high level of salinity.
Based on the above mentioned results and works reported previously, carbon isotope discrimination might be applicable, as a useful tool, for study of salinity tolerance of wheat genotypes. It shows a remarkable correlation with photosynthesis apparatus and it is responsive to salt stress in cultivars with different salt tolerance even in a short span of time. Therefore, if the mass spectrometer system be available, the measurement of δ is easy and quick. It needs dried small sample size; either leaf, stem, or grain, with minimal problem with storage and handling of samples and analysis of carbon isotopes could be used in salinity studies of wheat genotypes.
REFERENCES
- Araus, J.L., J. Bort, S. Ceccarelli and S. Grando, 1997. Relationship between leaf structure and carbon isotope discrimination in field grown barley. Plant Physiol. Biochem., 35: 533-541.
Direct Link - Brugnoli, E. and M. Lauteri, 1991. Effects of salinity on stomatal conductance, photosynthetic capacity and carbon isotope discrimination of salt-tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C3 non-halophytes. Plant Physiol., 95: 628-635.
CrossRefDirect Link - Farquhar, G.D., Ball, S.M.C., von Ceammerer and Z. Rocsandic, 1982. Effect of salinity and humidity on 13C value of halophytes: Evidence for diffusional isotope fractionation determined by the ratio of intercellular/atmospheric partial pressure of CO2 under different environmental conditions. Oecologia, 52: 121-124.
- Farquhar, G.D., M.H. O'Leary and J.A. Berry, 1982. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol., 9: 121-137.
CrossRefDirect Link - Farquhar, G.D. and R.A. Richards, 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust. J. Plant Physiol., 11: 539-552.
CrossRefDirect Link - Farquhar, G.D., J.R. Ehleringer and K.T. Hubick, 1989. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Mol. Biol., 40: 503-537.
Direct Link - Fischer, R.A., D. Rees, K.D. Sayre, Z.M. Lu, A.G. Condon and A.L. Saavedra, 1998. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate and cooler canopies. Crop Sci., 38: 1467-1475.
Direct Link - Guy, R.D., D.M. Reid and H.R. Krouse, 1986. Factors affecting 13C/12C ratios of inland halophytes. I. Controlled studies on growth and isotopic composition of Puccinellia nuttalliana. Can. J. Bot., 64: 2693-2699.
CrossRefDirect Link - Hawkins, H.J. and O.A.M. Lewis, 1993. Effect of NaCl salinity, nitrogen form, calcium and potassium concentration on nitrogen uptake and kinetics in Triticum aestivum L. cultivar Gamtoos. New Phytol., 124: 171-177.
Direct Link - Hubick, K.T., G.D. Farguhar and R. Shorter, 1986. Correlation between water use efficiency and carbon isotope discrimination in diverse peanut (Arachis) germplasm. Aust. J. Plant Physiol., 13: 803-816.
CrossRefDirect Link - Isla, R., R. Aragues and A. Royo, 1998. Validity of various physiological traits as screening criteria for salt tolerance in barley. Field Crops Res., 58: 97-107.
CrossRef - Merah, O., E. Deleens and P. Monneveux, 1999. Grain yield, carbon isotope discrimination, mineral and silicon content in durum wheat under different precipitation regimes. Physiol. Plant., 107: 387-394.
Direct Link - Munns, R., 1993. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant Cell Environ., 16: 15-24.
CrossRefDirect Link - Munns, R., D.P. Schachtman and A.G. Condon, 1995. The significance of a two-phase growth response to salinity in wheat and barley. Aust. J. Plant Physiol., 22: 561-569.
CrossRefDirect Link - Jiang, Q., D. Roche, A.T. Monaco and S. Durham, 2006. Gas exchange, chlorophyll fluorescence parameters and carbon isotope discrimination of 14 barley genetic lines in response to salinity. Field Crops Res., 96: 269-278.
CrossRefDirect Link - Quarrie, S.A., J. Stojanovic and S. Pekic, 1999. Improving drought resistance in small-grained cereals: A case study, progress and prospects. Pl. Growth Regulat., 29: 1-21.
CrossRef - Shaheen, R. and R.C. Hood-Nowotny, 2005. Effect of drought and salinity on carbon isotope discrimination in wheat cultivars. Plant Sci., 168: 901-909.
CrossRefDirect Link - Voltas, J., I. Romagosa, A. Lafarga, A.P. Armesto, A. Sombrero and J.L. Araus, 1999. Genotype by environment interaction for grain yield and carbon isotope discrimination of barley in Mediterranean Spain. Aust. J. Agric. Res., 50: 1263-1271.
CrossRef