
S. Ekvised
Not Available
S. Jogloy
Not Available
C. Akkasaeng
Not Available
M. Keerati-kasikorn
Not Available
T. Kesmala
Not Available
I. Buddhasimma
Not Available
A. Patanothai
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 6 | Page No.: 923-931







ABSTRACT
The objectives of this study were to estimate broad sense heritability for thrips number parameters and thrips damage parameters and to evaluate correlation between thrips number parameters and thrips damage parameters and correlations of these parameters with agronomic traits. A total number of 192 peanut progenies derived from F2 generation of 16 crosses were evaluated by multiple assessment method staring at 30 days after planting (DAP) for number of thrips adults, number of lavae, number of total thrips, percentage of damaged plants, percentage of damaged leaves and visual damage rating in F4 through F6 generation in three environments under natural infestation of thrip population. Some agronomic characters were evaluated at harvest. A randomized complete block design with six replications in three environments was used. Heritability estimates across evaluation times for thrips number parameters were generally lower and lesser consistent than those for plant damage parameters. Selection for lower thrips number alone would be either inconsistent or ineffective. These results suggest that greater genetic gain for thrips resistance in peanut can be achieved by selection on a family basis using plant damage parameters rather than using thrips number parameters as selection criteria. The lack of correlation between thrips number parameters and plant damage parameters implies that different genetic mechanisms control thrips resistance in this population and inherit independently. Weak correlations between thrips resistance parameters and agronomic characters also indicate the independent inheritance of these characters. Considering high heritability estimates and simplicity of evaluation, the improvement of thrips resistance in this population based on plant damage parameters would be possible.
PDF Abstract XML References Citation
How to cite this article
S. Ekvised, S. Jogloy, C. Akkasaeng, M. Keerati-kasikorn, T. Kesmala, I. Buddhasimma and A. Patanothai, 2006. Heritability and Correlation of Thrips Resistance and Agronomic Traits in Peanut. Asian Journal of Plant Sciences, 5: 923-931.
DOI: 10.3923/ajps.2006.923.931
URL: https://scialert.net/abstract/?doi=ajps.2006.923.931
DOI: 10.3923/ajps.2006.923.931
URL: https://scialert.net/abstract/?doi=ajps.2006.923.931
INTRODUCTION
Thrips (Thysanoptera: Thripidae) are important pests of a number of crops throughout the world. Nine thrips species were found in association with peanut eco-agrosystems in Thailand (Wongkeaw, 1993) and are important as disease vectors rather than as damaging pests (Mound, 1996). Significant yield losses have been noted when heavy infestation of thrips population occurred at early growth stage. Yield losses become greater when associated with drought stress, herbicide application, extreme temperature and viral diseases (Funderburk et al., 1998). Control of thrips is relied heavily on the application of insecticides, in which frequent chemical sprays are needed to achieve the effective control level. The opportunity for use of biological control under field conditions is very less. Additionally, several thrips species are capable to develop insecticide resistance (Daughtrey et al., 1997; Immaraju et al., 1992).
Painter (1951) described three mechanisms of plants resistance to insect pests. Non-preference refers to the groups of plant traits and insect responses that lead to or away from the use of particular plant or variety, for oviposition, for food, for shelter or for combinations of the three. Antibiosis exhibits those adverse effects on the insect life history when the insect uses resistant plant for food. Tolerance denotes the ability of plant to grow and reproduce itself or to repair injury in spite of the supporting thrips population similar to that damaging. Kogan and Ortman (1978) proposed antixenosis to describe more accurately the term of non-preference of insects for a resistant plant.
Two mechanisms underlying resistance to thrips in peanut have been reported. Amin (1985) found that antixenosis was the basis of resistance to thrips in peanut cultivar Robut 33-1 and the resistance was associated with dark green leaf color and spreading growth habit. Dwivedi et al. (1993) reported close association between resistance to thrips and dark green leaf color, leaf wax and hairy characters in peanut cultivar ICGV 86031. Linch and Stalker (1997) reported that antibiosis was a basis of resistance to thrips in wild Arachis species and interspecific hybrids.
The use of cultivars with resistance to thrips is one of the most promising alternative control measures since it is economically and environmentally safe and can be easily integrated with other control measures. Thrips resistance might affect the spread of tospoviruses either in positive or negative way. Effect of thrips resistance in pepper impeded thrips population development (Maris et al., 2003a), restricted spread of Tomato spotted wilt virus (Maris et al., 2003b) and also reduced residence time (Maris et al., 2004). Amin (1985) found that Groundnut bud necrosis virus (GBNV) was reduced in peanut accession Robut 33-1, which was attributed to fewer thrips on the plants on this accession than on susceptible accession. In contrast, thrips resistance might alter thrips feeding behavior leading to the increased spread of tospovirus in thrips resistant cultivars of chrysanthemum (Van de Wetering, 1999).
Most knowledge of thrips resistance in peanut has been derived from other crops and limited number of publications on thrips resistance in peanut is available in literature. Cordona et al. (2002) reported resistance to melon thrips (Thrips palmi Karny) in beans (Phaseolus vulgaris L.) that correlations between visual damage scores and bean production and between damage scores and yield were high and significant. The resistance was enhanced by antixenosis and antibiosis mechanisms (Frei et al., 2003).
Although resistant germplasm lines are available and mechanisms underlying thrips resistance are well understood, effective and reliable screening procedures are also important in developing new resistant cultivars. Furthermore, information on genetic variability and heritability and correlation among traits are of importance for the success of breeding programs. This information provides guidelines for further selection of breeding lines and helps breeders to formulate appropriate breeding strategies. However, the information is not well documented in the literature. Recently, Niyomsil (2004) found low magnitude of additive gene effect for thrips number and thrips damage. Studies on other crops found both high and low magnitude of genetic variability in breeding populations for thrips resistance. Stoner et al. (1989) found that narrow sense heritability estimates for resistance to damage by Thrips tabaci Linderman were high (>90%) in two crosses with common susceptible parent and low (0-11%) in other two crosses with common susceptible parent. In onion, Hamiton et al. (1999) reported, depending on methods, very low broad sense (5.3 and 4.0%) and narrow sense (4.1 and 8.0%) heritability estimates for thrips resistance in the cross TG1015Y x IPA-3.
As part of an undergoing peanut breeding program at Khon Kaen University, thrips resistant lines and/or peanut bud necrosis resistant lines were used as resistant parents to introduce resistance traits into high-yielding commercially available cultivars and early generation progenies were evaluated for combining ability of resistance to peanut bud necrosis virus under irrigated field conditions (Kesmala et al., 2003). The progenies in later generations were evaluated for thrips resistance parameters under field conditions. The objective of this study was to estimate broad sense heritability of thrips resistance parameters and correlations among traits for thrips resistance parameters and agronomic traits.
MATERIALS AND METHODS
Plant materials and experimental procedures: One hundred and ninety-two peanut progenies in the F4 generation of 16 crosses each of which had 12 lines were evaluated under drip-irrigated field condition during January to May 2003 at Khon Kaen University Agronomy Farm in Khon Kaen province in the Northeast of Thailand. These crosses involved four well-adapted high yielding cultivars as female parents (KK 60-3, KKU 72-1, KKU 72-2 and Luhua 11) and four thrips resistant lines as male parents (IC 10, IC 34, ICGV 86031 and ICGV 86388). Descriptions of these parents and cross regeneration were described previously (Kesmala et al., 2003). The resulting F1 crosses were allowed to self-pollinate to obtain F2 seed. F2 generation was used to evaluate combining ability for resistance to peanut bud necrosis virus under field conditions (Kesmala et al., 2003). F3 seeds from single F2 plants were grown for seed increase and then 192 randomly chosen lines of F4 generation derived from F2 plants were available for evaluation.
Under drip-irrigated field conditions in 2003, 192 peanut lines were arranged in a randomized complete block design with 6 replications. A single-row plot, 3.0 m long and spaced 0.5 m apart with 13 plants, was used. Ethrel (2-chloroethylphosphonic acid) was used as seed treatment to break possible seed dormancy. Seeds were over-planted and the seedlings were then thinned to obtain one seedling per hill at 15 days after planting. Neither fungicide nor insecticide was applied to the crop and other cultural practices were in accordance with the recommendations for irrigated peanut, including uniform fertilization, application of gypsum and weed control. Drip-irrigation was supplied to the crop under optimum growing conditions to evaluate agronomic performance of these lines in addition to the evaluation of thrip resistance.
In 2004 and 2005, the experiment was repeated in a peanut growing area in Kalasin province in the Northeast of Thailand, using F5 and F6, respectively, under furrow-irrigated field conditions after rice harvest. The experiment procedures were similar to those for field experiment in 2003.
Data collection: Data were recorded for total thrips number, number of adult thrips and number of larval thrips at 20, 30, 40, 50, 60 and 70 days after planting (DAP). Five plants in each plot were randomly chosen as a sample unit. Four folded leaflets in the same petiole on the main stem of each plant were harvested and immediately put in a small plastic vial containing 70% ethyl alcohol. Thrips count was conducted in laboratory under light microscope (12 x magnifications) and then total thrips number, number of adult thrips and number of larval thrips was determined.
Plant damage by thrips feeding based on 11 plants in each plot were recorded as percent damaged plants (plants having thrips feeding scars) and percent damaged leaves (leaves on main stems showing thrips feeding damage) at 30, 40, 45, 50, 60, 70 and 80 days after planting. Percentage of damaged leaves was recorded based on percentage of leaves showing visible scars on main stems of 11 plants in each plot. In each leaf, a single leaflet or more in a petiole having scars of thrips feeding was considered damaged.
Thrips damage ratings of 1-9 were given to 11 individual plants in each plot, with 1 = no damage to damage 10%, 3 = damage 11-30%, 5 = damage 31-50%, 7 = damage 51-70% and 9 = damage 71-100% (Keerati-Kasikorn and Singha, 1986).
Monitoring of thrips abundance by sticky traps was also conducted in 2005 at 60 and 67 days after planting. The most abundant species was Scirtothrips dorsalis Hood accounting for 48.7% of total thrips adults followed by Megalurothrips usitatus Bagnall (18.7%) and Caliothrips indicus Bagnall (12.8%), respectively (Ekvised et al., 2006). Thus, S. dorsalis was a target species in this study.
Agronomic data were recorded at harvest for pod number per plant, pod dry weight per plant, above ground dry weight per plant, seed weight per plant, 100 seed weight and shelling percentage at two locations in 2003 and 2005 only.
Data analysis: Data of total thrips number, juvenile thrips number and adult thrips number were transformed to square root (sqrt(x + 0.5) for statistical analyses. Traits other than thrips number were also analyzed statistically, using original data.
| Table 1: | Analysis of variance in randomized complete block design in e environments |
 | |
Combined analysis of variance across three locations was also performed for thrips and plant damage parameters except for agronomic data.
Estimates of heritability for 16 crosses were calculated by partitioning variance components of family mean squares to obtain pooled environmental variance (σ2E) and genotypic variance (σ2G) and then broad sense heritability estimates (h2b) were calculated as follows (Table 1):
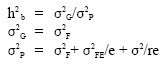 |
Where :
| h2 b | = | broad sense heritability |
| σ2G | = | genotypic variation |
| σ2P | = | phenotypic variation |
| r | = | replications |
| e | = | environments |
Simple linear correlation coefficients were calculated based on data of individual plots of three environments unless mentioned otherwise and all calculations were done by MSTAT-C package (Bricker, 1989). Because agronomic data were recorded at two locations only (data of years 2003 and 2005), correlations related to these characters were calculated based on data in two environments.
RESULTS
Determination of assessment times: The best assessment times for thrips number parameters and plant damage parameters were determined by low coefficients of variation (CV) and high F-ratios from analysis of variance. At 60 and 70 days after planting were the most appropriate assessment times for adult thrips number, juvenile thrips number and total thrips number.
The best assessment times for percentage of damaged plants were relatively narrow and 50 and 60 days after planting could be evaluated more accurately. Assessments at 30 and 40 days after planting were too early and assessments at later than 60 days after planting are too late as plants were severely infested by thrips population.
| Table 2: | Mean squares from combined analysis of variance for thrips number parameters and plant damage parameters of 16 crosses of peanut evaluated across three environments under natural infestation of thrips population |
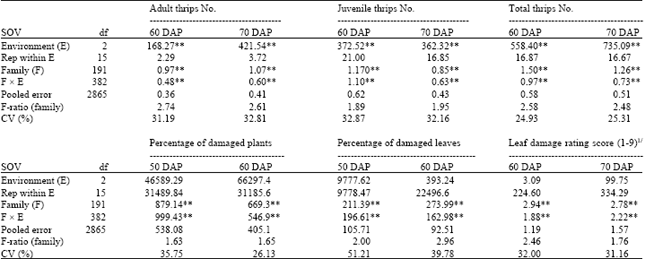 | |
| ** significant at 0.01 probability level, 1/1 = no damage to damage 10%, 3 = damage 11-30%, 5 = damage 31-50%, 7 = damage 51-70% and 9 = damage 71-100%, DAP = Day after planting | |
| Table 3: | Mean squares from combined analysis of variance for agronomic traits of 16 crosses of peanut evaluated across two environments |
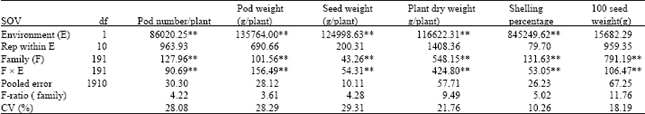 | |
| ** significant at 0.01 probability level | |
The best assessment times for percentage of damaged leaves and leaf damage rating score were relatively wide from 50 to 80 days after planting. Data of 50 and 60 days after planting were selected for percentage of damaged leaves and data of 60 and 70 days after planting for visual damage rating.
Differences among environments were highly significant for adult thips number, juvenile thrips number and total thrips number (Table 2). Differences among families and interactions between family and environment were also highly significant for all traits under study, but family mean squares were much greater than family x environment interaction mean squares except for percentage of damaged plants at 50 days after planting.
Analysis of variance for agronomic traits showed highly significant variations among families for pod number per plant, pod weight per plant, seed weight per plant, plant dry weight, shelling percentage and 100 seed weight (Table 3). Environmental variations were also highly significant for most characters except for 100 seed weight. Family x environment interactions (F x E) were highly significant for all characters and interaction mean squares were quite large, but only mean square for pod weight per plant was larger than its corresponding family mean square. F-ratios of family differences for plant dry weight and 100 seed weight were larger than those for the rest of agronomic traits.
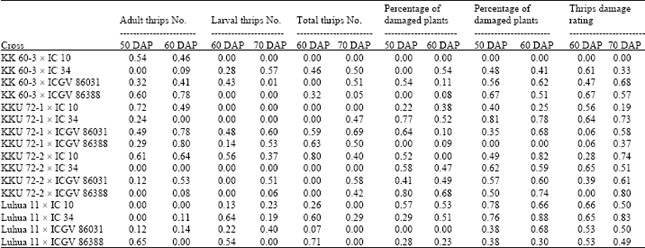
Heritability: Heritability estimates of 16 crosses were calculated for three thrips number parameters and three plant damage parameters (Table 4). The heritability estimates varied among crosses and characters, ranging from zero to 0.88. Estimates of heritability for thrips number parameters were generally lower and lesser consistent than those for plant damage parameters. However, more consistent and moderate estimates among adult thrips number, juvenile thrips number and total thrips number were observed in three crosses (KKU 72-1 x ICGV 86031, KKU 72-2 x IC 10 and KKU 72-1 x ICGV 86388), ranging from 0.14 to 0.80. Heritability estimates of these crosses were less consistent with those for plant damage parameters.
Consistencies of heritability estimates for thrips number parameters between assessment times were also noted, but there were few crosses for each character.
| Table 4: | Estimates of broad sense heritability for thrips number parameters and plant damage parameters of 16 crosses of peanut evaluated across three environments under natural infestation of thrips population |
 | |
| Table 5: | Simple linear correlation coefficients between thrips number parameters and thrips damage parameters of 16 crosses of peanut evaluated across 3 locations under natural infestation of thrips population (with 3455 degree of freedom) |
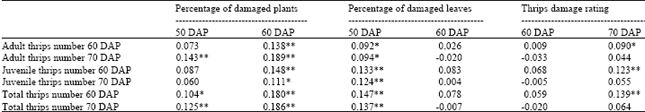 | |
| *, ** Significant at 0.05 and 0.01 probability levels, respectively with 3455 degree of freedom, DAP = Day after planting | |
| Table 6: | Simple linear correlation coefficients between thrips number parameters of 16 crosses of peanut evaluated at 3 locations under natural infestation of thrips population (with 3455 degree of freedom) |
 | |
| ** Significant at 0.01 probability level with 3455 degree of freedom, DAP = Day after planting | |
For adult thrips number, the crosses with consistent and meaningful heritability estimates were KK 60-3 x IC 10, KK 60-3 x ICGV 86031, KK 60-3 x ICGV 86388, KKU 72-1 x IC 10, KKU 72-1 x ICGV 86031 and KKU 72-1 x IC 10. The promising crosses for making selection for lower juvenile thrips number were KK 60-3 x IC 34, KKU 72-1 x ICGV 86031 and KKU 72-2 x IC 10 and the crosses KK 60-3 x IC 34, KKU 72-1 x ICGV 86031 and KKU 72-1 x ICGV 86388 showed good possibility for selecting lines with lower total thrips number.
Estimates of heritability for plant damage parameters were generally higher and more consistent than those for thrips number parameters. The most consistent crosses across the characters were KKU 72-1 x IC 34, KKU 72-2 x IC 34, KKU 72-2 x ICGV 86031, Luhua 11 x IC 10 and Luhua 11 x IC 34. Percentage of damaged plant had lesser and lower heritability estimates than did percentage of damaged leaves and thrips damage rating score and the cross KKU 72-1 x ICGV 86388 showed high and consistent heritability estimates between assessment times. Several crosses (KK 60-3 x IC 34, KK 60-3 x ICGV 86031, KK 60-3 x ICGV 86388, KKU 72-1 x IC 10, KKU 72-2 x IC 10, Luhua 11 x ICGV 86031 and Luhua 11 x ICGV 86388) showed high and consistent heritability estimates between percentage of damaged plants and thrips damage rating score and also between assessment times. However, the two crosses (KKU 72-1 x ICGV 86031 and KKU 72-2 x IC 10) had high and consistent heritability estimates for percentage of damaged leaves only.
Simple linear correlation: Simple linear correlations between thrips number parameters and thrips damage parameters were presented in Table 5 and correlations among groups of thrips number parameters and plant damaged parameters were reported in Table 6 and 7, respectively.
| Table 7: | Simple linear correlation coefficients between thrips damage parameters of 16 crosses of peanut evaluated across 3 locations under natural infestation of thrips population (with 3455 degree of freedom) |
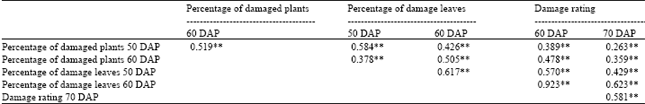 | |
| ** Significant at 0.01 probability level with 3455 degree of freedom, DAP = Day after planting | |
| Table 8: | Simple linear correlation coefficients between thrips resistance parameters and agronomic traits of 16 crosses of peanut evaluated in one environment in Kalasin in 2005 at 50, 60 and 70 days after planting (DAP) under natural infestation of thrips population |
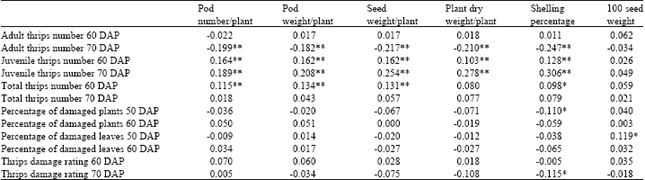 | |
| *,** Significantly different from zero at 0.05 and 0.01 probability levels, respectively with 2303 degree of freedom | |
In general, correlations between thrips number parameters and plant damage parameters were from non-significant to highly significant and positive (p≤0.01), but the associations were relatively low when compared with the associations among characters within the same group of characters. Better associations between thrips number parameters and percentage of damaged plants at 60 days after planting and percentage damaged leaves at 50 days after planting were noted as all correlations coefficients were significant. Few significant correlation coefficients were indicated between thrips number parameters and percentage of damaged plants at 50 days after planting and between thrips number parameters and visual damage rating at 60 days after planting. However, all correlation coefficients between thrips number parameters and percentage of damaged leaves at 60 days after planting and between thrips number parameters and visual damage rating at 60 days after planting were not significant (Table 5).
Highly significant (p≤0.01) and positive correlation coefficients among thrips number parameters were observed for all combinations (Table 6). The correlation coefficients were from moderate (r = 0.236, p≤0.01) to high (r = 0.894, p≤0.01). Similar results were also found for correlation coefficients among plant damage parameters. All correlation coefficients were highly significant (p≤0.01) and positive, ranging from 0.263 to 0.923 (Table 7).
Agronomic traits were not generally associated with plant damage parameters. However, shelling percentage was slightly and negatively associated with percentage of damaged plants at 50 DAP (r = -0.110, p≤0.01) and visual damage rating at 70 DAP (r = -0.115, p≤0.05) (Table 8). One hundred seed weight was positively associated with percentage of damaged leaves at 50 DAP (0.119, p≤0.01).
Low associations for both directions were observed between thrips number parameters and agronomic traits. Adult thrips numbers at 70 DAP were negatively associated with pod number/plant (r = -0.199, p≤0.01), pod weight/plant (r = -0.182, p≤0.01), seed weight/plant (r = -0.217, p≤0.01), plant dry weight/plant (r = -0.210, p≤0.01) and shelling percentage (r = -0.-0.247), but these associations did not exist at 50 DAP.
Correlation coefficients between juvenile thrips number at 60 and 70 DAP and pod number/plant, pod weight/plant, seed weight/plant, plant dry weight/plant and shelling percentage were all positive and highly significant (p≤0.01), with the correlation coefficients ranging from 0.103 to 0.306. Total thrips number at 60 DAP was positively and significantly associated with pod number/plant (r = 0.115, p≤0.01), pod weight/plant (r = 0.134, p≤0.01), seed weight/plant (r = 0.131, p≤0.01) and shelling percentage (r = 0.098, p≤0.05), except for plant dry weight/plant and 100 seed weight, but none of agronomic traits was associated with total thrips number at 70 DAP.
DISCUSSION
Determination of assessment times: Evaluation of plant reactions to insect pests is usually based on multiple time evaluation. It is important to determine which assessment times are most appropriate and give the most accurate data. This information is useful for peanut breeders to eliminate unnecessary assessment times. Assessments at early growth stages may not be appropriate because fewer thrips numbers were detected and immigrating thrips population needed more time to establish in the test field. Our previous work indicated that thrips number evaluation could be done at early as 40 days after planting but not at 30 days after planting (Ekvised et al., 2006). The most appropriate assessment times could not be exactly determined because many factors influencing thrip population and there would be spatial and temporal differences. However, they should be varies between 40 to 80 days after planning.
As plant damage is resulted by thrip feeding, it should occur some days after thrip population enters to the test field. However, the appropriate times to evaluate percentage of damaged plants are very narrow from 40 to 60 days after planting. When valuations were done at early growth stages, very few plants in the plots showed visual damage, whereas most plants in the plots showed visual damage at later growth stages.
Numbers of damaged leaves on main stems were then calculated as percentage and could be assessed with reasonable accuracy as early as 40 days after planting. The possible assessment times were wider than other characters evaluated and could be assessed until 80 days after planting with similar results. However, the best assessment times would be at 50 and 60 days after planting. Visual damage score followed the similar pattern of percentage of damaged plants, but the best assessment times (60-70 DAP) were somewhat later than those for percentage of damaged leaves.
Significances of family mean squares for all characters indicated that genetic variability existed and heritability estimates were calculated based on plot means of individual families. Data for thrips number parameters and thrips damaged parameters were combined from three environments to eliminate the variances of family x environment interactions from total genetic variances. Genetic variances were much purer than those for individual environment. The tested progenies were subjected to several generations of inbreeding and non-additive variances were not important because they should be diminished after successive inbreeding. Therefore heritability estimates should be good indicators for genetic variation in this population.
Heritability: Scirtothrips dorsalis was the most abundant species and was persistently the first rank for several years. The data were well supported previous studies conducted in different years and locations in the dry season. Buddhasimma (2004) reported the most abundance of S. dorsalis followed by C. indicus in peanut with increasing number after seven weeks of planting. Chuapong (1997) also found predominance of S. dorsalis in the trials for peanut bud necrosis disease, which is vectored by Thrips palmi Karny. The breeding objective is to against S. dorsalis. However, other species which were present in a considerably large portion could confound the results.
Heritability estimates by method of calculation were broad-sense. However, the materials were in more advanced generations or near linbred lines and F x E variances were eliminated by combined analysis of variance. Therefore, additive genetic variance accounted for most of the gene action possible for these characters under study. Environmental and F x E variances were the most of phenotypic variance.
Lack of consistency and low heritability estimates for parameters of number thrips adults, number of lavae and number of total thrips were not unexpected due to dynamic nature for these characters. The temporal and spatial occurrence of thrips varied with sampling methods, crop species, regions, localized areas within region (Eckel et al., 1996) and even different plant parts within plant (Reitz, 2002). Hamilton et al. (1999) reported low heritability for numbers of onion thrips in onion because thrips populations are clumped rather than randomly distributed. Similarly, Culbreath et al. (2000) did not found consistent cultivar effect of peanut on adult tobacco thrips (F. fusca) or adult western flower thrips (F. occidentalis). Differences among cultivars for numbers of lavae of Flankliniela sp. also were few and inconsistent. Jan de Kogel et al. (1997) found that differences among chrysanthemum cultivars for thrips resistance were observed in winter but disappeared in summer. Furthermore, high variations of F x E interactions might cause low heritability estimates and extensive evaluations in several environments are required to identify superior genotypes. Therefore, because of difficulty to assess thrips numbers accurately under field conditions and low and inconsistent heritability estimates, the reliability in selecting a single plant that is truly genetically resistant or susceptible is very low. However, a family based selection scheme should be used to breed for thrips resistance in this peanut population due to low levels of heritability.
Reactions of parental lines to thrips infestation were also studied and reported in the accompanying paper (Ekvised et al., 2006). However, differences between parental lines for thrips numbers did not guaranteed the higher heritability estimates in their progenies. For example, Progenies involving IC 10 as male parent generally showed low heritability estimates. IC 10 showed consistently low thrips number. In the course of evaluations, parental lines responded differently to thrips infestation depending mainly on differences in severity of insect pressure at different times of evaluations. Also differences between parental lines were not large and the thrips resistant lines from ICRISAT showed reactions to thrips infestation similar to those of commercially available cultivars (KK 60-3, KKU 72-1 and KKU 72-2). Possible explanations for the similar reactions of the cultivars include the difference in components of thrips species in India and Thailand and the commercial cultivars which were developed in Thailand should have some degree of thrips resistance after extensive testing for agronomic performance under growing conditions in Thailand.
Heritability estimates for parameters of percentage of damaged plants, percentage of damaged leaves and thrips damage rating were generally higher and more consistent than those for thrips number parameters possibly due to static nature of the characters. These characters would be more useful than thrips number parameters for evaluating thrips resistance considering the simplicity to evaluate and their discriminating ability to separate the differences among peanut genotypes. Kumar et al. (1996) reported that rating for thrips damage was more reliable and efficient than estimating thrips number in screening pepper accessions against S. dorsalis.
The heritability estimates for these characters are high enough to be used as an indicator for the success of further selection scheme. The consistency of heritability estimates across evaluation times might indicate that these characters were less affected by environmental variations.
Simple linear correlation: Simple linear correlation among characters is useful information for breeders to formulate effective simultaneous selection strategies. High correlation coefficients indicate the close association between characters. The correlation coefficients, though significant, will not give a strong confidence if the values are not larger than 0.5.
Correlations between thrips number parameters and plant damage parameters were not well associated. This was possibly due to the difference in nature of the characters. Plant damage parameters were static, whereas thrips number parameters were dynamic and these characters were evaluated at specific times. Peanut genotypes might respond differently to thrips infestation at different evaluation times depending on insect pressure.
Correlations within the same groups of characters were quite well associated. For example, adult thrips number was well associated with juvenile thrips number and both characters made contribution to total thrips number. In a similar way, most correlation coefficients between percentage of damaged plants and percentage of damaged leaves were relatively high and positive and both characters were also well associated with visual damage rating.
Peanut cultivars with resistance to biotic and physical stresses are the ideal goal of any peanut breeding program. Thrips resistant lines should be of acceptable agronomic performance and thus correlation between thrips resistance parameters and agronomic traits is a useful guideline for simultaneous selection program. Generally, correlation coefficients between agronomic characters and thrips resistance parameters were not significant. Although some combinations were significant for both positive and negative directions, none of the correlation coefficients exceeded 0.5, which was considered to be useful. It has been concluded that thrips resistance parameters and agronomic characters were not associated. The results suggested that different genetic systems control both groups of characters, causing random segregation of the characters among progenies. It is implied that selection for one character will not affect another character. Two-stage selection for thrips resistance and then for agronomic characters should be carried out in this population.
ACKNOWLEDGEMENTS
This study was funded by the Senior Research Scholar Project of Prof. Dr. Aran Patanothai under the Thailand Research Fund and also supported by the Peanut Improvement Project, Faculty of Agriculture, Khon Kaen University, Khon Kaen, Thailand.
REFERENCES
- Amin, P.W., 1985. Apparent resistance of groundnut cultivar Robut 33-1 to bud necrosis disease. Plant Dis., 69: 718-719.
Direct Link - Daughtrey, M.L., R.K. Jones, J.W. Moyer, M.E. Daub and J.R. Baker, 1997. Tospovirus strike the greenhouse industry: INSV has becomes a major pathogen on flower crops. Plant Dis., 81: 1220-1230.
Direct Link - Eckel, C.S., K. Cho, J.F. Waigenbach, G.G. Kennedy and J.W. Moyer, 1996. Variation in Thrips species composition in field crops and implications for tomato spotted wilt epidemiology in North Carolina. Entomol. Exp. Applicata, 78: 19-29.
Direct Link - Funderburk, J.E., D.W. Gorbet, I.D. Teare and J. Stavisky, 1998. Thrips injury can reduce peanut yield and quality under conditions of multiple stress. Agron. J., 90: 563-566.
Direct Link - Hamilton, B.K., L.M. Pike, A.N. Sparks, D.A. Bender, R.W. Jones, J. Candeia and G. De Franca, 1999. Heritability of thrips resistance in the IPA-3 onion cultivar in South Texas. Euphytica, 109: 117-122.
CrossRef - Immaraju, J.A., T.D. Paine, J.A. Bethke, K.L. Robb and J.P. Newman, 1992. Western flower thrips (Thysanoptera: Thripidae) resistance to insecticides in costal California greenhouses. J. Econ. Entomol., 83: 9-14.
Direct Link - Jan de Kogel, W., M. Van der Hoek, M.T.A. Dik, B. Gebala, F.R. van Dijken and C. Mollema, 1997. Seasonal variation in resistance of Chrysanthemum cultivars to Frankliniela occidentalis (Thysanoptera: Thripidae). Euphytica, 97: 283-288.
CrossRef - Kumar, N.K.K., M. Aradya, A.A. Deshpande, N. Anund and P.R. Ramachandar, 1996. Initial screening of chili and sweet pepper germplasm for resistance to chili thrips, Scirtothrips dorsalis Hood. Euphytica, 89: 319-324.
Direct Link - Maris, P.C., N.N. Joosten, R.W. Goldbach and D. Peters, 2003. Restricted spread of tomato spotted wilt virus in thrips resistant pepper. Phytopathology, 93: 1223-1227.
Direct Link - Maris, P.C., N.N. Joosten, R.W. Goldbach and D. Peters, 2004. Decreased preference and reproduction and increased mortality of Frankliniela occidentalis on thrips resistant pepper plants. Entomol. Exp. Appl., 113: 149-155.
Direct Link - Mound, L.A., 1996. The thysanoptera vector species of tospoviruses. Acta Hortic., 431: 298-309.
Direct Link - Reitz, S.R., 2002. Seasonal and within plant distribution of Frankliniella thrips (Thysanoptera: Thripidae) in North Florida tomatoes. Florida Entomol., 85: 431-439.
Direct Link - Stoner, K.A., M.H. Dickson and A.M. Shelton, 1989. Inheritance of resistance to damage by Thrips tabaci Lindeman (Thysanoptera: Thripidae) in Cabbage. Euphytica, 40: 233-239.
Direct Link - Maris, P.C., N.N. Joosten, R.W. Goldbach and D. Peters, 2003. Spread of Tomato spotted wilt virus and population development of Frankliniela occidentalis in pepper resistant to thrips. Proceedings of the Section Experiment and Applied Entomology Netherlands Entomological Society, 2003, Amsterdam, pp: 95-101.