
A. Gholamhoseinian
Not Available
N. Ghaemi
Not Available
F. Rahimi
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 5 | Page No.: 827-831







ABSTRACT
Catalase (EC. 1.11.1.6.H2O2. oxidoreductase) has been found in all aerobic organisms. Most of the work preformed on this enzyme obtained from mammalian, bacterial and fungal sources as there is less information about plant catalases. Partial purification of catalase from Brassica oleracea capitata (Cabbage) and its kinetics was studied. To this intention, freshly harvested cabbage leaves freezed in liquid nitrogen, reduced to small pieces and blended. The extraction with 0.1 M Na2 HPO4 buffer solutions have performed. The filtrate after centrifugation half-saturated with solid Am-Sulfate (A.S) then 35% saturated with solid A.S. After the partially purified enzyme dialyzed, the extract was eluted from a sephadex G-200 column equilibrated with phosphate buffer. The enzymatic activity was observed in only one peak. The optimal pH of the cabbage leaf catalase was 7-8. When the concentration of stabilized catalase increased, the reaction rate increased concomitantly. The substrate was not inhibitory to the reaction rate up to 0.1 M of H2O2 concentration. In this study Vmax and Km of cabbage leaf catalase was 31.12 μM min-1 and 25.5 mM, respectively.
PDF Abstract XML References Citation
How to cite this article
A. Gholamhoseinian, N. Ghaemi and F. Rahimi, 2006. Partial Purification and Properties of Catalase from Brassia oleracea capitata. Asian Journal of Plant Sciences, 5: 827-831.
DOI: 10.3923/ajps.2006.827.831
URL: https://scialert.net/abstract/?doi=ajps.2006.827.831
DOI: 10.3923/ajps.2006.827.831
URL: https://scialert.net/abstract/?doi=ajps.2006.827.831
INTRODUCTION
Catalase is a major primary antioxidant defense component that primarily catalyses the decomposition of H2O2 to H2O. The enzyme is one of the earliest enzyme had been studied and purified (Percy, 1984). Since the preparation of crystalline catalase from beef liver (Sumner and Dounce, 1937) crystalline catalase has been obtained from a number of other sources. These are lamb liver (Dounce, 1942), Bovine brain ( Choi et al., 1999) Human liver (Jin et al., 2003) and Vibrio rumoiensis (Yumoto et al., 2000). Catalase has been found in all aerobic cells containing cytochorom (Percy, 1984). Different organisms have shown different catalases, for example, S. cerevisia have two type of enzyme called as A and T (Izawa, 1996).
Although the enzyme is present in green plants, but usually it had been extracted from those with higher concentration in their cells. The purification from plant cells such as potato tuber (Beaumount, 1990), wheat germ (Garcia et al., 2000) and spinacia (Galeston, 1955) have been reported. The latest was the earliest enzyme had been purified.
In 1996, a special catalase from tylakoid membrane of Spinach leaves associated with photo system was purified which was different in their molecular weight, pH optimum, sensitivity to variety of inhibitors and their UV-visible spectra (Sheptovitsky and Brudvig, 1996). Few catalases have been purified from plants ; they include spinach (Galeston, 1955 ) , Van apple (Yoruk et al., 2005) and wheat germ (Garcia et al., 2000). Since various catalases purified so far had shown different characteristics and due to abundance of cabbage in the food stuff, it used as another source for the purification of this enzyme. No record on the purification and kinetics of catalase from the cabbage had been found in the literature. Most of the studies had been focused on the effect of different types of stress on the activity of this enzyme , these include zinc toxicity (Prasad et al., 1999); allevation of water-logging (Leul and Zhou, 1999), harmful effect of Hg (Ma-Chang and Ma, 1998), effect of salt (Verma and Mishra, 2005) and cadmium toxicity (Singh and Tewari, 2003).
Here we partially purified the catalase from the cabbage leaves by means of conventional methods such as ammonium sulfate precipitation and gel filtration and the kinetics were determined.
MATERIALS AND METHODS
The fresh leaves of cabbage (B. oleracea) were harvested during spring from Tehran area, washed with tap water, chapped and immerged in liquid nitrogen for a few minutes.
| Table 1: | Purification of Catalase from Brassia olercea capitata |
 | |
The ice dried leaves then smashed and layered over a cheese cloth, the resulting powder was washed with cold (0-2°C) 0-1M phosphate buffer, pH = 7, with equal amount of the original wet weight. The eluent then centrifuged at 4°C, at 8000 RPM. in an IEC-B-20 refrigerator equipped centrifuge for 10 min.
Enzyme assays: Catalase activity was routinely assayed spectero-photometrically at 25°C following the decrease in absorption at 240nm in 10 mM H2O2 in 50 mM sodium phosphate buffer pH = 7 (Aebi, 1984), protein content was determined by modified procedures (Lowry et al., 1951; Spector, 1978). Bovine Serum Albumin (BSA) was used as the standard.
The enzyme unit was defined as the amount of enzyme that catalyzes the oxidation of one μmole hydrogen peroxide per min under the assay conditions. The specific activities were expressed in term of enzyme units per mg protein.
Enzyme purification
First ammonium sulfate precipitation: Crude extracts was adjusted to 50% saturation with powder of ammonium sulfate, left overnight at 4°C followed by centrifugation at 10000 RPM for 10 min. This precipitate was resuspended in small volume of 0.1M phosphate buffer pH = 7.
Second ammonium sulfate precipitation: The enzyme suspension from step one was brought to 35% saturation by ammonium sulfate, left overnight at 4°C followed by centrifugation at 10000 RPM for 10 min. This precipitate was resuspend in minimum volume of 0.1 m phosphate buffer pH = 7.
Gel filtration: After dialyzing the (NH4)2 SO4 solution through dialysis bag (two days stirring in 0.1M buffer), the enzyme solution was clarified by centrifugation. The supernatant was applied to a Sephadex G200 column (2x20 cm) equilibrated with 0.1 M phosphate buffer pH 7. The catalase was eluted with the same buffer.
RESULTS
Enzyme purification: The results obtained for the partial purification of catalase from Brassica oleracea capitata (BOC) extract summarized in Table 1.
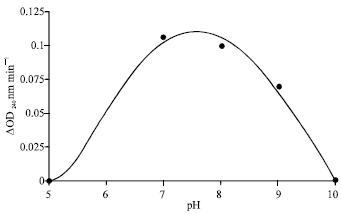 | |
| Fig. 1: | Catalase activity as a function of pH for cabbage extract. Assays were done according to the standard condition, except that the pH of incubation mixtures was varied. The buffers used were citrate phosphate, Tris-HCl, glycine-NaOH, the enzyme source were protein extracted from 35% (NH4)2 SO2 fractionation |
First (NH4) SO4 fractionation gave rise to 13 fold purification. After subsequent salt precipitation (35% saturation), the purification fold increased to about 66 folds. Gel filtration of the second step by Sephadex G200 resulted in partially purified catalase with almost 82 fold purification but with a low recovery of 0.5%.
Kinetic studies on the partially purified enzyme
Dependence of enzyme activity on pH: The effect of pH on the activity of the enzyme from BOC is shown in Fig. 1. As indicated, the pH optimum of the enzymes was found to be between 7 and 8.
Effect of the enzyme concentration: The activity of the enzyme was not linear at low protein concentration (Fig. 2), but at high protein concentration (supplemented with 12 mg mL-1 solution of albumin as diluents) it became linear (Fig. 3).
Dependence of enzyme activity on substrate concentration: Using different concentration of H2O2, the activity of the enzyme was assayed under the standard condition at pH = 7. The initial velocities for the enzyme in terms of enzyme units, as a function of H2O2 concentration was increasing.
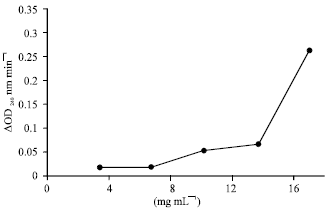 | |
| Fig. 2: | The effect to protein concentration on the activity of the Catalase semi-purified from cabbage. Assays were done under standard condition except that the protein concentration was varied as indicated. The dialyzed sample extracted from 35% (NH4)2 SO4 was used as the enzyme source. d H2O was used as diluents |
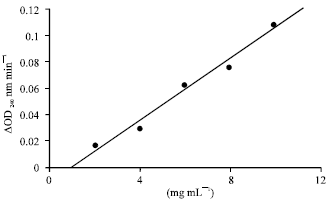 | |
| Fig. 3: | Effect of protein concentration on the activity of the partial purified Catalase from Cabbage diluted with protein solution. Assays were done under the standard conditions except the enzyme solution diluted with BSA to final concentration of undiluted sample |
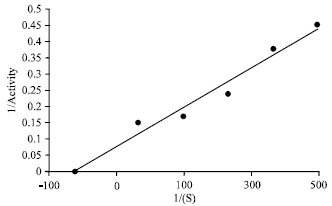 | |
| Fig. 4: | Lineweaver-Burk double reciprocal plot at different H2O2 concentration for cabbage catalase. Assays were done according to the conditions described under optimum condition. The enzyme source was protein extract from 35% (NH4)2 SO4 fractionation |
In order to calculate the Km and Vmax values. The lineweaver Burk double reciprocal plot for the effect of H2O2 concentration on enzyme activity was performed. From the intercept of the regression lines, Km value of 25.5 mM with Vmax of 31.12 μ mol min-1 for BOC catalase was found (Fig. 4).
DISCUSSION
Catalase is widespread in nature, having been found in all aerobic organisms studied to date. Most of the work has been performed on the enzyme obtained from mammalian and bacterial sources where it is present in highest concentration (Jin et al., 2003; Yumoto et al ., 2000 ).
The enzymes from different species have been found to be quite similar in physical and structural properties except for an enzyme from Baker's yeast (Jacob and Johnson, 1979). Among the plants, Spinacia oleracea is one of earliest enzyme which had been studied (Galeston, 1955). The enzyme was characterized in terms of molecular weight and subunit size, amino acid composition, UV-visible absorption spectra, heme content, pH optimum, inhibitor sensitivity and Km value for H2O2.
No study has been performed for purification and properties of any different species of Brassica so far to correlate our findings to them. All researches were confined on the effect of stress on reduction of catalase activity in this plant (Verma and Mishra, 2005; Singh and Tewari, 2003).
When acetone dehydrated leaves (Galeston method) was used for Brassica oleracea no enzyme activity was detected. These phenomena may be due to lack of cytoplasmic catalase and organelle’s removal of catalase under such a condition. For this reason, the leaves of Brassica oleracea were freezed by liquid nitrogen. The leaves were now very brittle and could be blended. The powder then extracted repeatedly with ice cold 0.1M Na2 HPO4, buffer, yielding the filtrate, which then used for ammonium sulfate precipitation. Two step fractionation by 50 and 35% saturation gave rise to 66 folds purification. For determination of iso-enzyme profile non-denatured page was performed on protein precipitated of the final step. No catalase activity was revealed under this condition (results not shown). After gel filtration only one active fraction was obtained with 82.3 folds purification .this was 10 times as much as that found for Von apple catalase ( Yoruk et al., 2005 ) but half of that obtained for the Chicken erythrocytes (Aydemir and Kuru, 2003). Poor recovery (0.48% ) for this enzyme was also found for the enzyme purified from Chicken erythrocyte (1.68%), unlike to that originated from Wheat germ (11.5%). Although the eluent of gel filtration of the extract was very dilute to find another active fraction, it might indicate of one iso enzyme for this catalase. This was in contrast to finding in Wheat germ (Garcia et al., 2000) with 2 different iso-enzyme and the study of Helmes and Masters (1972) showing the multiplicity of Mammalian liver catalases. The comparison of active fraction of the Brassica oleracea with that of spinacia oleracea leaves (Spinach) prepared under the same conditions may be an indication of higher MW of the enzyme from Cabbage than that of Spinach (results not shown). The active pH of most catalases have shown to be around 5 to 9, with optimal pH around 6.8-7.5 (Aebi, 1984). In this study the optimum pH for Cabbage was found around 6 to 8.
There was found a direct relationship between the enzyme concentration and the catalase activity. The velocity of the reaction was reduced when dilute enzyme solution was used. This effect was omitted by addition BSA to the reaction mixture. This could be due to instability of the enzyme at low concentration (Jacob and Johnson, 1979). Increasing the concentration of substrate was in accordance with the velocity of the reaction. No substrate inhibition was observed at high concentration of hydrogen peroxide a phenomenon of tipic catalase (Brown-Peterson and Salin, 1993). The highest concentration of H2O2 used in this study was 0.01 M (10 times less than catalase inactivation threshold). The concentration that was possible to measure the Vmax and Km.
The Km values calculated to be 25.5 mM H2O2 and Vmax values was found to be 31.12 μ mole min-1 for Cabbage leave. This Km Value was seventeen times less than that was found for the van apple (Yoruk et al., 2005) but very similar to that of Chicken erythrocyte (Aydemir and Kuru, 2003). In conclusion cabbage showed a unstable catalase, the activity which could be lost mainly during column chromatography, for this reason other types of chromatography such as that was applied for purification of catalase from leaves of Zantedschia aethiopica (Trindade and Kanmali., 1988) or that the one which used for Bacterium Vibrio rumoiensis s-1 (Yumoto et al., 2000) showed be examined.
ACKNOWLEDGMENTS
This study was supported by a grant from Research Committee of Kerman University of Medical Sciences. The authors also wish to thank the Department of Chemistry Tehran University for their technical assistance and helpful cooperation.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Aydemir, T. and K. Kuru, 2003. Purification of catalase from Chicken erythrocytes. Turk. J. Chem., 27: 85-97.
Direct Link - Garcia, R., N. Kaid, C. Vignaud and J. Nicolas, 2000. Purification and some properties of catalase from wheat germ (Triticum aestivum L.). J. Agric. Food Chem., 48: 1050-1057.
Direct Link - Holmes, R. and C. Masters, 1972. Specific specific features of the distribution and multiplicity of mammalian liver catalases. Arch. Biochem. Biophys., 48: 217-223.
PubMedDirect Link - Izawa, S., 1996. Importance of catalase in the adaptive response to hydrogen peroxide: Analysis of acatalasaemic Saccharomyces cervisiae. Biochem. J., 320: 61-67.
PubMedDirect Link - Jacob, G.S., S.H. Orme and O. Johnson, 1979. Catalase of neurospora crasa. Induction purification and physical properties. Biochemistry, 18: 2967-2975.
CrossRefDirect Link - Jin, L.H., D.W. Kim, W.S. Eum, C.S. Yoon and S.H. Jang et al., 2003. Human liver catalase: Cloning, expression and characterization of monoclonal antribodies. Mol. Cells, 15: 381-386.
PubMedDirect Link - Leul, M. and W.J. Zhou, 1999. Alleviation of waterlogging damage in winter rape by uniconazole application: Effects on enzyme activity, lipid peroxidation and membrane integrity. J. Plant Growth Regul., 18: 9-14.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Percy, M.E., 1984. Catalase an old enzyme with a new role?. Can. J. Biochem. Cell. Biol., 62: 1006-1014.
PubMedDirect Link - Prasad, K.V.S.K., P.P. Saradhi and P. Sharmila, 1999. Concerted action of antioxidant enzymes and curtailed growth under zinc toxicity in Brassica juncea. Environ. Exp. Bot., 42: 1-10.
CrossRefDirect Link - Sheptovitskey, Y.G.and G.W. Brudvig, 1996. Isolation and characterization of spinach photo system II membrane associated catalase and polyphenol oxidase. Biochemistry, 35: 19255-19263.
CrossRefDirect Link - Singh, P.K. and R.K. Tewari, 2003. Cadmium toxicity induced changes in plant water and oxidative metabolism of Brassica juncea L. plants. J. Environ. Biol., 24: 107-112.
PubMed - Spector, T., 1978. Refinement of the coomassie blue method of protein quantitation. Anal. Biochem., 86: 142-146.
PubMedDirect Link - Trindade, H., A. Karmali, M.S. Pais, 1988. One-step purification and properties of catalase from leaves of Zantedeschia aethiopica. Biochimical, 70: 1759-1764.
PubMedDirect Link - Verma, S. and S.N. Mishra, 2005. Putrescine alleviation of growth in salt stressed Brassica juncea by inducing antioxidative defence system. J. Plant Physiol., 162: 669-677.
CrossRef - Yoruk, I.H., H. Demir, K. Ekici and A. Savran, 2005. Purification and properties of catalase from Van Apple (Golden delicious). Pak. J. Nutr., 4: 8-10.
CrossRefDirect Link - Yumoto, I., D. Ichihashi, H. Iwata, A. Istokovics and I. Nobutoshi et al., 2000. Purification and characterization of a catalase from the facultatively psychrophilic bacterium Vibrio rumoiensis S-1 T exhibiting high catalase activity. J. Bacteriol., 182: 1903-1909.
Direct Link