
J.G. Nyaanga
Not Available
A.W. Kamau
Not Available
J.K. Wanjama
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 4 | Page No.: 713-716







ABSTRACT
Cereal aphids infest plants in distinct patterns both in spatial and temporal terms. The distribution patterns of Rhopalosiphum padi (Linnaeus), Metopolophium dirhodum (Walker) and Schizaphis graminum (Rondani) in time and space on wheat varieties Fahari, Mbega and Duma were studied in field plots and sampled at six growth stages. The experiment was done during the long seasons of 2001 and 2003. Results showed that aphid species differed in their time of colonisation. R. padi appeared at two leaf growth stage followed by S. graminum at the two tiller stage. M. dirhodum came later at the stem elongation stage. Aphid abundance differed among the species, wheat varieties and crop growth stages. The aphid species R. padi, wheat variety Duma and the flowering growth stage had the highest aphid population. The aphid species also differed in their points of colonisation. R. padi was most abundant on stems of wheat at seedlings, lower leaves, flag leaf and the spike as the crop matured. S. graminum mainly colonised the lower leaves. M. dirhodum was mainly on the underside of upper leaves including the flag leaf. The studies showed a crop growth stage and feeding site preferences among cereal aphids.
PDF Abstract XML References Citation
How to cite this article
J.G. Nyaanga, A.W. Kamau and J.K. Wanjama, 2006. Niche and Temporal Differences in Colonization of Wheat Varieties by Three Species of Cereal Aphids. Asian Journal of Plant Sciences, 5: 713-716.
DOI: 10.3923/ajps.2006.713.716
URL: https://scialert.net/abstract/?doi=ajps.2006.713.716
DOI: 10.3923/ajps.2006.713.716
URL: https://scialert.net/abstract/?doi=ajps.2006.713.716
INTRODUCTION
Kenya has the potential to produce enough wheat for local consumption which is estimated at 600,000 MT annually (GoK, 1997). However, the average annual national production of wheat is about 250,000 MT (Cereal Growers Association of Kenya, 2001). Pests and diseases are major factors that have significantly contributed to the low yields of wheat (Arama and Wanjama, 1998). Aphids are the major pests of small grain cereals such as wheat, barley, oats and triticale in Kenya (Wangai et al., 2000). They affect plant growth directly through sap sucking and indirectly as vectors of Barley Yellow Dwarf Virus (BYDV) which can cause yield losses of 8-54% in the field (Wangai and Torres, 1991). According to Muthangya et al. (1991) and Wangai (1994), the most important and commonly found species of aphids on cereal crops in Kenya are English grain aphid (Sitobion avenae F.), Rose grain aphid (Metopolophium dirhodum W), Bird cherry aphid (Rhopalosiphum padi L.) and the Greenbug (Schizaphis graminum R).
Cereal aphids infest plants in distinct patterns, both in spatial and temporal terms (Gianoli, 2000). Adults of each species are more selective than juveniles in determining suitable locations for settlement, feeding and reproduction. Feeding site preferences are considered important, particularly in relation to the type and amount of damage caused by various aphid species (Vickerman, 1979). Research by Leather and Dixon (1981), Wiktelius (1987) and Wiktelius et al. (1990) in discussing patterns of within-plant distribution of R. padi considered factors such as nutritional value of tissues and avoidance of extreme temperature in the field.
The type of host plant and its growth stage markedly affect colonization and the subsequent rate of increase in aphid abundance on crops (Guildemond et al., 1998). Yield reduction in crops caused by virus infection is influenced by the growth stage at which a plant is infested by the vectors. Greatest damage is caused when plants are infested at early growth stages (Watson, 1942; Doodson and Saunders, 1970).
Cereal aphids also have differential performance on different hosts even at the varietal level. Hesler et al. (1999), found that MV4 wheat variety lowered the intrinsic rate of natural increase of R. padi. He also showed that the proportion of alate R. padi was greater on variety MV4 than most other accessions indicating that the former is not a suitable host for the pest. Cultivar and aphid species interact with plant growth stage in increase in abundance of aphids (Guldemond et al., 1998).
The patterns of increase in abundance of different cereal aphids on a wheat crop in Kenya have not been documented. This phenomenon can be better understood by studying the movement of individuals or colonies on the plants. Information on population fluctuations of cereal aphids is essential for determining control strategies. When the pattern of aphid infestation of crops is well documented, specific recommendations on date of planting, time and method of chemical application and other management strategies can be developed. This study sought to investigate wheat varietal differences through the pattern of infestations and increase in abundance of three cereal aphids namely; R. padi, S. graminum and M. dirhodum.
MATERIALS AND METHODS
Three wheat varieties (Duma, Mbega and Fahari) were planted using a factorial Randomised Complete Block Design (RCBD) replicated four times during the long rains of 2001 and 2003 at KARI-Njoro. The experimental plots were 1.5x6 m with an inter-row spacing of 20 cm. The plots and replications were separated by 0.5 m and 1m gap respectively. Di-ammonium phosphate (DAP) 18:46:0 (NPK) fertilizer at a rate of 90 kg ha-1 was hand drilled into the soil and well mixed before hand planting the wheat at a rate of 90 kg ha-1 of seeds. Weeding was done by hand at growth stage 16 (Zadok et al., 1974).
Observations were made on the times and locations of colonisation by the three species of cereal aphids (R. padi, S. graminum and M. dirhodum) on the three varieties of wheat at six growth stages. The wheat growth stages used were: GS 12 (2-leaf stage), GS 22 (2-tiller stage), GS 32 (stem elongation stage) GS 45 (booting stage, GS at 69 (flowering stage) and GS 91 (ripening stage) (Zadok et al., 1974). Data was collected from ten plants selected randomly in each plot.
Data on the mean number of aphids of each species on three wheat varieties at six growth stages was transformed using the square root (1+x) to homogenise and stabilise the variance. General linear model (GLM) was used to help analyse the data (SAS Institute; 1996). Means were compared using the Least Significant difference (LSD) tested at the 5% level. Point of colonisation was analysed using descriptive analysis.
RESULTS
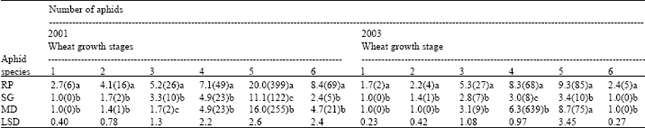
Abundance of R. padi on the three varieties of wheat was significantly greater (p<0.05), compared by those of M. dirhodum and S. graminum in both 2001 and 2003 (Table 1). Abundance of all the three species of cereal aphid was greatest at growth stage 69 (flowering) in both years. There were no significant differences in abundance of M. dirhodum and S. graminum before flowering stage in 2003. There was however some differences in abundance between the two species at 2-tiller and stem elongation in 2001. Abundance of all the three aphid species decreased abruptly at growth stage 91(ripening) during the two years but more distinctively in 2003.
Table 1 also reveals that there were distinct differences in the time of colonisation among the three species of aphid. R. padi appeared earlier, at the 2-leaf stage, followed by S. graminum at the 2-tiller stage and last was M. dirhodum at stem elongation. S. graminum and M. dirhodum disappeared after the crop flowered in 2003. The order and time of colonisation of the three cereal aphids did not differ significantly among the three varieties of wheat both in 2001 or 2003.There were mixed results in the abundance of aphids among the wheat varieties except for Duma, which was consistently infested with more aphids (Table 2) during the two years.
R. padi changed sites of greatest colonization with the age of the plant. R. padi was mainly on the stems of wheat seedling, the aphids moved to the leaves and the leave sheaths during the stem elongation stage. At the latter stages, R. padi was mainly observed on the flag leaves and spikes. M. dirhodum was found on the underside of the middle and flag leaves. M. dirhodum shared a niche with R. padi on the top green leaves including the flag leaf at ripening stage in 2001. Rarely did M. dirhodum move to the spikes. S. graminum colonies were found on the upper side of the lower leaves which turned necrotic within a few days of colonisation.
| Table 1: | Three cereal aphid species numbers per ten plants at six wheat growth stages |
 | |
| Table 2: | Number of aphids per ten plants of three varieties of wheat |
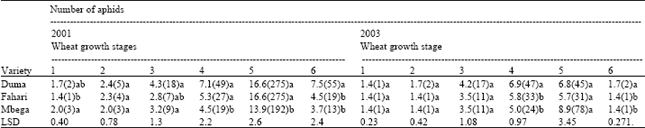 | |
| • Means in the same column followed by the same letter are not significantly different (LSD= 0.05), • Wheat growth stages (1-leaf, 2-tiller, 3-elongation, 4-booting, 5-flowering and 6-ripenining), • Figures in parentheses represent the original data | |
The aphids completely destroyed the lower leaves and progressed upwards to the middle leaves where they settled mainly at the angled points of these leaves. Rarely were the aphids found on the upper and flag leaves. They were completely absent on the spikes in both years.
DISCUSSION
The three species of cereal aphids occupied different feeding sites on the wheat plants. The occurrence of different species of cereal aphids at different growth stages of the crop and at different feeding sites probably to reduces inter specific competition allows aphids to fully exploit the host plant. Results for R. padi are consistent with those obtained by others. Holland and Thomas (1997) found this aphid on crops four weeks after planting. Wiktellius et al. (1990) observed that R. padi arrived first in the season (late spring-early summer) when cereals are in the seedling or early tillering stage. He further noted that abundance peaked during the stem elongation stage and declined during ear emergence. In this study however, abundance of aphids peaked at early flowering and declined abruptly at early ripening stage in 2001 and 2003.
The strategy of early colonisation (seedling) stage makes R. padi a pest among cereal aphids. This is confounded by the ability of R. padi to exploit a host at different stages of growth i.e., from the tender and nutritious seedling stage to plants at the deteriorating ripening stage, irrespective of variety as observed in this study. This aphid was found to aggregate in colonies on the tender stems during the seedling stage, the middle leaves during the elongation stage and the flag leaf and spike in the latter stages. It is a serious pest since it occurs in large numbers and inflicts serious direct damage through removal of sap. R. padi reduced plant biomass dramatically, regardless of the growth stage (Gianolli, 2000).
S. graminum mostly exploits crops at the seedling stage (Wanjama, 1986). It mainly feeds on leaves of the lower half of the plant. Its preferred niche was on the angled points of the leaves probably to avoid extremely hot and dry field conditions (Wiktelius, 1987) or direct impact of rain splash. Direct and side effects of feeding by S. graminum damages crops through sap removal and injection of toxic saliva, a few aphids are likely to cause severe crop damage particularly at early growth stages. Riedell and Blackman (1999) found that infestations by S. graminum also reduced dry weight, leaf area and chlorophyll concentration of upper and younger leaves. Results from this study showed that offsprings of S. graminum had almost killed the plant by six days after infestation; plants were drooping and wilting. Therefore, outbreaks of S. graminum are likely to cause devastating crop losses.
M. dirhodum arrived later at stem elongation. Its preference of the cool underside of the dense canopy created by the middle leaves could be avoidance of extreme temperature in the field (Wiktelius, 1987). The abundance of M. dirhodum declined abruptly as the foliage thinned out because of crop senescence at the ripening stage. In 2001, M. dirhodum shared a niche with R. padi on the few top green leaves including the flag leaf at ripening stage but none of these aphids settled on the spike. In 2003 however, this species of aphid was absent at ripening stage. A similar pattern for M. dirhodum was observed by Watt (1979) and Howard and Dixon (1992). The latter observed that very many M. dirhodum alates (winged forms) develop on mature plants and fly away even when the number of aphids on each plant is very low. The production of winged forms when wheat is mature results from reduced plant nutrition (Dixon and Glen, 1971) caused by aging and hardening of the awns and kernels. In 2003 however, rainfall was greater than in 2001. The rainfall that continued until the ripening stage during this year could have also contributed to the generally slow increase in abundance of all the aphid species and the absence of S. graminum and M. dirhodum at this stage. Aphid species are greatly reduced by the impact of rain splash (Wiktelius, 1987).
The mixed results in the abundance of aphids in the three wheat varieties (Table 2) may suggest little or absence of resistance for these varieties. Wanjama (1986) however reported some resistance in Kenya Fahari to the greenbug (S. graminum) because it was resistant for all mechanisms of resistance tested. Further work is required to establish whether this holds true for other species of cereal aphids.
ACKNOWLEDGMENTS
We greatly acknowledge the financial and material support given towards this work from the following institutions; Kenya Agricultural Institute (KARI) Njoro, Egerton University and Winrock International.
REFERENCES
- Dixon, J.K. and D.M. Glen, 1971. Morph determination in the bird cherry oat aphid, R. padi L. Annal. Applied Biol., 68: 11-21.
Direct Link - Gianoli, E., 2000. Competition in cereal aphids (Homoptera: Aphididae) on wheat plants. Environ. Entomol., 29: 213-219.
Direct Link - Guildemond, J.A., W.J. Brink and E. Belder, 1998. Methods of assessing population increase in aphids and the effect of growth stage of the host on population growth rate. Entomol. Exp., 86: 163-173.
Direct Link - Hesler, L.S., W.E. Riedell, S.D. Haley and R.D. Collins, 1999. Resistance to R. padi (Homptera: Aphididae) in wheat germplasim accessions. J. Econ. Ent., 92: 1234-1238.
Direct Link - Holland, J.M. and S.R. Thomas, 1997. Quantifying the impact of polyphagous predators in controlling cereal aphids and in preventing wheat yield and quality reduction. Ann. Applied Sci., 131: 375-397.
Direct Link - Howard, M.T. and A.F.F. Dixon, 1992. The effect of plant phenology on the induction of alatae and the development of populations of Metapolophium dirhodum (Walker), the rose-grain aphid, on winter wheat. Ann. Applied Biol., 120: 203-213.
Direct Link - Leather, S.R. and A.F.G. Dixon, 1981. The effect of cereal growth stage and feeding site on the reproductive activity of the bird cherry aphid, R. padi. Ann. Applied Biol., 97: 135-141.
Direct Link - Muthangya, P., S.M. Migui, M. Macharia and J.K. Wanjama, 1991. Survey of cereal aphid predominance and barley yellow dwarf disease incidence in wheat and barley growing areas of kenya. Turner, D.G. and E.M. Mwangi (Eds.). Proceedings of the 7th Regional Wheat Workshop for Eastern, Central and Southern Africa. CIMMYT. Nakuru, Kenya.
- Riedell, W.E. and T.M. Blackman, 1999. Leaf reflectance and spectra of cereal aphid damaged wheat. Crop Sci., 39: 1835-1840.
Direct Link - Wangai, A.W., R.T. Plumb and H.F. Van-Emden, 2000. Effects of sowing date and insecticides on cereal aphid populations and bydvon barley in Kenya. J. Phytopathol., 148: 33-37.
Direct Link - Watt, A.D., 1979. The effect of cereal growth stages on the reproductive activity of S. avenae and M. dirhodum. Ann. Applied Biol., 92: 147-157.
Direct Link - Watson, M.A., 1942. Sugar beet yellow virus: A preliminary account of experiments and observations on its effects on the field. Ann. Applied. Biol., 29: 358-365.
Direct Link - Wiktelius, S., 1987. Distribution of Rhopalosiphum padi on spring barley plants. Ann. Applied Biol., 110: 1-7.
Google - Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link