
Tolga Goksan
Not Available
Ilknur Ak
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 3 | Page No.: 455-460







ABSTRACT
The growth of the green alga Haematococcus pluvialis in vegetative stage is one of the most important parts in the whole cultivation process. The aim of the study was to show the growth characteristics of the alga under 100 and 170 μmol photon m-2 s-1 light intensities in the longer Light Path (LP) lengths (17.5, 23.0 and 27.5 cm) than the ones usually used. Maximum cell numbers increased from 15.7, 11.5 and 10.1 x 104 cells mL-1 in 100 μmol light to 17.9, 12.9 and 11.1 x 104 cells mL-1 under 170 μmol light for 17.5, 23.0 and 27.5 cm LP lengths, respectively. The cell concentration per liter volume increased as in the pattern of cell count. The Chl/Car ratio in the whole groups, a very useful indicator to evaluate the stress on the cells, increased by the shift of light from 100 to 170 μmol. The close correlation between cell number and absorbance value at 680 nm (p<0.04) showed the biomass concentration to depend on the cell count rather than DW and cell diameter. In addition, cell diameter did not change during the experiments. As a result of the findings, the use of longer LP lengths may be an alternative method in the vegetative cultivation of the light sensitive alga Haematococcus pluvialis by the higher light intensities than 200 μmol photon m-2 s-1 using mutual shading effect.
PDF Abstract XML References Citation
How to cite this article
Tolga Goksan and Ilknur Ak, 2006. Vegetative Growth of the Green Alga Haematococcus pluvialis Cultivated in Different Light-Path Lengths. Asian Journal of Plant Sciences, 5: 455-460.
DOI: 10.3923/ajps.2006.455.460
URL: https://scialert.net/abstract/?doi=ajps.2006.455.460
DOI: 10.3923/ajps.2006.455.460
URL: https://scialert.net/abstract/?doi=ajps.2006.455.460
INTRODUCTION
Haematococcus pluvialis (Chlorophyceae) is one of the most important species in microalgal biotechnology due to the capability of accumulating a commercially valuable pigment astaxanthin (3,3'-dihydroxy-β, β-carotene-4-4′-dione) in massive amounts under several stress factors such as high salinity and temperature, nutrient deficiency and especially high irradiance (Boussiba, 2000; Torzillo, 2003). Although some other organisms, such as a yeast species Phaffia rhodozyma, can synthesize astaxanthin (Bon et al., 1997), the amount accumulated is very low to compete with Haematococcus. In this respect, Haematococcus is a unique organism yielding the highest astaxanthin concentration in all over the biological sources in the world, e.g., up to 5% of the dry weight (Harker et al., 1996; Torzillo et al., 2003).
Although the astaxanthin, mostly the synthetic one, is basically used to improve the coloration of fins, skin and flesh of the cultured salmon and trout fish, it has further applications such as ornamental fish breeding, shrimp and poultry farming to improve the color of the egg yolk sack (Yamada et al., 1990; Christiansen et al., 1995; Inborr, 1998, Ako and Tamaru, 1999). Besides, many researchers have recently focused on the clinical application of astaxanthin (Guerin et al., 2003). In this respect, astaxanthin from Haematococcus has attracted great interest due to the facts that it is used in a variety of fields, accumulated as much as 5% of dry weight, produced biologically and consumers become more conscious on natural products rather than the synthetic ones.
Astaxanthin accumulation in Haematococcus is generally achieved through a two-stage culture, e.g., green (vegetative) and red stage. This procedure to produce the cysts rich by astaxanthin suffers from certain drawbacks, especially in vegetative stage due to the difficulties to reach higher biomass concentrations, low specific growth rate and contamination problems. Therefore, the vegetative cultivation of Haematococcus is the most problematic part in astaxanthin production. Most of the light optimisation experiments on Haematococcus reported that growth was saturated below 100 μmol photon m-2 s-1 (Fan et al., 1994; Harker et al., 1995). However, many other factors are responsible for the effective exploitation of light by the cells, such as cell concentration, mixing rate, light-path (LP) length and geometry of the cultivation vessel (Richmond, 2003).
Almost, all of the vegetative Haematococcus cultures under laboratory conditions have been cultivated in the common culture flasks with short LP lengths, ranging from 5 to 15 cm. The cultivation of the cells in the vessels having longer LP length was not achieved to date. So, this work aimed to examine the changes in growth parameters in longer LP lengths (17.5, 23.0 and 27.5 cm) under 100 and 170 μmol photon m-2 s-1 light intensities.
MATERIALS AND METHODS
The microalga Haematococcus pluvialis (strain 5/99) was obtained from the culture collection of the CNR, Istituto per lo Studio degli Ecosistemi of Florence (Italy). The first inocula were provided by the green cells acclimated to 50 μmol photon m-2 s-1. Temperature was kept constant at 25±0.5°C and cultures were aerated by bubbling air under continuous illumination. Cultures were illuminated by cool-white two fluorescent lambs at two different light intensities of 100 and 170 μmol photon m-2 s-1. Photosyntheticaly Active Radiation (PAR) was measured with a LI-250A light meter (Li-Cor USA). The cells were grown in 5.0, 10.0 and 19.0 L cylindrical transparent barrels having 17.5, 23.0 and 27.5 cm Ligh Path (LP) lengths, respectively. The heights of the barrels were kept at 13 cm to equalise the illumination to be exposed to. So, the working volumes of the barrels were found to be 3.0, 4.5 and 6.0 L for 17.5, 23.0 and 27.5 cm LP lengths, respectively. The cells were cultivated in BG-11 growth medium (Rippka et al., 1979) in batch cultures. The initial dry weights in both 100 and 170 μmol photon m-2 s-1 light intensities were 50 mg L-1, which corresponded to a cell count of 1.8x104 cells mL-1. The low concentration was set to minimise the self-shading of the cells. Growth was carried out 13 and 15 days for 100 and 170 μmol photon m-2 s-1, respectively. Each experiment was repeated three times.
Absorbance values at 680 nm wavelength (Abs680) and pigment extracts for chlorophyll and carotenoids analysis were read in a Jasco UV/Visible Spectrophotometer. For Dry Weight (DW) measurements, 25 mL sample was filtered through a pre-dried and pre-weighed GF/C Whatman filter papers in duplicate. Dws were calculated in mg L-1 after the filter papers were dried in an oven at 105°C for two hours. Total chlorophyll (chlorophyll a and b) and carotenoids were calculated according to Lichtenthaler (1987). Cell count was found by triplicate samples in a Neubauer haemocytometer. Specific growth rate (μ) and doubling time (d.t.) were calculated according to the equations below:
X2 and X1 in the equation represent the biomass concentrations at the times t2 and t1, respectively.
RESULTS
The changes in the maximum Dry Weight (DW) values were compared in Fig. 1. DW values increased in each LP group by increasing light, while declining with increasing LP lengths. The effect of the increase in irradiance was higher at lower LP lengths. The increase in light intensity had no effect on the longest LP group (27.5 cm). The highest DW values in 17.5 cm LP showed a reduction 33 and 40% in 27.5 cm LP for 100 and 170 μmol lights, respectively.
The cell number indicated the same pattern as the DW (Fig. 2). The counts increased in the same LP lengths by the shift of light intensity from 100 μmol to 170 μmol. The increase of LP length resulted in a decrease of cell density per culture volume. In 17.5 cm LP group, the cells carried on the increase until the 9th day in 100 μmol (15.7x104 cells mL-1). However, the highest cell number was reached on the 10th day to be 17.9x104 cells mL-1 by 170 μmol light intensity. Maximum cell numbers increased from 15.7, 11.5 and 10.1x104 cells mL-1 in 100 μmol light to 17.9, 12.9 and 11.1 x 104 cells mL-1 under 170 μmol light for 17.5, 23.0 and 27.5 cm LP lengths, respectively. All of the groups attained to maximum values one day later by the shift of the light to 170 μmol. In addition, the cell density of all the experimental groups demonstrated a close correlation with the absorbance value at 680 nm (p<0.04). This correlation revealed that the increase in biomass concentration was dependent to the cell count, not to the parameters such as DW value or diameter of the cells, increase of which sign to the stress. In addition, the cell diameter did not change much in the groups (23±0.8 μm).
Although the chlorophyll and carotenoids levels per liter volume reduced with the increase in LP length (Fig. 3 and 4) as in the dry weight, they interestingly increased in cell basis (Fig. 5) and DW basis (not shown) at the end of log phase for each group.
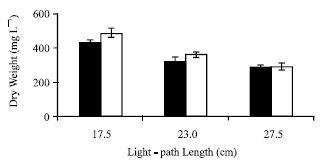 | |
| Fig. 1: | Maximum DW values under 100 (filled) and 170 (empty) μmol photon m-2 s-1 light intensities at 17.5, 23.0 and 27.5 cm light-path lengths (vertical bars represent±SD (n = 3) |
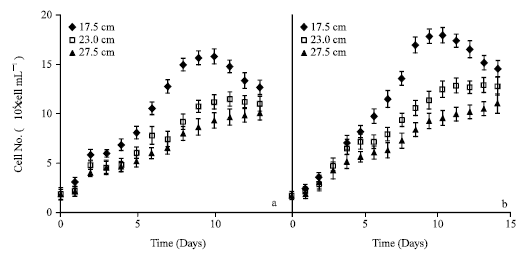 | |
| Fig. 2: | Increase in cell density under 100 (a) and 170 (b) μmol photon m-2 s-1 light intensities at 17.5, 23.0 and 27.5 cm light-path lengths (Each value represents the mean of 3 experiments, less than 15%) |
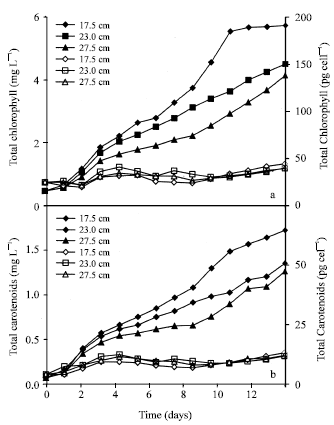 | |
| Fig. 3: | Changes in chlorophyll (a) and carotenoids (b) amounts per liter volume (filled) and per cell (empty) under 100 μmol photon m-2 s-1 light intensity at 17.5, 23.0 and 27.5 cm light-path lengths (Each value represents the mean of 3 experiments, less than 8%) |
Similarly, the shift up of light to 170 μmol caused the same effect on the cells in each LP group (Fig. 2). The levels of chlorophyll and carotenoids dramatically increased, especially by the end of log phase.
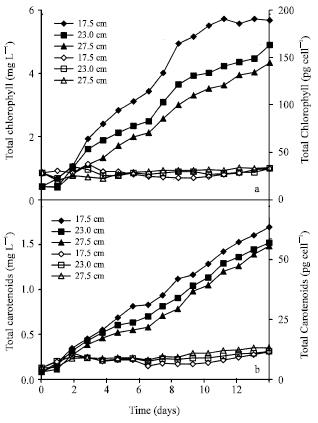 | |
| Fig. 4: | Changes in chlorophyll (a) and carotenoids (b) amounts per liter volume (filled) and per cell (empty) under 170 μmol photon m-2 s-1 light intensity at 17.5, 23.0 and 27.5 cm light-path lengths (Each value represents the mean of 3 experiments, less than 10%) |
Chlorophyll and carotenoids started to increase in the first 4 days and then to decrease in the following 4 days at 100 μmol light.
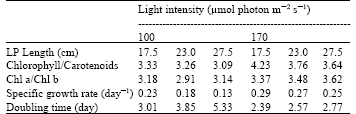
| Table 1: | The mean chlorophyll/carotenoids and Chl a/Chl b ratios, Specific growth rates and Doubling times at 17.5, 23.0 and 27.5 cm LP lengths with 100 and 170 μmol photon m-2 s-1 light intensity |
 | |
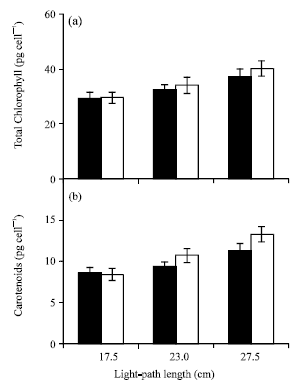 | |
| Fig. 5: | Comparison of the chlorophyll and carotenoids per cell at the end of log phase under 100 (filled) and 170 (empty) μmol photon m-2 s-1 light intensities at 17.5, 23.0 and 27.5 cm light-path lengths (vertical bars represent±SD (n = 3) |
The levels started to increase again after the log phase until the end of the experiment. The same pattern was also observed with the 170 μmol light.
The ratio of chlorophyll to carotenoids was enhanced in all the LP lengths by the increase in light intensity. In contrary, the ratio in both light intensities diminished by increasing LPs. The ratios of chlorophyll to carotenoids, Chl a to Chl b, specific growth rates (μ) and doubling times were given in Table 1.
DISCUSSION
Most of the experiments on light optimisation in vegetative stage of Haematococcus took into account just levelling of light intensity (Torzillo et al., 2005; Göksan and Gökpınar, 2005; Fan et al., 1994). However, there are various factors affecting light regime of the cells rather than just the manipulation of light intensity, such as cell concentration, mixing rate and Light Path (LP) length (Richmond, 2003). Each species shows a specific light requirement, therefore, a specific LP length. For instant, 10 cm is optimal for outdoor cultivation of Nannochloropsis sp. (Zou and Richmond, 1999), while 20 cm is optimal for Chaetoceros and Isochrysis sp. (Zhang and Richmond, 2003), which are sensitive to high light intensities. The same concept is also valid for Haematococcus in the vegetative stage, being so susceptible to shift up in light intensity. The cells are immediately triggered to accumulate the secondary carotene astaxanthin by the increase in light intensity. To see the effect of LP length, the cultures were exposed to the light intensities of 100 and 170 μmol photon m-2 s-1 in three LP groups, e.g., 17.5, 23.0 and 27.5 cm.
The shift of light to 170 μmol had a positive effect on DW (in liter volume) over the groups except for the one having the longest LP 27.5 cm. No change was observed in this group. The chlorophyll and carotenoids gradually increased in cell and DW basis by increasing light intensity and LP length. However, it was reported that chlorophyll molecules in DW basis declined by increasing light while carotenoids went up in the experiment 5 different light intensities were applied (Torzillo et al., 2005). The demolition of chlorophyll molecules in the experiment by the increase of light was observed even in the groups illuminated very low, i.e., from 50 to 100 μmol photon m-2 s-1. The contradiction between the results aroused from the difference in cell density and LP length. The researchers carried out the experiment at a constant chlorophyll concentration of 4 μg mL-1 by a semi-continuous harvesting regime in 5 cm Roux bottles, while our cultures were batch. It is clear that the cells immediately start to accumulate carotenoids and chlorophyll molecules gradually decrease by increasing light intensity as shown in Torzillo et al. (2005), unless mutual shading of the cells exist. As for in the cultures that mutual shading takes place, both of the chlorophyll molecules and carotenoids carry on increasing.
This concept was further indicated by the ratio of chlorophyll to carotenoids in Table 1. It is a generally accepted phenomenon that Chl/Car ratio is declined by the increase of light intensity, when the cells become stressed (Cifuentes et al., 2003; Torzillo et al., 2003). Present results showed that the Chl/Car ratio in 17.5 cm LP group decreased gradually from 3.33 to 3.26 in 27.5 cm LP group at 100 μmol illumination. The same effect was seen by the shift of light to 170 μmol, decreasing from 4.23 to 3.64. These surprising results may easily lead someone to an opinion that the cells were under stress, so the ratio started to decrease. In our opinion, the increase of LP length in the cultures must have caused a reduction effect in light intensity. However, the shift of light to 170 μmol caused the ratios of the same LP lengths to increase from 3.33, 3.26 and 3.09 to 4.23, 3.76 and 3.64 for 17.5, 23.0 and 27.5 cm LP lengths, respectively. This phenomenon was interpreted as in the following; both of the chlorophyll and carotenoids per cell increased, but the carotenoids were accumulated faster even in the low light like 100 μmol photon m-2 s-1 that is optimal for vegetative growth of Haematococcus. When the carotenoids started to be accumulated, they kept on accumulating. However, the situation in the chlorophyll molecules was differed from carotenoids in that the cells requiring more light up to a level becoming growth saturated. The increase of the Chl/Car ratio was dependent on the increase of light intensity and that was showed by the shift up of light to 170 μmol. In our opinion, the use of even much higher light intensities above 200 μmol would enhance the ratio positively.
Zhekisheva et al. (2002) observed that dry weight, chlorophyll and carotenoids in cell basis showed three differentiated stage i.e., a rise stage during the first 2 days, a decrease stage of following 2 days and the last stage when the cells increased the components. The cells reached at the end of first stage to the weights of about 4.5 and 8.0 ng cell-1 in 75 and 350 μmol light intensity, respectively. The same differentiation was observed in our study as well, the intervals between the stages being 4 instead of 2 days. The maximum cell weights were found to be 2.1 and 2.6 ng for 100 and 170 μmol light. However, the cell DW in the study of Zhekisheva et al. (2002) were decreased below to 2 ng after the cells adapted to the culture conditions, while they were above 2 ng per cell-1 in our study. The strength of the differentiation was associated with the short length of LP, because such a big fluctuation between the stages in our study was not observed.
The specific growth rate (μ) on DW basis in BG-11 medium was much lower than that found by Fan et al. (1994). While μ of the researchers in 4 cm LP length was 0.035 h-1 with 150 μmol light, it was found in our experiment in the shortest LP group (17.5 cm) to be 0.29 d-1 (0.012 h-1) with 170 μmol light. The μ was about 1/3 of the other study. However, it should be noted that μ increased from 0.0096 to 0.012 h-1 by the shift of light from 100 to 170 μmol. The cell counts were found to be 27.3 x 104 cells mL-1 in the experiment of Domínguez Bocanegra et al. (2004) carried out in 1 L volume erlenmeyer containing BG-11 medium with a light of 177 μmol, while 17.9x104 cells mL-1 in our study with the shortest LP length. The cell numbers attained were very close to each other, just being 1.5 times higher. The differences in μ and cell counts were just because of the LP lengths. In addition, the cell diameter, which is another important indicator parameter of the cells to understand whether the stress on the cells s tarted, did not change much in the groups (mean value of 23±0.8 μm).
As can be seen, the use of longer LP lengths under low light intensities, usually used in vegetative cultivation in Haematococcus, did not create a significant difference in terms of cell count when compared to the studies carried on the similar conditions except the study made by Fan et al. (1994) and Zhekisheva et al. (2002) who used a very short LP length of 4 cm. However, the difference derived from LP length can be compensated by the exposition of the cells to the higher illuminations avoiding the cells to become stressed. In this respect, to increase the LP length using very high illumination, i.e., above 200 μmol photon m-2 s-1, which is considered to be a threshold light intensity triggering the astaxanthin accumulation, may be a good solution for the large volume commercial production of Haematococcus cultures in vegetative stage.
ACKNOWLEDGMENTS
This work was supported by the Scientific Research Project (BAP 2004/30) of Çanakkale Onsekiz Mart University.
REFERENCES
- Bon, J.A., T.D. Leathers and R.K. Jayaswal, 1997. Isolation of astaxanthin overproducing mutants of Phaffia rhodozyma. Biotechnol. Lett., 19: 109-112.
Direct Link - Boussiba, S., 2000. Carotenogenesis in the green alga Heamatococcus pluvialis Cellular physiology and stress response. Physiol. Plantorum, 108: 111-117.
Direct Link - Christiansen, R., O. Lie and O.J. Torrissen, 1995. Growth and survival of Atlantic salmon, Salmo salar L., fed different dietary levels of astaxanthin. Juveniles Aquacult. Nutr., 1: 189-198.
Direct Link - Cifuentes, A.S., M.A. Gonzales, S. Vargas, M. Hoeneisen and N. Gonzales, 2003. Optimization of biomass, total carotenoids and astaxanthin production in Haematococcus pluvialis flotow strain Steptoe (Nevada, USA) under laboratory conditions. Biol. Res., 36: 343-357.
Direct Link - Dominguez-Bocanegra, A.R., I.G. Legarreta, F.M. Jeronimo and A.T. Campocosio, 2004. Influence of environmental and nutritional factors in the production of astaxanthin from Haematococcus pluvialis. Bioresour. Technol., 92: 209-214.
Direct Link - Fan, L., A. Vonshak and S. Boussiba, 1994. Effect of temperature and irradiance on growth of Haematococcus pluvialis. J. Phycol., 30: 829-833.
Direct Link - Guerin, M., M.E. Huntley and M. Olaizola, 2003. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol., 21: 210-216.
Direct Link - Harker, M., A.J. Tsavalos and A.J. Young, 1995. Use of response surface methodology to optimise carotenogenesis in the microalga Haematococcus pluvialis. J. Applied Phycol., 7: 399-406.
Direct Link - Harker, M., A.J. Tsavalos and A.J. Young, 1996. Factors responsible for astaxanthin formation in the chlorophyte Hamatococcus pluvialis. Bioresor. Technol., 55: 207-214.
Direct Link - Lichtenthaler, H.K., 1987. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol., 148: 350-382.
CrossRefDirect Link - Richmond, A., C.W. Zhang and Y. Zarmy, 2003. Efficient use of strong light for high photosynthetic productivity: Interrelationship between the optical path, the optimal population density and cell-growth inhibition. Biomol. Eng., 20: 229-236.
Direct Link - Rippka, R., J. Deruelles, J.B. Waterbury, M. Herdman and R.Y. Stanier, 1979. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology, 111: 1-61.
CrossRefDirect Link - Torzillo, G., T. Goksan, C. Faraloni, J. Kopecky and J. Masojidek, 2003. Interplay between photochemical activities and pigment composition in an outdoor culture of Haematococcus pluvialis during the shift from the green to red stage. J. Applied Phycol., 15: 127-136.
Direct Link - Torzillo, G., T. Goksan, O. Isik and S. Gokpinar, 2005. Photon irradiance required to support optimal growth and interrelations between irradiance and pigment composition in the green alga Haematococcus pluvialis. Eur. J. Phycol., 40: 233-240.
Direct Link - Yamada, S., Y. Tanaka, M. Sameshima and Y. Ito, 1990. Pigmentation of prawn (Penaeus japonicus) with carotenoids. Effect of dietary astaxanthin, beta-carotene and canthaxanthin on pigmentation. Aquaculture, 87: 323-330.
Direct Link - Zhang, C.W. and A. Richmond, 2003. Sustainable, high-yielding outdoor mass cultures of Chaetoceros muelleri var. subsalsum and Isochrysis galbana in vertical plate reactors. Mar. Biotechnol., 5: 302-310.
Direct Link - Zhekisheva, M., S. Boussiba, I. Khozin-Goldberg, A. Zarka and Z. Cohen, 2002. Accumulation of oleic acid in Haematococcus pluvialis (chlorophyceae) under nitrogen starvation or high light is correlated with that of astaxanthin esters. J. Phycol., 38: 325-331.
CrossRefDirect Link - Zou, N. and A. Richmond, 1999. Effect of light-path length in outdoor flat plate reactors on output rate of cell mass and of EPA in Nannochloropsis sp. J. Biotechnol., 70: 351-356.
Direct Link